Stable Isotope Enrichment (Δ15N) in the Predatory Flower Bug (Orius majusculus) Predicts Fitness-Related Differences between Diets


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Stable Isotope Analysis
2.3. Statistical Analysis
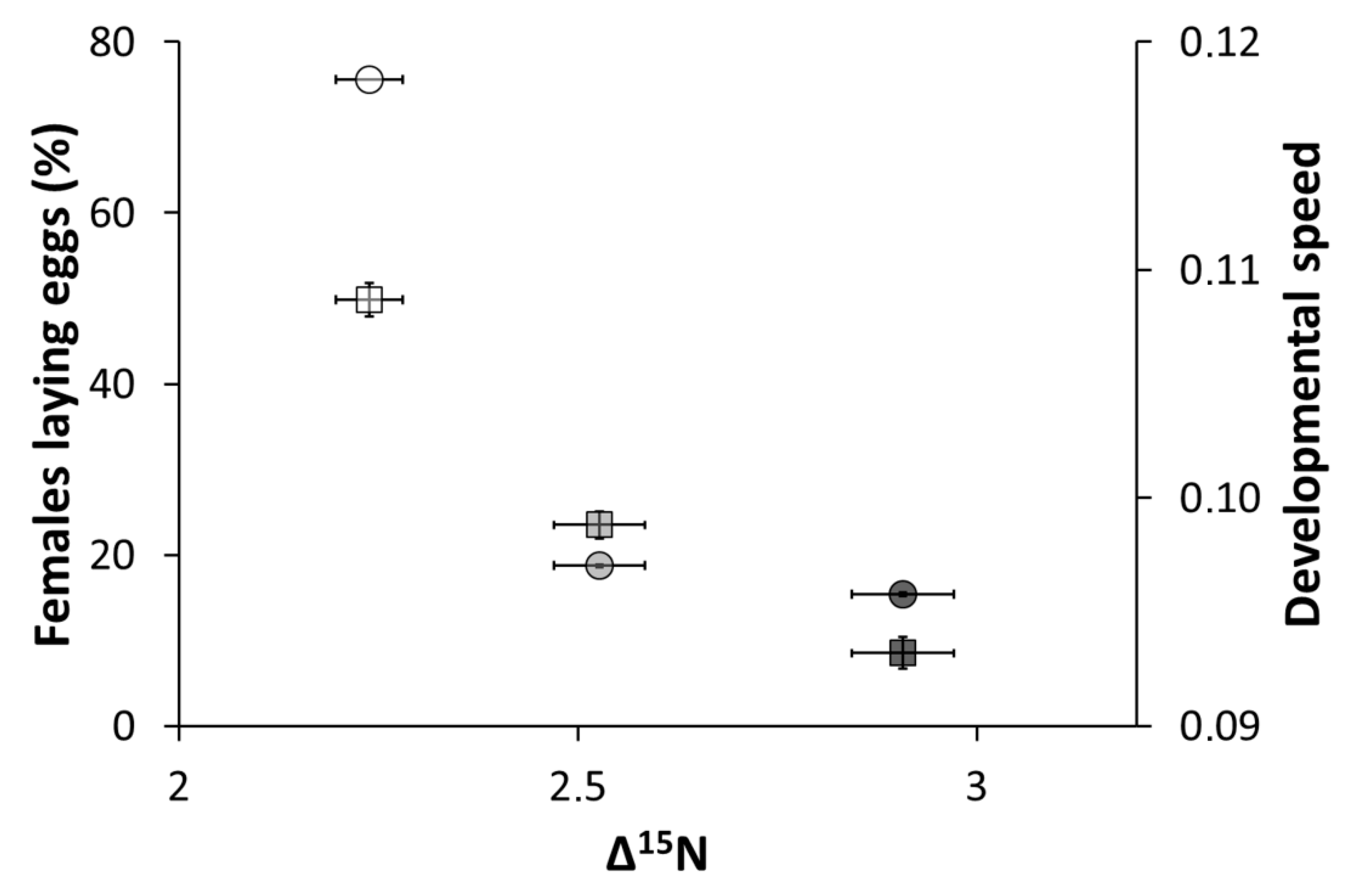
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ferkovich, S.M.; Shapiro, J.P. Increased egg-laying in Orius insidiosus (Hemiptera: Anthocoridae) fed artificial diet supplemented with an embryonic cell line. Biol. Control 2004, 31, 11–15. [Google Scholar] [CrossRef]
- De Clercq, P.; Arijs, Y.; Van Meir, T.; Van Stappen, G.; Sorgeloos, P.; Dewettinck, K.; Rey, M.; Grenier, S.; Febvay, G. Nutritional value of brine shrimp cysts as a factitious food for Orius laevigatus (Heteroptera: Anthocoridae). Biocontrol Sci. Techn. 2005, 15, 467–479. [Google Scholar] [CrossRef]
- Zapata, R.; Specty, O.; Grenier, S.; Febvay, G.; Pageaux, J.F.; Delobel, B.; Castañé, C. Carcass analysis to improve a meat-based diet for the artificial rearing of the predatory mirid bug Dicyphus tamaninii. Arch. Insect Biochem. 2005, 60, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Sighinolfi, L.; Febvay, G.; Dindo, M.L.; Rey, M.; Pageaux, J.; Baronio, P.; Grenier, S. Biological and biochemical characteristics for quality control of Harmonia axyridis (Pallas) (Coleoptera, Coccinellidae) reared on a liver-based diet. Arch. Insect Biochem. 2008, 68, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Schultz, H.; da Silva, E.; de Lima Aguiar-Menezes, E.; Santos Resende, A.L.; Ribeiro Costa Rouws, J.; Moreira da Silva, A.R. Adequacy of Drosophila melanogaster as prey for the development and reproduction of Coleomegilla maculata. BioControl 2019, 64, 43–54. [Google Scholar] [CrossRef]
- Montoro, M.; De Fine Licht, H.H.; Sigsgaard, L. Nutritional quality of Drosophila melanogaster as factitious prey for rearing the predatory bug Orius Majusculus. Insect Sci. 2020. [Google Scholar] [CrossRef]
- Montoro, M.; De Clercq, P.; Overgaard, J.; Sigsgaard, L. Fitness consequences of artificial diets with different macronutrient composition for the predatory bug, Orius majusculus. Entomol. Exp. Appl. 2020. [Google Scholar] [CrossRef]
- Specty, O.; Febvay, G.; Grenier, S.; Delobel, B.; Piotte, C.; Pageaux, J.F.; Ferran, A.; Guillaud, J. Nutritional plasticity of the predatory ladybeetle Harmonia axyridis (Coleoptera: Coccinellidae): Comparison between natural and substitution prey. Arch. Insect Biochem. 2003, 52, 81–91. [Google Scholar] [CrossRef]
- He, X.; Sigsgaard, L. A floral diet increases the longevity of the coccinellid Adalia bipunctata but does not allow molting or reproduction. Front. Ecol. Evol. 2019, 7, 1–11. [Google Scholar] [CrossRef]
