Ontogenic Development of Digestive Enzymes in Mealworm Larvae (Tenebrio molitor) and Their Suitable Harvesting Time for Use as Fish Feed

 ,
,
Abstract
1. Introduction
2. Materials and Methods
3. Results
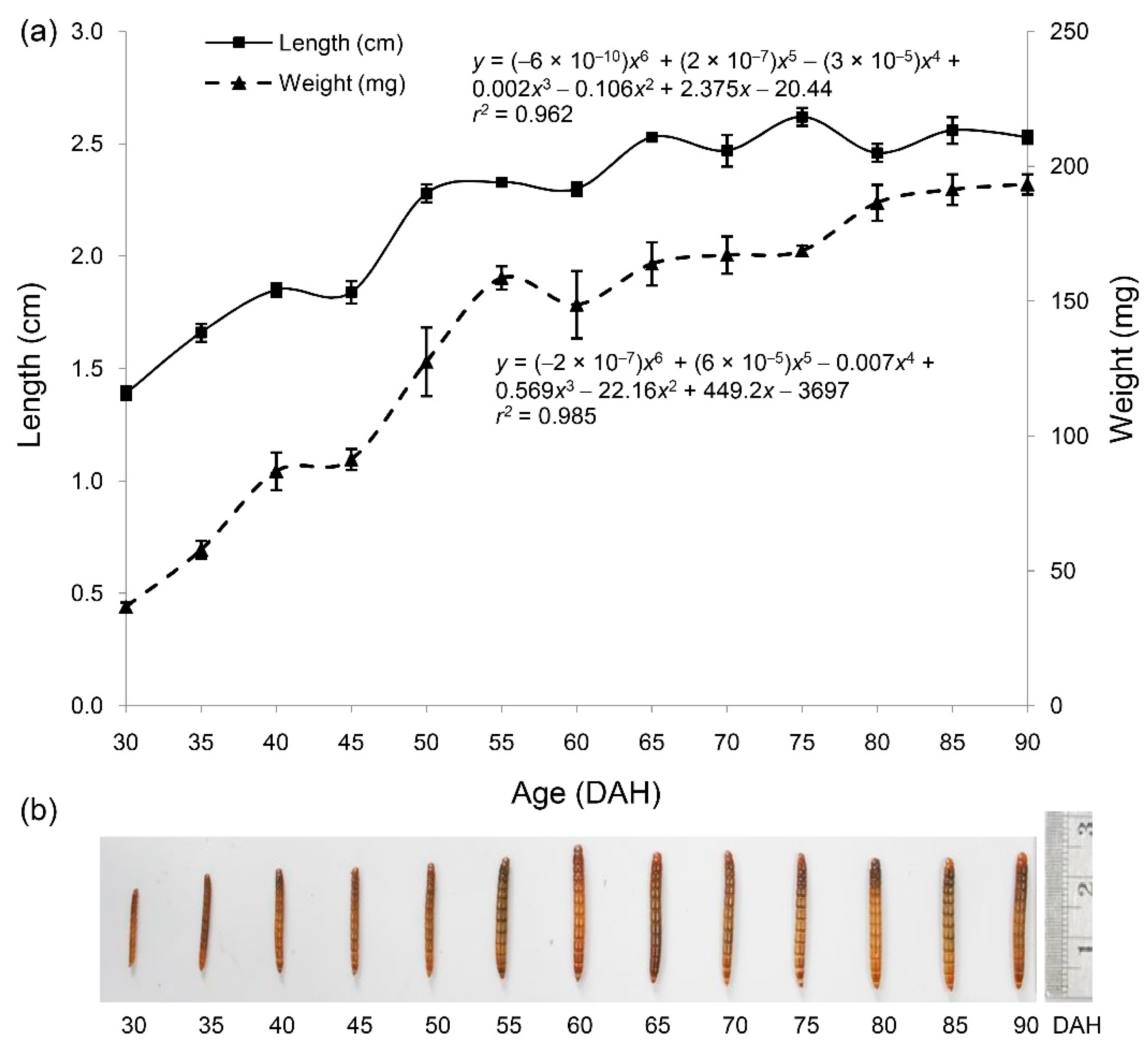
3.1. Changes in Weight and Length across Studied Period
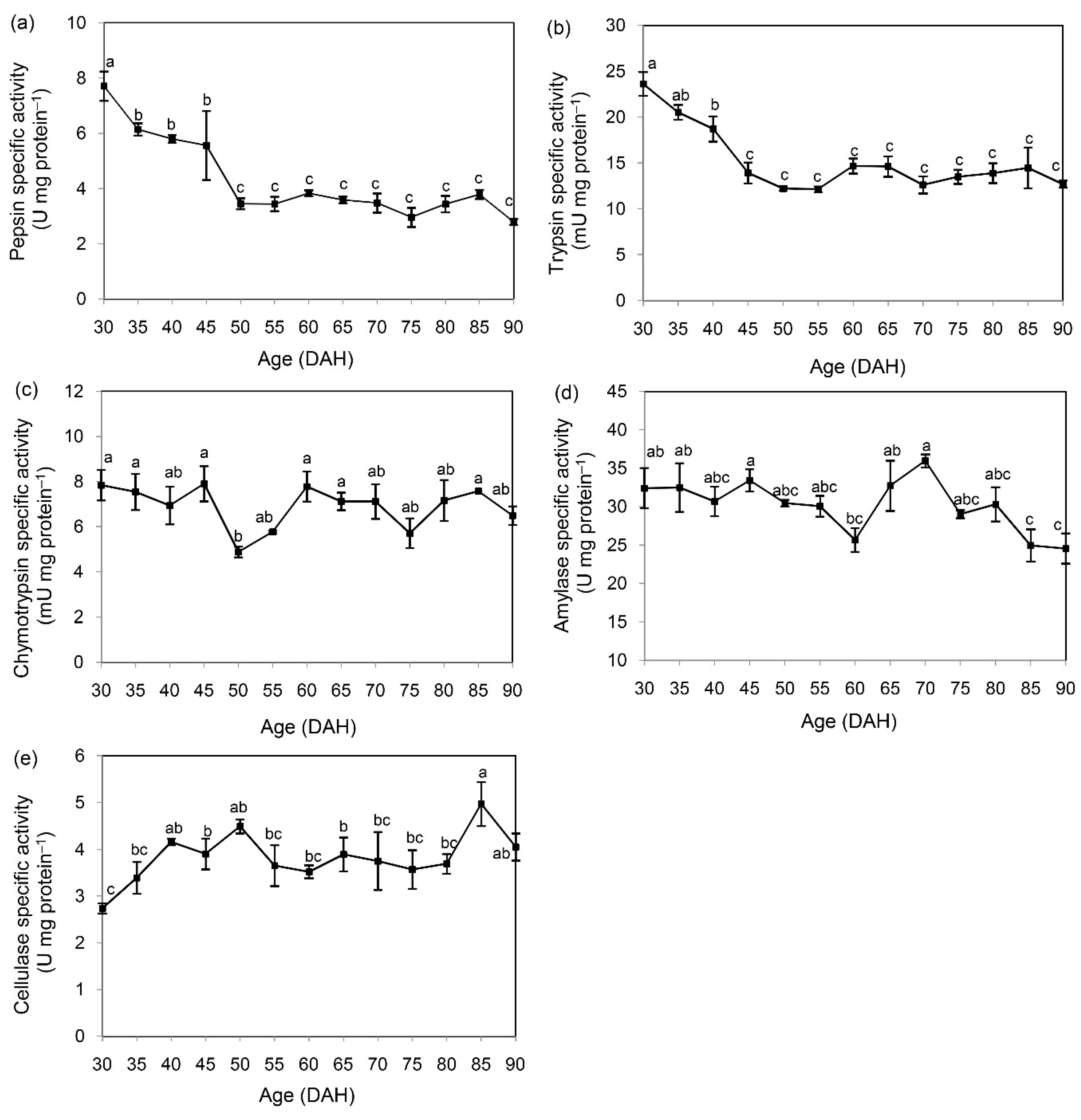
3.2. Ontogenic Development of Digestive Enzymes
3.3. Chemical Composition of Mealworm Larvae
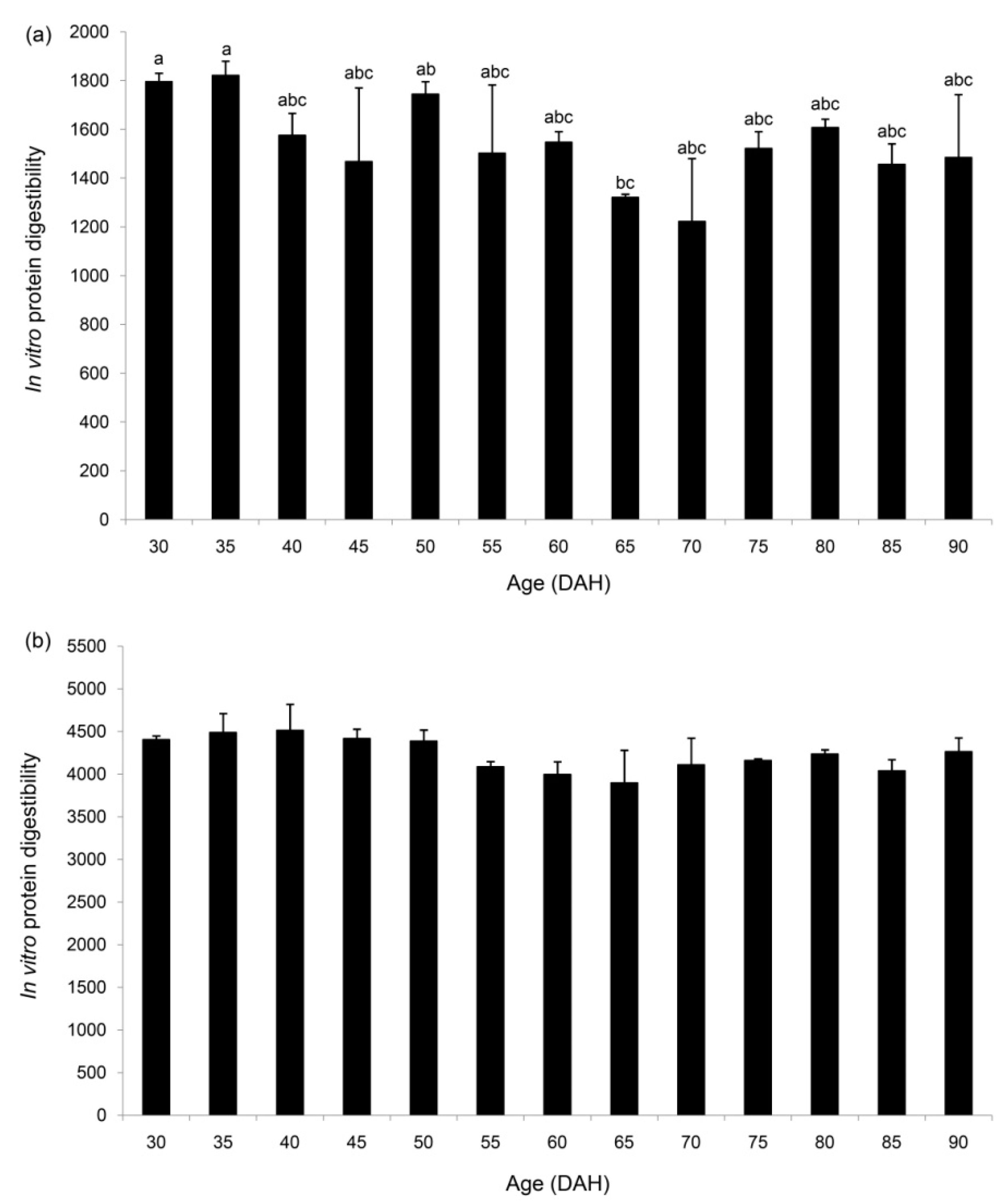
3.4. In Vitro Protein Digestibility Using Digestive Enzymes from Two Fish Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ghosh, S.; Lee, S.-M.; Jung, C.; Meyer-Rochow, V.B. Nutritional composition of five commercial edible insects in South Korea. J. Asia-Pac. Entomol. 2017, 20, 686–694. [Google Scholar] [CrossRef]
- Aguilar-Miranda, E.D.; López, M.G.; Escamilla-Santana, C.; Barba de la Rosa, A.P. Characteristics of maize flour tortilla supplemented with ground Tenebrio molitor larvae. J. Agric. Food Chem. 2002, 50, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Makker, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Ng, W.K.; Liew, F.L.; Ang, L.P.; Wong, K.W. Potential of mealworm (Tenebrio molitor) as an alternative protein source in practical diets for African catfish, Clarias gariepinus. Aquac. Res. 2001, 32, 273–280. [Google Scholar] [CrossRef]
- Lock, E.R.; Arsiwalla, T.; Waagbø, R. Insect larvae meal as an alternative source of nutrients in the diet of Atlantic salmon (Salmo salar) postsmolt. Aquac. Nutr. 2016, 22, 1202–1213. [Google Scholar] [CrossRef]
- Iaconisi, V.; Marono, S.; Parisi, G.; Gasco, L.; Genovese, L.; Maricchiolo, G.; Bovera, F.; Piccolo, G. Dietary inclusion of Tenebrio molitor larvae meal: Effects on growth performance and final quality treats of blackspot sea bream (Pagellus bogaraveo). Aquaculture 2017, 476, 49–58. [Google Scholar] [CrossRef]
