Drench Application of Systemic Insecticides Disrupts Probing Behavior of Diaphorina citri (Hemiptera: Liviidae) and Inoculation of Candidatus Liberibacter asiaticus

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and D. citri CLas-Infection
2.2. Insecticide Application
2.3. Electrical Penetration Graph (EPG) Recordings
2.4. Effects of Insecticides on CLas Transmission by D. citri
2.5. Detection of CLas by qPCR
2.5.1. DNA Extraction
2.5.2. qPCR Analysis
2.6. Statistical Analyses
3. Results
3.1. Probing Behavior of D. citri
3.2. Feeding Behavior and CLas Transmission
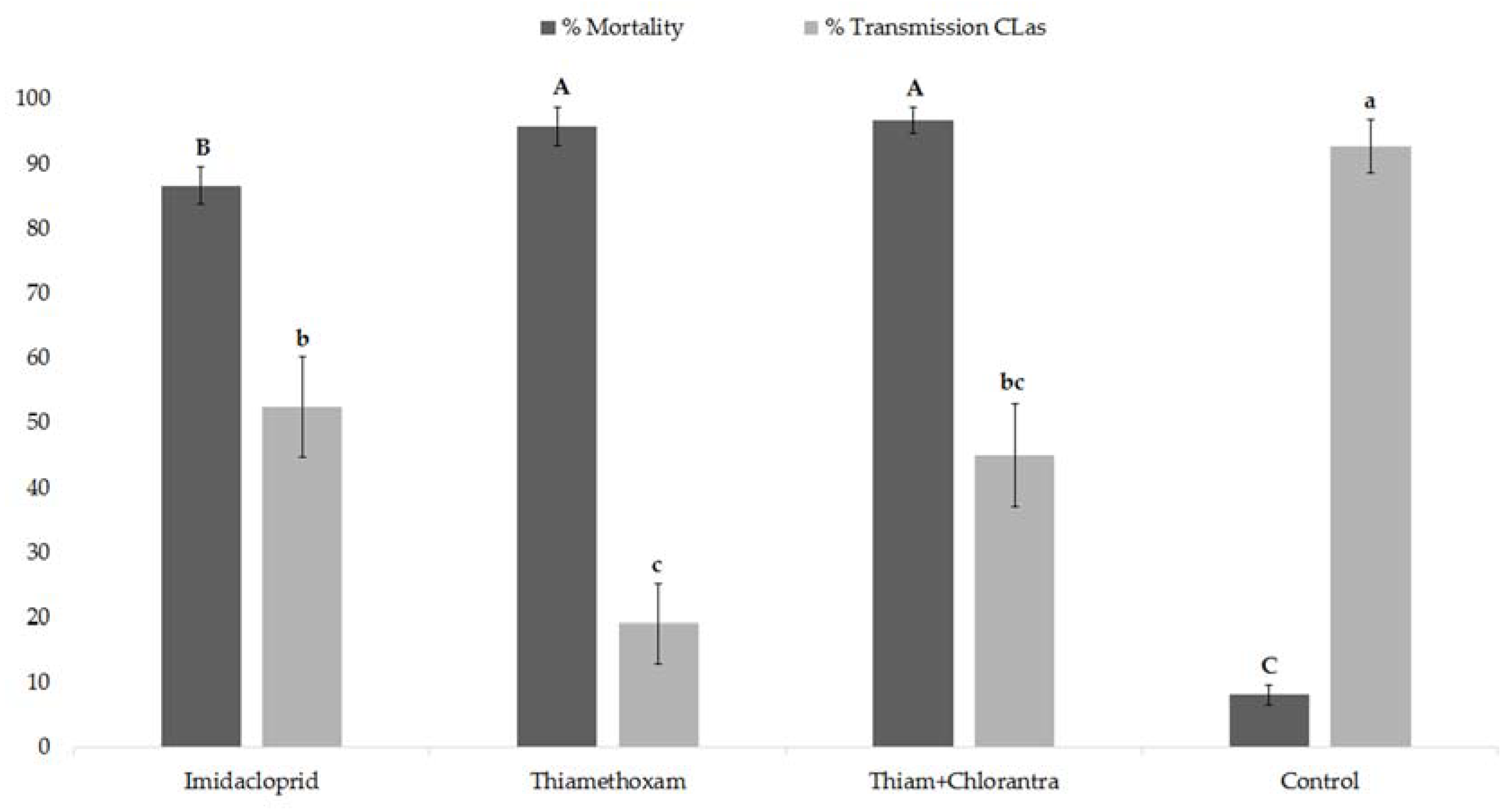
3.3. Insecticide Effects on Psyllid Mortality and CLas Transmission
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bové, J.M. Huanglongbing: A destructive, newly-emerging, century-old disease of citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar] [CrossRef]
- Coletta-Filho, H.D.; Targon, M.L.P.N.; Takita, M.A.; De Negri, J.D.; Pompeu, J., Jr.; Machado, M.A. First report of the causal agent of Huanglongbing (“Candidatus Liberibacter asiaticus”) in Brazil. Plant Dis. 2004, 88, 1382. [Google Scholar] [CrossRef]
- Teixeira, D.C.; Danet, J.L.; Eveillard, S.; Martins, E.C.; Jesus, W.C., Jr.; Yamamoto, P.T.; Lopes, S.A.; Bassanezi, R.B.; Ayres, A.J.; Saillard, C.; et al. Citrus huanglongbing in São Paulo State, Brazil: PCR detection of the ‘Candidatus’ Liberibacter species associated with the disease. Mol. Cell. Probes 2005, 19, 173–179. [Google Scholar] [CrossRef]
- Halbert, S. The discovery of huanglongbing in Florida. In Proceedings of the Second International Citrus Canker and Huanglongbing Research Workshop; Florida Citrus Mutual: Orlando, FL, USA, 2005; p. 50. [Google Scholar]
- Lopes, S.A.; Luiz, F.Q.B.F.; Martins, E.C.; Fassini, C.G.; Sousa, M.C.; Barbosa, J.C.; Beattie, G.A.C. Candidatus Liberibacter asiaticus titers in citrus and acquisition rates by Diaphorina citri are decreased by higher temperature. Plant Dis. 2013, 97, 1563–1570. [Google Scholar] [CrossRef] [Green Version]
- Fundo de Defesa da Citricultura. Sucesso Do Manejo Externo. Rev. Citric. 2019, 50, 12–15. Available online: https://www.fundecitrus.com.br/comunicacao/revista_detalhes/revista-citricultor---edicao-50/61 (accessed on 17 November 2019).
