Host Plants Influence the Symbiont Diversity of Eriosomatinae (Hemiptera: Aphididae)


Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and DNA Extraction
2.2. Amplification and Sequencing of the 16S Ribosomal RNA Gene
2.3. Sequencing Data Analysis
2.4. Symbiont Community Analysis
3. Results
3.1. Sequencing Data
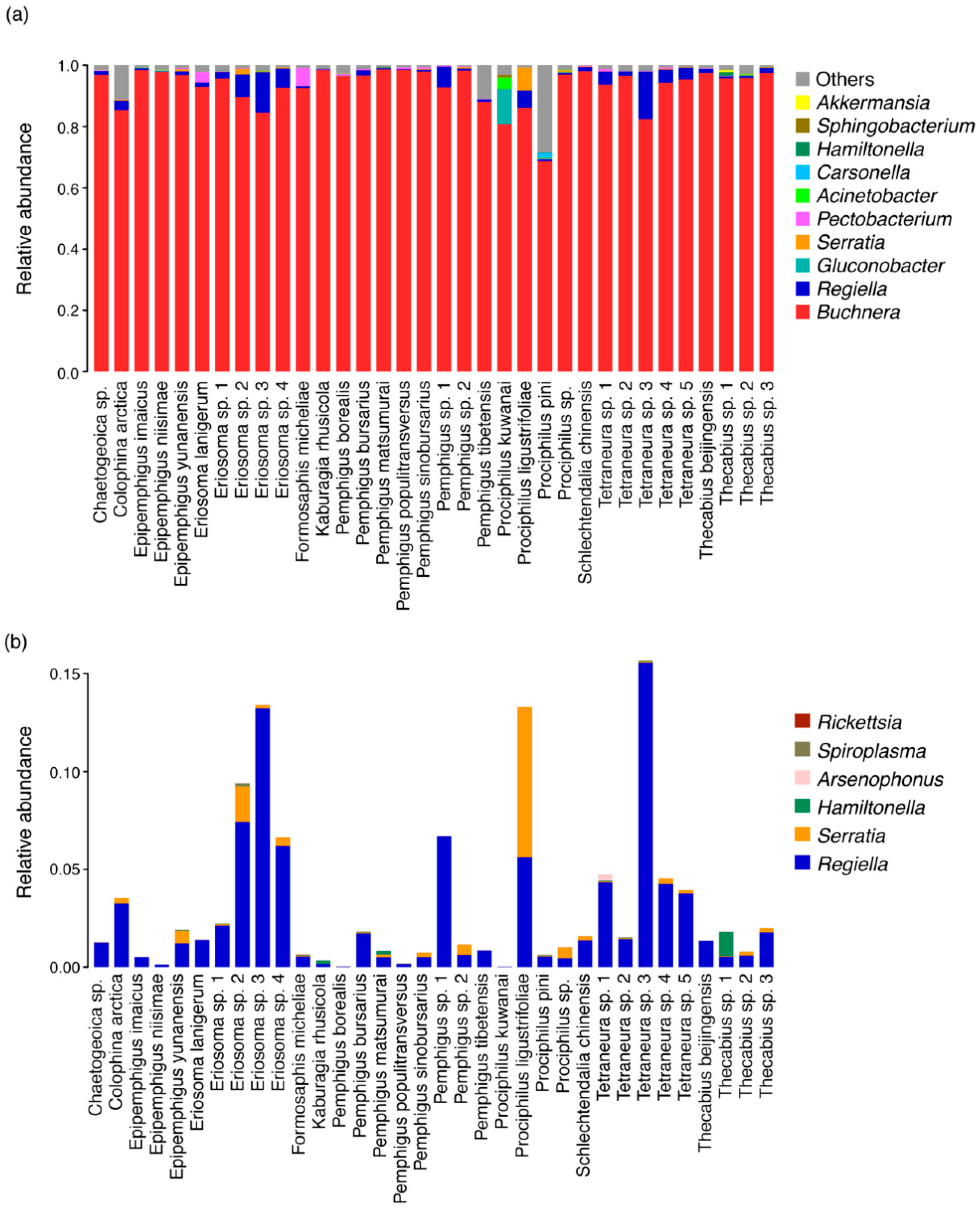
3.2. Bacterial Diversity across Eriosomatinae Aphids
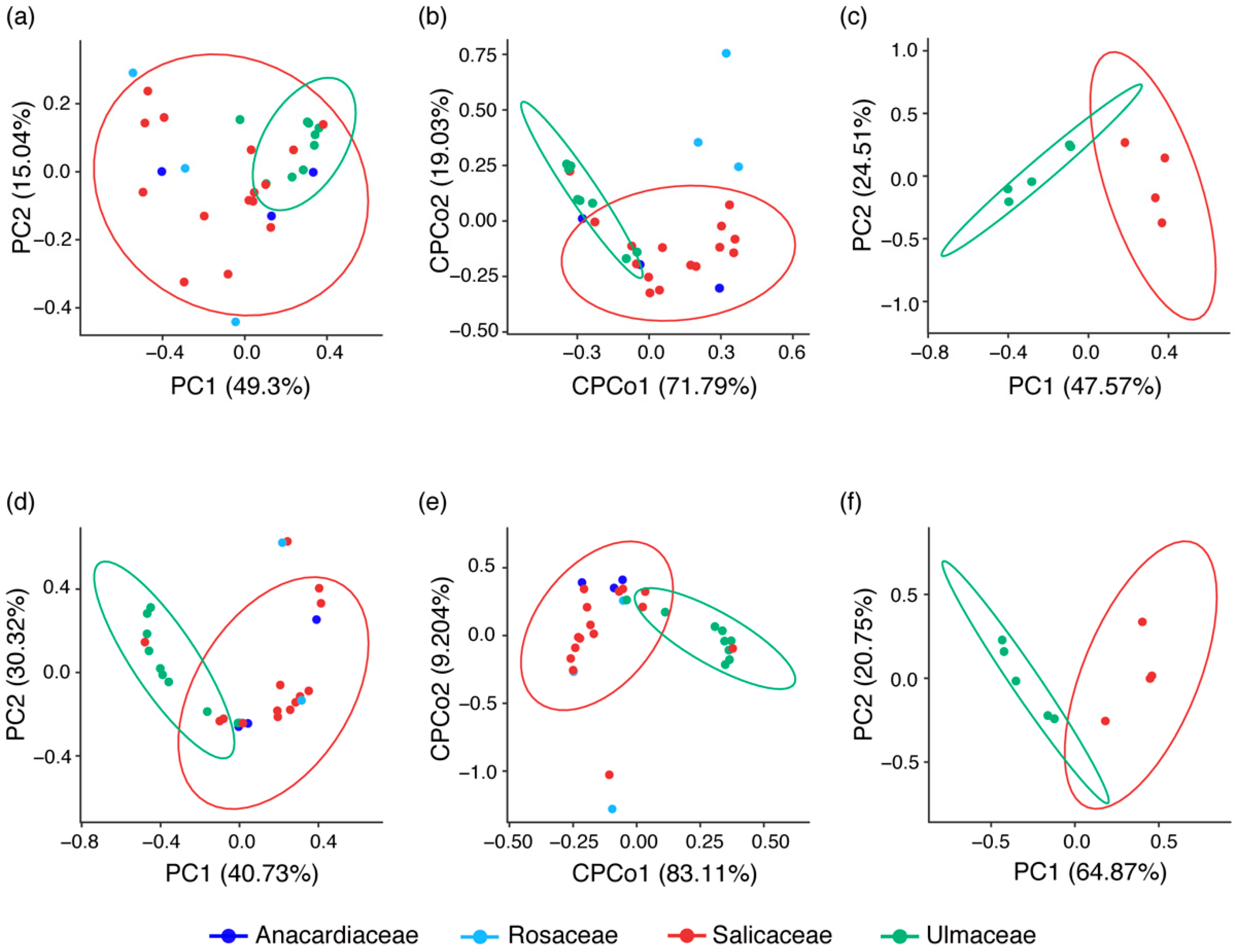
3.3. Comparison of Microbiota Associated with Eriosomatinae among Plants, Geographic Distributions and Aphid Relatedness
3.4. Correlation Test between Different Symbionts Associated with Eriosomatinae Aphids
4. Discussion
4.1. Diversity and Composition of Eriosomatinae Aphid Symbionts
4.2. Structure of Symbiont Community in Relation to Ecological Conditions
4.3. Interactions between Symbionts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buchner, P. Endosymbiosis of Animals with Plant. Microorganisms; Interscience Publishers: New York, NY, USA, 1965. [Google Scholar]
- Douglas, A.E.; Prosser, W.A. Synthesis of the essential amino-acid tryptophan in the pea aphid (Acyrthosiphon pisum) symbiosis. J. Insect. Physiol. 1992, 38, 565–568. [Google Scholar] [CrossRef]
- Douglas, A.E. Nutritional interactions in insect-microbial symbioses: Aphids and their symbiotic bacteria Buchnera. Annu. Rev. Entomol. 1998, 43, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Shigenobu, S.; Watanabe, H.; Hattori, M.; Sakaki, Y.; Ishikawa, H. Genome sequence of the endocellular bacterial symbiont of aphids Buchnera sp. Aps. Nature 2000, 407, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Munson, M.A.; Baumann, P.; Clark, M.A.; Baumann, L.; Moran, N.A.; Voegtlin, D.J.; Campbell, B.C. Evidence for the establishment of aphid-eubacterium endosymbiosis in an ancestor of four aphid families. J. Bacteriol. 1991, 173, 6321–6324. [Google Scholar] [CrossRef] [PubMed]
- Moran, N.A.; Munson, M.A.; Baumann, P.; Ishikawa, H. A molecular clock in endosymbiotic bacteria is calibrated using the insect hosts. Proc. R. Soc. B 1993, 253, 167–171. [Google Scholar] [CrossRef]
- Baumann, P.; Baumann, L.; Lai, C.Y.; Rouhbakhsh, D.; Moran, N.A.; Clark, M.A. Genetics, physiology, and evolutionary relationships of the genus Buchnera: Intracellular symbionts of aphids. Annu. Rev. Microbiol. 1995, 49, 55–94. [Google Scholar] [CrossRef]