- Piccolo, G.; Iaconisi, V.; Marono, S.; Gasco, L.; Loponte, R.; Nizza, S.; Bovera, F.; Parisi, G. Effect of Tenebrio molitor larvae meal on growth performance, in vivo nutrients digestibility, somatic and marketable indexes of gilthead sea bream (Sparus aurata). Anim. Feed Sci. Technol. 2017, 226, 12–20. [Google Scholar] [CrossRef]
- Sánchez-Muros, M.J.; DeHaro, C.; Sanz, A.; Trenzado, C.E.; Villareces, S.; Barroso, F.G. Nutritional evaluation of Tenebrio molitor meal as fishmeal substitute for tilapia (Oreochromis niloticus) diet. Aquac. Nutr. 2016, 22, 943–955. [Google Scholar] [CrossRef]
- Ido, A.; Hashizume, A.; Ohta, T.; Takahashi, T. Replacement of fish meal by defatted yellow mealworm (Tenebrio molitor) larvae in diet improves growth performance and disease resistance in red seabream (Pagrus major). Animals 2019, 9, 100. [Google Scholar] [CrossRef]
- Finke, M.D. Complete nutrient composition of commercially raised invertebrates used as food for insectivores. Zoo Biol. 2002, 21, 269–285. [Google Scholar] [CrossRef]
- Lovett, D.L.; Felder, D.L. Ontogenetic changes in enzyme distribution and midgut function in developmental stages of Penaeus setiferus (Crustacea, Decapoda, Penaeidae). Biol. Bull. 1990, 178, 160–174. [Google Scholar] [CrossRef]
- Elpidina, E.N.; Tsybina, T.A.; Dunaevsky, Y.E.; Belozersky, M.A.; Zhuzhikov, D.P.; Oppert, B. A chymotrypsin-like proteinase from the midgut of Tenebrio molitor larvae. Biochimie 2005, 87, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Applebaum, S.W.; Jankovic, M.; Birk, Y. Studies on the midgut amylase activity of Tenebrio molitor L. larvae. J. Insect Physiol. 1961, 7, 100–108. [Google Scholar] [CrossRef]
- Rivers, D.B.; Acca, G.; Fink, M.; Brogan, R.; Schoeffield, A. Spatial characterization of proteolytic enzyme activity in the foregut region of the adult necrophagous fry, Protophormai terraenovae. J. Insect Physiol. 2014, 67, 45–55. [Google Scholar] [CrossRef]