- Cohen, A.C. Function of Insect Diet Components. In Insect diets, Science and Technology; Cohen, A.C., Ed.; CRC Press: Boca Raton, FL, USA, 2015; pp. 29–56. [Google Scholar]
- Grenier, S.; De Clercq, P. Comparison of artificially vs. naturally reared natural enemies and their potential for use in biological control. In Quality Control and Production of Biological Control Agents –Theory and Testing Procedures; van Lenteren, J.C., Ed.; CABI Publishing: Cambridge, UK, 2003; pp. 115–131. [Google Scholar]
- Dindo, M.L.; Grenier, S.; Sighinolfi, L.; Baronio, P. Biological and biochemical differences between in vitro- and in vivo-reared Exorista larvarum. Entomol. Exp. Appl. 2006, 120, 167–174. [Google Scholar] [CrossRef]
- Grenier, S. Artificial rearing of entomophagous insects, with emphasis on nutrition and parasitoids-General outlines from personal experience. Karaelmas Sci. Eng. J. 2012, 2, 1–12. [Google Scholar] [CrossRef][Green Version]
- Ferkovich, S.M.; Shapiro, J.P. Enhanced oviposition in the insidious flower bug, Orius majusculus (Hemiptera: Anthocoridae) with a partially purified nutritional factor from prey eggs. Fla. Entomol. 2005, 88, 253–257. [Google Scholar] [CrossRef]
- DeNiro, M.; Epstein, S. Influence of diet on the distribution of nitrogen isotopes in animals. Geochim. Cosmochim. Ac. 1981, 45, 341–351. [Google Scholar] [CrossRef]
- Kling, G.W.; Fry, B.; O’Brien, W.J. Stable isotopes and planktonic trophic structure in arctic lakes. Ecology 1992, 73, 561–566. [Google Scholar] [CrossRef]
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Gratton, C.; Forbes, A.E. Changes in δ13C stable isotopes in multiple tissues of insect predators fed isotopically distinct prey. Oecologia 2006, 147, 615–624. [Google Scholar] [CrossRef]
- Caut, S.; Angulo, E.; Courchamp, F. Variation in discrimination factors (Δ15N and Δ13C): The effect of diet isotopic values and applications for diet reconstruction. J. Appl. Ecol. 2009, 46, 443–453. [Google Scholar] [CrossRef]
- Peterson, B.; Fry, B. Stable isotopes in ecosystem studies. Annu. Rev. Ecol. Syst. 1987, 18, 293–320. [Google Scholar] [CrossRef]
- Jensen, P.M.; Madsen, P.; Jensen, L.S.; Pipper, C.B. Differences in carbon and nitrogen stable isotope signatures amongst wild and released pheasant populations. Eur. J. Wildlife Res. 2012, 58, 755–760. [Google Scholar] [CrossRef]
- Webb, S.C.; Hedges, R.E.M.; Simpson, S.J. Diet quality influences the δ13C and δ15N of locusts and their biochemical components. J. Exp. Biol. 1998, 201, 2903–2911. [Google Scholar]
- Ben-David, M.; Flaherty, E.A. Stable isotopes in mammalian research: A beginner’s guide. J. Mammal. 2012, 93, 312–328. [Google Scholar] [CrossRef]