- Baumann, P.; Moran, N.A.; Baumann, L. The evolution and genetics of aphid endosymbionts. Bioscience 1997, 47, 12–20. [Google Scholar] [CrossRef]
- Clark, M.A.; Moran, N.A.; Baumann, P.; Wernegreen, J.J. Cospeciation between bacterial endosymbionts (Buchnera) and a recent radiation of aphids (Uroleucon) and pitfalls of testing for phylogenetic congruence. Evolution 2000, 54, 517–525. [Google Scholar] [CrossRef]
- Jousselin, E.; Desdevises, Y.; Coeur d’acier, A. Fine-scale cospeciation between Brachycaudus and Buchnera aphidicola: Bacterial genome helps define species and evolutionary relationships in aphids. Proc. R. Soc. B 2009, 276, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Huang, X.L.; Zhang, R.L.; Jiang, L.Y.; Qiao, G.X. Phylogenetic congruence between Mollitrichosiphum (Aphididae: Greenideinae) and Buchnera indicates insect-bacteria parallel evolution. Syst. Entomol. 2013, 38, 81–92. [Google Scholar] [CrossRef]
- Liu, L.; Li, X.Y.; Huang, X.L.; Qiao, G.X. Evolutionary relationships of Pemphigus and allied genera (Hemiptera: Aphididae: Eriosomatinae) and their primary endosymbiont, Buchnera aphidicola. Insect Sci. 2014, 21, 301–312. [Google Scholar] [CrossRef]
- Xu, T.T.; Chen, J.; Jiang, L.Y.; Qiao, G.X. Historical and cospeciating associations between Cerataphidini aphids (Hemiptera: Aphididae: Hormaphidinae) and their primary endosymbiont Buchnera aphidicola. Zool. J. Linn. Soc. 2018, 183, 604–613. [Google Scholar] [CrossRef]
- Chen, D.Q.; Purcell, A.H. Occurrence and transmission of facultative endosymbionts in aphids. Curr. Microbiol. 1997, 34, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Sandström, J.P.; Russell, J.A.; White, J.P.; Moran, N.A. Independent origins and horizontal transfer of bacterial symbionts of aphids. Mol. Ecol. 2001, 10, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.A.; Latorre, A.; Sabater-Munoz, B.; Moya, A.; Moran, N.A. Side-stepping secondary symbionts: Widespread horizontal transfer across and beyond the Aphidoidea. Mol. Ecol. 2003, 12, 1061–1075. [Google Scholar] [CrossRef]
- Russell, J.A.; Moran, N.A. Horizontal transfer of bacterial symbionts: Heritability and fitness effects in a novel aphid host. Appl. Environ. Microbiol. 2005, 71, 7987–7994. [Google Scholar] [CrossRef]