- Chandang, P.; Thongprajukaew, K.; Chotimanothum, B.; Kovitvadhi, A.; Kovitvadhi, U.; Pakkong, P. The effects on in vitro digestibility from different developmental stages of silkworm larvae, Bombyx mori (Lepidoptera: Bombycidae) and position of mulberry leaves, Morus alba (Rosales: Moraceae). J. Asia-Pac. Entomol. 2017, 20, 1134–1139. [Google Scholar] [CrossRef]
- Kovitvadhi, A.; Chundang, P.; Thongprajukaew, K.; Tirawattanawanich, C.; Srikachar, S.; Chotimanothum, B. Potential of insect meals as protein sources for meat-type ducks based on in vitro digestibility. Animals 2019, 9, 155. [Google Scholar] [CrossRef] [PubMed]
- Marono, S.; Piccolo, G.; Loponte, R.; Di Meo, C.; Attia, Y.A.; Nizza, A.; Bovera, F. In vitro crude protein digestibility of Tenebrio molitor and Hermetia illucens insect meals and its correlation with chemical composition traits. Ital. J. Anim. Sci. 2015, 14, 338–343. [Google Scholar] [CrossRef]
- Carpenter, J.W.; Mashima, T.Y.; Rupiper, D.J. Exotic Animal Formulary, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2013; p. 46. [Google Scholar]
- Noga, E.J. Fish. Disease: Diagnosis and Treatment, 2nd ed.; Wiley Blackwell: Ames, IA, USA, 2010; p. 410. [Google Scholar]
- Lowry, O.H.; Rosenbrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Worthington, V. Worthington Enzyme Manual. Enzymes and Related Biochemicals; Worthington Chemical: Lakewood, NJ, USA, 1993; p. 399. [Google Scholar]
- Houseman, J.G.; Downe, A.E.R. Cathepsin D-like activity in the posterior midgut of hemipteran insects. Comp. Biochem. Physiol. 1983, 75, 509–512. [Google Scholar] [CrossRef]