- Oelbermann, K.; Scheu, S. Stable isotope enrichment (δ15N and δ13C) in a generalist predator (Pardosa lugubris, Araneae: Lycosidae): Effects of prey quality. Oecologia 2002, 130, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Jensen, K.; Mayntz, D.; Wang, T.; Simpson, S.J.; Overgaard, J. Metabolic consequences of feeding and fasting on nutritionally different diets in the wolf spider Pardosa prativaga. J. Insect Physiol. 2010, 56, 1095–1100. [Google Scholar] [CrossRef] [PubMed]
- UCDavis Stable Isotope Facility. Available online: https://stableisotopefacility.ucdavis.edu/ (accessed on 6 April 2020).
- R Core Team. R: A Language and Environment for Statistical Computing. Version 3.2.5 [Software]. Available online: http://www.R-project.org/ (accessed on 15 January 2020).
- Markow, T.A.; Anwar, S.; Pfeiler, E. Stable isotope ratios of carbon and nitrogen in natural populations of Drosophila species and their hosts. Funct. Ecol. 2000, 14, 261–266. [Google Scholar] [CrossRef]
- Adams, T.S.; Sterner, R.W. The effect of dietary nitrogen content on trophic level 15N enrichment. Limnol. Oceanogr. 2000, 45, 601–607. [Google Scholar] [CrossRef]
- Arostegui, M.C.; Schindler, D.E.; Holtgrieve, G.W. Does lipid-correction introduce biases into isotopic mixing models? Implications for diet reconstruction studies. Oecologia 2019, 191, 745–755. [Google Scholar] [CrossRef]
- Robbins, C.T.; Felicetti, L.A.; Florin, S.T. The impact of protein quality on stable nitrogen isotope ratio discrimination and assimilated diet estimation. Oecologia 2010, 162, 571–579. [Google Scholar] [CrossRef]


{kind=link}
| Diet | Parameters | |||
|---|---|---|---|---|
| δ15N (‰) | δ13C (‰) | Δ15N (‰) | Δ13C (‰) | |
| E. kuehniella | 6.40 ± 0.04a | −28.38 ± 0.06a | 2.24 ± 0.04c | 0.64 ± 0.06a |
| P-rich flies | 6.06 ± 0.06a | −23.46 ± 0.04b | 2.53 ± 0.06b | −0.02 ± 0.06b |
| L-rich flies | 2.52 ± 0.06b | −23.91 ± 0.06b | 2.91 ± 0.06a | 0.44 ± 0.04a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montoro, M.; Jensen, P.M.; Sigsgaard, L. Stable Isotope Enrichment (Δ15N) in the Predatory Flower Bug (Orius majusculus) Predicts Fitness-Related Differences between Diets. Insects 2020, 11, 255. https://doi.org/10.3390/insects11040255
Montoro M, Jensen PM, Sigsgaard L. Stable Isotope Enrichment (Δ15N) in the Predatory Flower Bug (Orius majusculus) Predicts Fitness-Related Differences between Diets. Insects. 2020; 11(4):255. https://doi.org/10.3390/insects11040255
Chicago/Turabian StyleMontoro, Marta, Per M. Jensen, and Lene Sigsgaard. 2020. "Stable Isotope Enrichment (Δ15N) in the Predatory Flower Bug (Orius majusculus) Predicts Fitness-Related Differences between Diets" Insects 11, no. 4: 255. https://doi.org/10.3390/insects11040255
APA StyleMontoro, M., Jensen, P. M., & Sigsgaard, L. (2020). Stable Isotope Enrichment (Δ15N) in the Predatory Flower Bug (Orius majusculus) Predicts Fitness-Related Differences between Diets. Insects, 11(4), 255. https://doi.org/10.3390/insects11040255