- Vorburger, C.; Siegrist, G.; Rhyner, N. Faithful vertical transmission but ineffective horizontal transmission of bacterial endosymbionts during sexual reproduction of the black bean aphid, Aphis fabae. Ecol. Entomol. 2017, 42, 202–209. [Google Scholar] [CrossRef]
- Rock, D.I.; Smith, A.H.; Joffe, J.; Albertus, A.; Wong, N.; O’Connor, M.; Oliver, K.M.; Russell, J.A. Context-dependent vertical transmission shapes strong endosymbiont community structure in the pea aphid, Acyrthosiphon pisum. Mol. Ecol. 2018, 27, 2039–2056. [Google Scholar] [CrossRef]
- Oliver, K.M.; Russell, J.A.; Moran, N.A.; Hunter, M.S. Facultative bacterial symbionts in aphids confer resistance to parasitic wasps. Proc. Natl. Acad. Sci. USA 2003, 100, 1803–1807. [Google Scholar] [CrossRef]
- Oliver, K.M.; Moran, N.A.; Hunter, M.S. Variation in resistance to parasitism in aphids is due to symbionts not host genotype. Proc. Natl. Acad. Sci. USA 2005, 102, 12795–12800. [Google Scholar] [CrossRef]
- Vorburger, C.; Gehrer, L.; Rodriguez, P. A strain of the bacterial symbiont Regiella insecticola protects aphids against parasitoids. Biol. Lett. 2010, 6, 109–111. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.K.; Vorburger, C.; Moran, N.A. Genomic basis of endosymbiont-conferred protection against an insect parasitoid. Genome. Res. 2012, 22, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Brandt, J.W.; Chevignon, G.; Oliver, K.M.; Strand, M.R. Culture of an aphid heritable symbiont demonstrates its direct role in defence against parasitoids. Proc. R. Soc. B 2017, 284, 20171925. [Google Scholar] [CrossRef] [PubMed]
- Frago, E.; Mala, M.; Weldegergis, B.T.; Yang, C.; McLean, A.; Godfray, H.C.J.; Gols, R.; Dicke, M. Symbionts protect aphids from parasitic wasps by attenuating herbivore-induced plant volatiles. Nat. Commun. 2017, 8, 1860. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, J.; Darby, A.C.; Daniell, T.J.; Godfray, H.C.J.; Douglas, A.E. Linking the bacterial community in pea aphids with host-plant use and natural enemy resistance. Ecol. Entomol. 2004, 29, 60–65. [Google Scholar] [CrossRef]
- Scarborough, C.L.; Ferrari, J.; Godfray, H.C. Aphid protected from pathogen by endosymbiont. Science 2005, 310, 1781. [Google Scholar] [CrossRef]
- Łukasik, P.; van Asch, M.; Guo, H.; Ferrari, J.; Godfray, H.C. Unrelated facultative endosymbionts protect aphids against a fungal pathogen. Ecol. Lett. 2013, 16, 214–218. [Google Scholar] [CrossRef]