- Rungruangsak-Torrissen, K.; Moss, R.; Andresen, L.H.; Berg, A.; Waagbo, R. Different expressions of trypsin and chymotrypsin in relation to growth in Atlantic salmon (Salmo salar L.). Fish. Physiol. Biochem. 2006, 32, 7–23. [Google Scholar] [CrossRef]
- Tsybina, T.A.; Dunaevsky, Y.E.; Belozersky, M.A.; Zhuzhikov, D.P.; Oppert, B.; Elpidina, E.N. Digestive proteinases of yellow mealworm (Tenebrio molitor) larvae: Purification and characterization of a trypsin-like proteinase. Biochemistry 2005, 70, 300–305. [Google Scholar] [CrossRef]
- Bernfeld, P. Enzymes of starch degradation and synthesis. Adv. Enzymol. 1951, 12, 379–428. [Google Scholar]
- Mendels, M.; Weber, J. The product of cellulose in cellulase and their application. Adv. Chem. Ser. 1969, 95, 391–443. [Google Scholar]
- AOAC. Official Method of Analysis of AOAC International; Association of Official Analytical Chemists: Arlington, VA, USA, 1990; pp. 69–84. [Google Scholar]
- Rungruangsak-Torrissen, K. Digestive efficiency, growth and qualities of muscle and oocyte in Atlantic salmon (Salmo salar L.) fed on diets with krill meal as an alternative protein source. J. Food Biochem. 2007, 31, 509–540. [Google Scholar] [CrossRef]
- Supannapong, P.; Pimsalee, T.; A-komol, T.; Engkagul, A.; Kovitvadhi, U.; Kovitvadhi, S.; Rungruangsak-Torrissen, K. Digestive enzymes and in vitro digestibility of different species of phytoplankton for culture of the freshwater pearl mussel, Hyriopsis (Hyriopsis) bialatus. Aquac. Int. 2008, 16, 437–453. [Google Scholar] [CrossRef]
- Thongprajukaew, K.; Kovitvadhi, U.; Kovitvadhi, S.; Somsueb, P.; Rungruangsak-Torrissen, K. Effects of different modified diets on growth, digestive enzyme activities and muscle compositions in juvenile Siamese fighting fish (Betta splendens Regan, 1910). Aquaculture 2011, 322–323, 1–9. [Google Scholar] [CrossRef]