- Li, W.B.; Hartung, J.S.; Levy, L. Quantitative real-time PCR for detection and identification of Candidatus Liberibacter species associated with citrus huanglongbing. J. Microbiol. Methods 2006, 66, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Lopes, S.A.; Frare, G.F.; Bertolini, E.; Cambra, M.; Fernandes, N.G.; Ayres, A.J.; Marin, D.R.; Bové, J.M. Liberibacters associated with citrus huanglongbing in Brazil: ‘Candidatus Liberibacter asiaticus’ is heat tolerant, ‘Ca. L. americanus’ is heat sensitive. Plant Dis. 2009, 93, 257–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capoor, S.P.; Rao, D.G.; Viswanath, S.M. Diaphorina citri Kuwayama, a vector of the greening disease of citrus in India. Indian, J. Agric. Sci. 1967, 37, 572–576. [Google Scholar]
- Yamamoto, P.T.; Felippe, M.R.; Garbim, L.F.; Coelho, J.H.C.; Ximenes, N.L.; Martins, E.C.; Leite, A.P.R.; Sousa, M.C.; Abrahão, D.P.; Braz, J.D. Diaphorina citri (Kuwayama) (Hemiptera: Psyllydae): Vector of the bacterium Candidatus Liberibacter americanus. In Proceedings of Huanglongbing—Greening International Workshop; Fund for Citrus Protection: Ribeirão Preto/São Paulo, Brazil, 2006; p. 96. [Google Scholar]
- Inoue, H.; Ohnishi, J.; Ito, T.; Tomimura, K.; Miyata, S.; Iwanami, T.; Ashihara, W. Enhanced proliferation and efficient transmission of Candidatus Liberibacter asiaticus by adult Diaphorina citri after acquisition feeding in the nymphal stage. Ann. Appl. Biol. 2009, 155, 29–36. [Google Scholar] [CrossRef]
- Pelz-Stelinski, K.S.; Brlansky, R.H.; Ebert, T.A.; Rogers, M.E. Transmission parameters for Candidatus Liberibacter asiaticus by Asian citrus psyllid (Hemiptera: Psyllidae). J. Econ. Entomol. 2010, 103, 1531–1541. [Google Scholar] [CrossRef]
- Ammar, E.D.; Shatters, R.G.; Hall, D.G. Localization of Candidatus Liberibacter asiaticus, associated with citrus huanglongbing disease, in its psyllid vector using fluorescence in situ hybridization. J. Phytopathol. 2011, 159, 726–734. [Google Scholar] [CrossRef]
- Ammar, E.D.; Ramos, J.E.; Hall, D.G.; Dawson, W.O.; Shatters, R.G., Jr. Acquisition, replication and inoculation of Candidatus Liberibacter asiaticus following various acquisition periods on huanglongbing-infected citrus by nymphs and adults of the Asian citrus psyllid. PLoS ONE 2016, 11, e0159594. [Google Scholar] [CrossRef] [PubMed]
- Canale, M.C.; Tomaseto, A.F.; Haddad, M.L.; Coletta-Filho, H.D.; Lopes, J.R.S. Latency and persistence of ‘Candidatus Liberibacter asiaticus’ in its psyllid vector, Diaphorina citri (Hemiptera: Liviidae). Phytopathol. 2017, 107, 264–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, D.G.; Albrecht, U.; Bowman, K.D. Transmission rates of ‘Ca. Liberibacter asiaticus’ by asian citrus psyllid are enhanced by the presence and developmental stage of citrus flush. J. Econ. Entomol. 2016, 109, 558–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belasque, J., Jr.; Bassanezi, R.B.; Yamamoto, P.T.; Ayres, A.J.; Tachibana, A.; Violante, A.R.; Tank, A., Jr.; Di Giorgi, F.; Tersi, F.E.A.; Menezes, G.M.; et al. Lessons from huanglongbing management in São Paulo state, Brazil. J. Plant Pathol. 2010, 92, 285–302. [Google Scholar] [CrossRef]