- Chen, D.Q.; Montllor, C.B.; Purcell, A.H. Fitness effects of two facultative endosymbiotic bacteria on the pea aphid, Acyrthosiphon pisum, and the blue alfalfa aphid, A. Kondoi. Entomol. Exp. Appl. 2000, 95, 315–323. [Google Scholar] [CrossRef]
- Montllor, C.B.; Maxmen, A.; Purcell, A.H. Facultative bacterial endosymbionts benefit pea aphids Acyrthosiphon pisum under heat stress. Ecol. Entomol. 2002, 27, 189–195. [Google Scholar] [CrossRef]
- Leonardo, T.E.; Muiru, G.T. Facultative symbionts are associated with host plant specialization in pea aphid populations. Proc. R. Soc. Lond. B 2003, 270, S209–S212. [Google Scholar] [CrossRef]
- Tsuchida, T.; Koga, R.; Fukatsu, T. Host plant specialization governed by facultative symbiont. Science 2004, 303, 1989. [Google Scholar] [CrossRef] [PubMed]
- Wagner, S.M.; Martinez, A.J.; Ruan, Y.M.; Kim, K.L.; Lenhart, P.A.; Dehnel, A.C.; Oliver, K.M.; White, J.A. Facultative endosymbionts mediate dietary breadth in a polyphagous herbivore. Funct. Ecol. 2015, 29, 1402–1410. [Google Scholar] [CrossRef]
- Tsuchida, T.; Koga, R.; Horikawa, M.; Tsunoda, T.; Maoka, T.; Matsumoto, S.; Simon, J.C.; Fukatsu, T. Symbiotic bacterium modifies aphid body color. Science 2010, 330, 1102–1104. [Google Scholar] [CrossRef] [PubMed]
- Nikoh, N.; Tsuchida, T.; Maeda, T.; Yamaguchi, K.; Shigenobu, S.; Koga, R.; Fukatsu, T. Genomic insight into symbiosis-induced insect color change by a facultative bacterial endosymbiont, Candidatus Rickettsiella viridis. mBio 2018, 9, e00890-18. [Google Scholar] [CrossRef]
- Pérez-Brocal, V.; Gil, R.; Ramos, S.; Lamelas, A.; Postigo, M.; Michelena, J.M.; Silva, F.J.; Moya, A.; Latorre, A. A small microbial genome: The end of a long symbiotic relationship? Science 2006, 314, 312–313. [Google Scholar] [CrossRef]
- Lamelas, A.; Gosalbes, M.J.; Manzano-Marín, A.; Pereto, J.; Moya, A.; Latorre, A. Serratia symbiotica from the aphid Cinara cedri: A missing link from facultative to obligate insect endosymbiont. PLoS Genet. 2011, 7, e1002357. [Google Scholar] [CrossRef]
- Manzano-Marín, A.; Latorre, A. Settling down: The genome of Serratia symbiotica from the aphid Cinara tujafilina zooms in on the process of accommodation to a cooperative intracellular life. Genome Biol. Evol. 2014, 6, 1683–1698. [Google Scholar] [CrossRef]