- Rungruangsak-Torrissen, K.; Rustad, A.; Sunde, J.; Eiane, S.A.; Jensen, H.B.; Opstvedt, J.; Nygård, E.; Samuelsen, T.A.; Mundheim, H.; Luzzana, U.; et al. In vitro digestibility based on fish crude enzyme extract for prediction of feed quality in growth trials. J. Sci. Food Agric. 2002, 82, 644–654. [Google Scholar] [CrossRef]
- Özsoy, A.N. Modeling of development and water consumption of mealworm (Tenebrio molitor L., 1758) (Coleoptera: Tenebrionidae) larvae using nonlinear growth curves and polynomial functions. Turk. J. Entomol. 2019, 43, 253–262. [Google Scholar] [CrossRef]
- Beanland, L.; Phelan, P.L.; Salminen, S. Micronutrient interactions on soybean growth and the developmental performance of three insect herbivores. Environ. Entomol. 2003, 32, 641–651. [Google Scholar] [CrossRef]
- Yadav, R.; Chang, N.-T. Effects of temperature on the development and population growth of the melon thrips, Thripspalmi, on eggplant, Solanum melongena. J. Insect Sci. 2013, 14, 78. [Google Scholar]
- Padilha, M.P.H.; Pimentel, A.C.; Ribeiro, A.F.; Terra, W.R. Sequence and function of lysosomal and digestive cathepsin D-like proteinases of Musca domestica midgut. Insect Biochem. Mol. Biol. 2009, 39, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Elorduy, J.; Avila Gonzalez, E.; Rocha Hernandez, A.; Pino, J.M. Use of Tenebrio molitor (Coleoptera: Tenebrionidae) to recycle organic wastes and as feed for broiler chickens. J. Econ. Entomol. 2002, 95, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.; Fitzpatrick, M.P.; Dierenfeld, E.S. Nutrient composition of selected whole invertebrates. Zoo Biol. 1998, 17, 123–134. [Google Scholar] [CrossRef]
- Jones, L.D.; Cooper, R.W.; Harding, R.S. Composition of mealworm Tenebrio molitor larvae. J. Zoo Anim. Med. 1972, 3, 34–41. [Google Scholar] [CrossRef]
- Paul, A.; Frederich, M.; Megido, R.C.; Alabi, T.; Malik, P.; Uyttenbroeck, R.; Francis, F.; Blecker, C.; Haubruge, E.; Lognay, G.; et al. Insect fatty acids: A comparison of lipids from three Orthopterans and Tenebrio molitor L. larvae. J. Asia-Pac. Entomol. 2017, 20, 337–340. [Google Scholar] [CrossRef]