- Miranda, M.P.; Volpe, H.X.L.; Veiga, A.C.; Zanardi, O.Z.; Bassanezi, R.B.; Andrade, D.J.; Carmo-Sousa, M. Manejo de insetos e ácaros vetores de fitopatógenos nos citros. Inf. Agropecuário (Belo Horizonte) 2017, 38, 83–93. [Google Scholar]
- McLean, D.L.; Kinsey, M.G. A technique for electronically recording aphid feeding and salivation. Nat. 1964, 202, 1358–1359. [Google Scholar] [CrossRef]
- Tjallingii, W.F. Electronic recording of penetration behaviour by aphids. Entomol. Exp. Appl. 1978, 24, 721–730. [Google Scholar] [CrossRef]
- Tjallingii, W.F. Electrical recording of stylet penetration activities. In Aphids, Their Biology, Natural Enemies and Control; Minks, A.K., Harrewijn, P., Eds.; Elsevier Science Publishers: Amsterdam, The Netherlands, 1988; Volume 1, pp. 95–108. [Google Scholar]
- Moreno-Delafuente, A.; Garzo, E.; Moreno, A.; Fereres, A. A plant virus manipulates the behavior of its whitefly vector to enhance its transmission efficiency and spread. PLoS ONE 2013, 8, e61543. [Google Scholar] [CrossRef]
- Backus, E.A.; Hunter, W.B. Comparison of feeding behavior of the potato leafhopper Empoasca fabae (Homoptera: Cicadellidae) on alfalfa and broad bean leaves. Environ. Entomol. 1989, 18, 473–480. [Google Scholar] [CrossRef]
- Miranda, M.P.; Fereres, A.; Appezzato-da-Gloria, B.; Lopes, J.R.S. Characterization of electrical penetration graphs of Bucephalogonia xanthophis, a vector of Xylella fastidiosa in citrus. Entomol. Exp. Appl. 2009, 130, 35–46. [Google Scholar] [CrossRef]
- Lucini, T.; Panizzi, A.R. Waveform characterization of the soybean stem feeder Edessa meditabunda: Overcoming the challenge of wiring pentatomids for EPG. Entomol. Exp. Appl. 2016, 158, 118–132. [Google Scholar] [CrossRef] [Green Version]
- Civolani, S.; Leis, M.; Grandi, G.; Garzo, E.; Pasqualini, E.; Musacchi, S.; Chicca, M.; Castaldelli, G.; Rossi, R.; Tjallingii, W.F. Stylet penetration of Cacopsylla pyri; an electrical penetration graph (EPG) study. J. Insect Physiol. 2011, 57, 1407–1419. [Google Scholar] [CrossRef] [PubMed]
- Pearson, C.C.; Backus, E.A.; Shugart, H.J.; Munyaneza, J.E. Feeding biology of the potato psyllid, Bactericera cockerelli. In Proceedings of the 10th Annual Zebra Chip Reporting Session; Texas A&M AgriLife Research: Dallas, TX, USA, 2010; pp. 178–182. [Google Scholar]