- Ravzanaadii, N.; Kim, S.H.; Choi, W.H.; Hong, S.J.; Kim, N.J. Nutritional value of mealworm, Tenebrio molitor as food source. Int. J. Ind. Entomol. 2012, 25, 93–98. [Google Scholar] [CrossRef]
- Zhao, X.; Vázquez-Gutiérrez, J.L.; Johansson, D.P.; Landberg, R.; Langton, M. Yellow mealworm protein for food purposes-Extraction and functional properties. PLoS ONE 2016, 11, e0147791. [Google Scholar] [CrossRef]
- Adámková, A.; Kouřimská, L.; Borkovcová, M.; Kulma, M.; Mlček, J. Nutritional values of edible coleoptera (Tenebrio molitor, Zophobas morio and Alphitobius diaperinus) reared in the Czech Republic. Potravin. Slovak. J. Food Sci. 2016, 10, 663–671. [Google Scholar]
- Song, Y.S.; Kim, M.W.; Moon, C.; Seo, D.J.; Han, Y.S.; Jo, Y.H.; Noh, M.Y.; Park, Y.K.; Kim, S.A.; Kim, Y.W.; et al. Extraction of chitin and chitosan from larval exuvium and whole body of edible mealworm, Tenebrio molitor. Entomol. Res. 2018, 48, 227–233. [Google Scholar] [CrossRef]
- Pascini, T.V.; Albeny, D.S.; Ramalho-Ortigão, M.; Vilela, E.F.; Serrão, J.E.; Martins, G.F. Changes in the fat body during the post-embryonic development of the predator Toxorhynchites theobaldi (Dyar & Knab) (Diptera: Culicidae). Neotrop. Entomol. 2011, 40, 456–461. [Google Scholar]
- Thongprajukaew, K.; Rodjaroen, S.; Tantikitti, C.; Kovitvadhi, U. Physicochemical modifications of dietary palm kernel meal affect growth and feed utilization of Nile tilapia (Oreochromis niloticus). Anim. Feed Sci. Technol. 2015, 202, 90–99. [Google Scholar] [CrossRef]
- Chumwaengwapee, S.; Soontornchai, S.; Thongprajukaew, K. Improving chemical composition, physicochemical properties, and in vitro carbohydrate digestibility of fish coconut meal. ScienceAsia 2013, 39, 636–642. [Google Scholar] [CrossRef]
- Bassompierre, M.; Ostenfeld, T.H.; McLean, E.; Rungruangsak-Torrissen, K. In vitro protein digestibility, and growth of Atlantic salmon with different trypsin isozymes. Aquac. Int. 1998, 6, 47–56. [Google Scholar] [CrossRef]
- Areekijseree, M.; Engkagul, A.; Kovitvadhi, S.; Kovitvadhi, U.; Thongpan, A.; RungruangsakTorrissen, K. Development of digestive enzymes and in vitro digestibility of different species of phytoplankton for culture of early juveniles of the freshwater pearl mussel, Hyriopsis (Hyriopsis) bialatus Simpson, 1900. Invert. Reprod. Dev. 2006, 49, 255–262. [Google Scholar] [CrossRef]
- Fontes, T.V.; Oliveira, K.R.B.; Almeida, I.L.G.; Orlando, T.M.; Rodrigues, P.B.; da Costa, D.V.; Rosa, P.V. Digestibility of insect meals for Nile tilapia fingerlings. Animals 2019, 9, 181. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Digestive Enzyme | Pepsin | Trypsin | Chymotrypsin | Amylase | Cellulase |
|---|---|---|---|---|---|
| Pepsin | 1 | ||||
| Trypsin | 0.761 ** | 1 | |||
| Chymotrypsin | 0.199 | 0.205 | 1 | ||
| Amylase | 0.417 ** | 0.284 | 0.103 | 1 | |
| Cellulase | –0.199 | –0.159 | 0.244 | 0.187 | 1 |
| Age (DAH) | Moisture | Crude Protein | Crude Lipid | Ash |
|---|---|---|---|---|
| 30 | 64.37 ± 0.83 a | 17.43 ± 3.34 e | 9.85 ± 0.17 e | 1.96 ± 0.06 a,b |
| 35 | 62.64 ± 1.24 a,b | 17.90 ± 1.82 d,e | 10.64 ± 1.21 d,e | 2.37 ± 0.55 a |
| 40 | 60.89 ± 0.24 b,c | 19.14 ± 0.65 d,e | 16.19 ± 0.98 a,b,c | 2.37 ± 0.28 a |
| 45 | 59.03 ± 0.17 c,d | 20.70 ± 1.04 c,d,e | 17.92 ± 2.53 a | 1.82 ± 0.08 b,c |
| 50 | 58.35 ± 0.46 d,e | 19.47 ± 0.61 d,e | 15.51 ± 1.43 a,b,c,d | 1.35 ± 0.11 c |
| 55 | 56.56 ± 0.44 e,f | 20.86 ± 0.92 c,d,e | 14.02 ± 0.70 a,b,c,d | 1.58 ± 0.08 b,c |
| 60 | 54.83 ± 0.42 f,g | 26.79 ± 0.45 a,b | 14.00 ± 2.18 a,b,c,d,e | 1.65 ± 0.04 b,c |