- Pearson, C.C.; Backus, E.A.; Shugart, H.J.; Munyaneza, J.E. Characterization and correlation of EPG waveforms of Bactericera cockerelli (Hemiptera: Triozidae): Variability in waveform appearance in relation to applied signal. Ann. Entomol. Soc. Am. 2014, 107, 650–666. [Google Scholar] [CrossRef] [Green Version]
- Sandanayaka, W.R.M.; Page-Weir, N.E.M.; Fereres, A. Real time EPG recordings of tomato-potato psyllid (Bactericera cockerelli) feeding on tomato. N. Z. Plant Prot. 2011, 64, 294. [Google Scholar] [CrossRef] [Green Version]
- Butler, C.D.; Walker, G.P.; Trumble, J.T. Feeding disruption of potato psyllid, Bactericera cockerelli, by imidacloprid as measured by electrical penetration graphs. Entomol. Exp. Appl. 2012, 142, 247–257. [Google Scholar] [CrossRef]
- Antolinez, C.A.; Moreno, A.; Appezzato-da-Gloria, B.; Fereres, A. Characterization of the electrical penetration graphs of the psyllid Bactericera trigonica on carrots. Entomol. Exp. Appl. 2017, 163, 127–139. [Google Scholar] [CrossRef]
- Bonani, J.P.; Fereres, A.; Garzo, E.; Miranda, M.P.; Appezzato-Da-Gloria, B.; Lopes, J.R.S. Caracterization of electrical penetration graphs of the Asian citrus psyllid, Diaphorina citri, in sweet orange seedlings. Entomol. Exp. Appl. 2010, 134, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Yen, A.L.; Powell, K.S.; Wu, F.; Wang, Y.; Zeng, L. Feeding behavior of Diaphorina citri (Hemiptera: Liviidae) and its acquisition of Candidatus Liberibacter asiaticus, on huanglongbing-infected Citrus reticulata leaves of several maturity stages. Fla. Entomol. 2015, 98, 186–192. [Google Scholar] [CrossRef]
- George, J.; Ammar, E.D.; Hall, D.G.; Shatters, R.G.; Lapointe, S.L. Prolonged phloem ingestion by Diaphorina citri nymphs compared to adults is correlated with increased acquisition of citrus greening pathogen. Sci. Rep. 2018, 8, 10352. [Google Scholar] [CrossRef] [PubMed]
- Sandanayaka, W.R.M.; Moreno, A.; Tooman, L.K.; Page-Weir, N.E.M.; Fereres, A. Stylet penetration activities linked to the acquisition and inoculation of Candidatus Liberibacter solanacearum by its vector tomato potato psyllid. Entomol. Exp. Appl. 2014, 151, 170–181. [Google Scholar] [CrossRef]
- Mustafa, T.; Alvarez, J.M.; Munyaneza, J.E. Effect of cyantraniliprole on probing behavior of the potato psyllid (Hemiptera: Triozidae) as measured by the electrical penetration graph technique. J. Econ. Entomol. 2015, 108, 2529–2535. [Google Scholar] [CrossRef] [PubMed]
- Butler, C.D.; Byrne, F.J.; Keremane, M.L.; Lee, R.F.; Trumble, J.T. Effects of insecticides on behavior of adult Bactericera cockerelli (Hemiptera: Triozidae) and transmission of Candidatus Liberibacter psyllaurous. J. Econ.Entomol. 2011, 104, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Serikawa, R.H.; Backus, E.A.; Rogers, M.E. Effects of soil-applied imidacloprid on Asian citrus psyllid (Hemiptera: Psyllidae) feeding behavior. J. Econ. Entomol. 2012, 105, 1492–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda, M.P.; Yamamoto, P.T.; Garcia, R.B.; Lopes, J.P.A.; Lopes, J.R.S. Thiamethoxam and imidacloprid drench applications on sweet orange nursery trees disrupt the feeding and settling behaviour of Diaphorina citri (Hemiptera: Liviidae). Pest Manag. Sci. 2016, 72, 1785–1793. [Google Scholar] [CrossRef]
- Sarriá, E.; Cid, M.; Garzo, E.; Fereres, A. Workbook for automatic parameter calculation of EPG data. Comput. Electron. Agric. 2009, 67, 35–42. [Google Scholar] [CrossRef]
- Van Helden, M.; Tjallingii, W.F. Experimental design and analysis in EPG experiments with emphasis on plant resistance research. In Principles and Applications of Electronic Monitoring and Other Techniques in the Study of Homopteran Feeding Behavior; Walker, G.P., Backus, E.A., Eds.; Entomological Society of America: Lanham, MD, USA, 2000; Volume 1, pp. 144–171. [Google Scholar]
- Backus, E.A.; Cline, A.R.; Ellerseick, M.R.; Serrano, M.S. Lygus Hesperus (Hemiptera: Miridae) feeding on cotton: New methods and parameters for analysis of non-sequential electrical penetration graph data. Ann. Entomol. Soc. Am. 2007, 100, 296–310. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, A.J.; Glower, J.C. The use of a multiple-transfer method in plant virus transmission studies-some statistical points arising in the analysis of results. Ann. Appl. Biol. 1960, 48, 75–83. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [Green Version]
- Hodkinson, I.D. The biology of the Psylloidea (Homoptera): A review. Bull. Entomol. Res. 1974, 64, 325–338. [Google Scholar] [CrossRef]
- Sétamou, M.; Simpson, C.R.; Alabi, O.J.; Nelson, S.D.; Telagamsetty, S.; Jifon, J.L. Quality matters: Influences of citrus flush physicochemical characteristics on population dynamics of the asian citrus psyllid (Hemiptera: Liviidae). PLoS ONE 2016, 11, e0168997. [Google Scholar] [CrossRef] [PubMed]
- Bethke, J.A.; Blua, M.J.; Redak, R.A. Effect of selected insecticides on Homalodisca coagulata (Homoptera: Cicadellidae) and transmission of oleander leaf scorch in a greenhouse study. J. Econ. Entomol. 2001, 94, 1031–1036. [Google Scholar] [CrossRef] [PubMed]
- Redak, R.; Blua, M. Impact of layering control tactics on the spread of pierce’s disease by the glassy-winged sharpshooter. In Proceedings of the CDFA Pierce’s Disease Research Symposium; California Department of Food and Agricultur: Coronado, CA, USA, 2001; pp. 159–160. [Google Scholar]