| 65 | 55.80 ± 1.38 f,g | 24.38 ± 1.91 a,b,c | 15.12 ± 0.75 a,b,c,d | 1.43 ± 0.09 c |
| 70 | 54.74 ± 1.41 f,g | 25.85 ± 2.25 a,b,c | 17.32 ± 0.50 a,b | 1.70 ± 0.10 b,c |
| 75 | 55.51 ± 0.79 f,g | 29.80 ± 1.05 a | 13.64 ± 2.07 a,b,c,d | 1.59 ± 0.05 b,c |
| 80 | 53.40 ± 0.43 g | 30.49 ± 2.76 a | 14.22 ± 0.20 a,b,c,d,e | 1.60 ± 0.11 b,c |
| 85 | 55.74 ± 0.32 f,g | 26.08 ± 1.45 a,b,c | 12.87 ± 0.16 b,c,d | 1.36 ± 0.14 c |
| 90 | 55.27 ± 0.69 f,g | 29.05 ± 0.97 a | 18.03 ± 2.28 a | 1.73 ± 0.06 b,c |
| p-value | <0.001 | <0.001 | 0.007 | 0.002 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodjaroen, S.; Thongprajukaew, K.; Khongmuang, P.; Malawa, S.; Tuntikawinwong, K.; Saekhow, S. Ontogenic Development of Digestive Enzymes in Mealworm Larvae (Tenebrio molitor) and Their Suitable Harvesting Time for Use as Fish Feed. Insects 2020, 11, 393. https://doi.org/10.3390/insects11060393
Rodjaroen S, Thongprajukaew K, Khongmuang P, Malawa S, Tuntikawinwong K, Saekhow S. Ontogenic Development of Digestive Enzymes in Mealworm Larvae (Tenebrio molitor) and Their Suitable Harvesting Time for Use as Fish Feed. Insects. 2020; 11(6):393. https://doi.org/10.3390/insects11060393
Chicago/Turabian StyleRodjaroen, Somrak, Karun Thongprajukaew, Puridet Khongmuang, Saowalak Malawa, Kimhun Tuntikawinwong, and Suktianchai Saekhow. 2020. "Ontogenic Development of Digestive Enzymes in Mealworm Larvae (Tenebrio molitor) and Their Suitable Harvesting Time for Use as Fish Feed" Insects 11, no. 6: 393. https://doi.org/10.3390/insects11060393
APA StyleRodjaroen, S., Thongprajukaew, K., Khongmuang, P., Malawa, S., Tuntikawinwong, K., & Saekhow, S. (2020). Ontogenic Development of Digestive Enzymes in Mealworm Larvae (Tenebrio molitor) and Their Suitable Harvesting Time for Use as Fish Feed. Insects, 11(6), 393. https://doi.org/10.3390/insects11060393