- Bergamin Filho, A.; Inoue-Nagata, A.K.; Bassanezi, R.B.; Belasque, J., Jr.; Amorim, L.; Macedo, M.A.; Barbosa, J.C.; Willocquet, L. The importance of primary inoculum and area-wide disease management to crop health and food security. Food Secur. 2016, 8, 221–238. [Google Scholar] [CrossRef]
- Wu, T.; Luo, X.; Xu, C.; Wu, F.; Qureshi, J.A.; Cen, Y. Feeding behavior of Diaphorina citri and its transmission of ‘Candidatus Liberibacter asiaticus’ to citrus. Entomol. Exp. Appl. 2016, 161, 104–111. [Google Scholar] [CrossRef]
- Tjallingii, W.F. Salivary secretions by aphids interacting with proteins of phloem wound responses. J. Exp. Bot. 2006, 57, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Ammar, E.D.; Hall, D.G.; Alvarez, J.M. Effect of cyantraniliprole, a novel insecticide, on the inoculation of Candidatus liberibacter asiaticus associated with citrus huanglongbing by the Asian citrus psyllid (Hemiptera: Liviidae). J. Econ. Entomol. 2015, 108, 399–404. [Google Scholar] [CrossRef]
- Bassanezi, R.B.; Montesino, L.H.; Gimenes-Fernandes, N.; Yamamoto, P.T.; Gottwald, T.R.; Amorim, L.; Bergamin Filho, A. Efficacy of area-wide inoculum reduction and vector control on temporal progress of huanglongbing in young sweet orange plantings. Plant Dis. 2013, 97, 789–796. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| Waveform * | Imidacloprid | Thiamethoxam | Thiam+Chlorantra | Control | χ2 | df | p |
|---|---|---|---|---|---|---|---|
| C | 100 (44/44) a | 100 (43/43) a | 100 (40/40) a | 100 (45/45) a | - | - | - |
| D + E1 | 81.8 (36/44) a | 67.4 (29/43) ab | 47.5 (19/40) b | 82.2 (37/45) a | 16.002 | 3 | 0.0011 |
| E2 E2 >10 min | 77.3 (34/44) a 43.2 (19/44) ab | 65.1 (28/43) ab 32.6 (14/43) b | 47.5 (19/40) b 27.5 (11/40) b | 80 (36/45) a 75.6 (34/45) a | 12.611 8.564 | 3 3 | 0.0056 0.0357 |
| EPG Variables | Imidacloprid | Thiamethoxam | Thiam + Chlorantra | Control | H | d.f | p |
|---|---|---|---|---|---|---|---|
| NPI a,b | 7.48 ± 0.91 a | 6.09 ± 0.65 a | 5.58 ± 0.72 a | 6.07 ± 0.82 a | 3.6759 | 3 | 0.2987 |
| NWEI a,b | |||||||
| np | 8.32 ± 0.91 a | 6.98 ± 0.65 a | 6.45 ± 0.72 a | 6.29 ± 0.84 a | 6.8362 | 3 | 0.0773 |
| C | 8.45 ± 0.93 a | 6.81 ± 0.65 a | 6.18 ± 0.70 a | 6.47 ± 0.86 a | 6.5814 | 3 | 0.0865 |
| D + E1 | 1.02 ± 0.11 a | 0.74 ± 0.09 ab | 0.53 ± 0.09 b | 1.04 ± 0.11 a | 16.5233 | 3 | 0.0009 |
| E2 | 0.90 ± 0.09 a | 0.67 ± 0.08 ab | 0.50 ± 0.09 b | 0.98 ± 0.10 a | 14.8963 | 3 | 0.0019 |
| NE2 >10 min | 0.48 ± 0.09 b | 0.33 ± 0.07 b | 0.28 ± 0.07 b | 0.91 ± 0.09 a | 23.6247 | 3 | <0.0001 |
| G | 0.02 ± NA a | 0 a | 0.125 ± 0.09 a | 0.07 ± 0.05 a | 0.201 | 3 | 0.9774 |
| WDI a,b,c | |||||||
| np | 369.05 ± 11.32 a | 373.38 ± 15.82 a | 386.12 ± 15.08 a | 158.05 ± 21.48 b | 52.3094 | 3 | <0.0001 |
| C | 95.31 ± 11.00 a | 94.26 ± 15.60 a | 85.53 ± 15.30 a | 95.03 ± 14.73 a | 3.366 | 3 | 0.3386 |
| D | 0.68 ± 0.10 a | 0.40 ± 0.07 b | 0.27 ± 0.05 b | 0.60 ± 0.08 a | 18.0769 | 3 | 0.0004 |
| E1 | 1.74 ± 0.27 a | 1.02 ± 0.22 b | 0.94 ± 0.22 b | 2.01 ± 0.29 a | 15.0493 | 3 | 0.0018 |
| E2 | 13.13 ± 2.19 b | 10.93 ± 2.29 bc | 6.34 ± 1.42 c | 219.35 ± 23.87 a | 43.167 | 3 | <0.0001 |
| G | 0.08 ± 0.08 a | 0 a | 0.80 ± 0.72 a | 4.95 ± 4.50 a | 0.1988 | 3 | 0.9778 |
| WDE a,b,c | |||||||
| np | 44.20 ± 4.26 a | 53.52 ± 5.86 a | 59.86 ± 6.47 a | 25.13 ± 2.59 b | 14.109 | 3 | 0.003 |
| C | 11.27 ± 1.07 a | 13.83 ± 2.15 a | 13.85 ± 2.49 a | 14.69 ± 1.81 a | 4.118 | 3 | 0.249 |
| D | 0.67 ± 0.05 a | 0.54 ± 0.06 b | 0.52 ± 0.05 ab | 0.58 ± 0.06 b | 8.0433 | 3 | 0.0451 |
| E1 | 1.71 ± 0.24 a | 1.38 ± 0.26 a | 1.79 ± 0.32 a | 1.97 ± 0.21 a | 4.9619 | 3 | 0.1746 |
| E2 | 14.81 ± 2.11 b | 16.21 ± 2.90 b | 12.68 ± 1.98 b | 224.33 ± 20.88 a | 65.9484 | 3 | <0.0001 |
| E2 > 10 min | 21.98 ± 3.15 b | 27.32 ± 4.30 b | 18.19 ± 2.54 b | 240.41 ± 20.20 a | 61.2412 | 3 | <0.0001 |
| G | 3.36 ± NA | - | 6.39 ± 3.20 | 74.31 ± 63.95 | - | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carmo-Sousa, M.; Garcia, R.B.; Wulff, N.A.; Fereres, A.; Miranda, M.P. Drench Application of Systemic Insecticides Disrupts Probing Behavior of Diaphorina citri (Hemiptera: Liviidae) and Inoculation of Candidatus Liberibacter asiaticus. Insects 2020, 11, 314. https://doi.org/10.3390/insects11050314
Carmo-Sousa M, Garcia RB, Wulff NA, Fereres A, Miranda MP. Drench Application of Systemic Insecticides Disrupts Probing Behavior of Diaphorina citri (Hemiptera: Liviidae) and Inoculation of Candidatus Liberibacter asiaticus. Insects. 2020; 11(5):314. https://doi.org/10.3390/insects11050314
Chicago/Turabian StyleCarmo-Sousa, Michele, Rafael Brandão Garcia, Nelson Arno Wulff, Alberto Fereres, and Marcelo Pedreira Miranda. 2020. "Drench Application of Systemic Insecticides Disrupts Probing Behavior of Diaphorina citri (Hemiptera: Liviidae) and Inoculation of Candidatus Liberibacter asiaticus" Insects 11, no. 5: 314. https://doi.org/10.3390/insects11050314
APA StyleCarmo-Sousa, M., Garcia, R. B., Wulff, N. A., Fereres, A., & Miranda, M. P. (2020). Drench Application of Systemic Insecticides Disrupts Probing Behavior of Diaphorina citri (Hemiptera: Liviidae) and Inoculation of Candidatus Liberibacter asiaticus. Insects, 11(5), 314. https://doi.org/10.3390/insects11050314