- Manzano-Marín, A.; Simon, J.C.; Latorre, A. Reinventing the wheel and making it round again: Evolutionary convergence in Buchnera-Serratia symbiotic consortia between the distantly related Lachninae aphids Tuberolachnus salignus and Cinara cedri. Genome Biol. Evol. 2016, 8, 1440–1458. [Google Scholar] [CrossRef]
- Manzano-Marín, A.; Szabo, G.; Simon, J.C.; Horn, M.; Latorre, A. Happens in the best of subfamilies: Establishment and repeated replacements of co-obligate secondary endosymbionts within Lachninae aphids. Environ. Microbiol. 2017, 19, 393–408. [Google Scholar] [CrossRef]
- Meseguer, A.S.; Manzano-Marín, A.; Coeur d’acier, A.; Clamens, A.L.; Godefroid, M.; Jousselin, E. Buchnera has changed flatmate but the repeated replacement of co-obligate symbionts is not associated with the ecological expansions of their aphid hosts. Mol. Ecol. 2017, 26, 2363–2378. [Google Scholar] [CrossRef]
- Manzano-Marín, A.; Coeur d’acier, A.; Clamens, A.L.; Orvain, C.; Cruaud, C.; Barbe, V.; Jousselin, E. A freeloader? The highly eroded yet large genome of the Serratia symbiotica symbiont of Cinara strobi. Genome Biol. Evol. 2018, 10, 2178–2189. [Google Scholar] [CrossRef] [PubMed]
- De Clerck, C.; Fujiwara, A.; Joncour, P.; Leonard, S.; Felix, M.L.; Francis, F.; Jijakli, M.H.; Tsuchida, T.; Massart, S. A metagenomic approach from aphid’s hemolymph sheds light on the potential roles of co-existing endosymbionts. Microbiome 2015, 3, 63. [Google Scholar] [CrossRef] [PubMed]
- Manzano-Marín, A. No evidence for Wolbachia as a nutritional co-obligate endosymbiont in the aphid Pentalonia nigronervosa. bioRxiv 2019, 609511. [Google Scholar] [CrossRef]
- Tsuchida, T.; Koga, R.; Shibao, H.; Matsumoto, T.; Fukatsu, T. Diversity and geographic distribution of secondary endosymbiotic bacteria in natural populations of the pea aphid, Acyrthosiphon pisum. Mol. Ecol. 2002, 11, 2123–2135. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, J.; West, J.A.; Via, S.; Godfray, H.C. Population genetic structure and secondary symbionts in host-associated populations of the pea aphid complex. Evolution 2012, 66, 375–390. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, J.P.; Outreman, Y.; Mieuzet, L.; Simon, J.C. Bacterial communities associated with host-adapted populations of pea aphids revealed by deep sequencing of 16S ribosomal DNA. PLoS ONE 2015, 10, e0120664. [Google Scholar] [CrossRef]
- Brady, C.M.; White, J.A. Cowpea aphid (Aphis craccivora) associated with different host plants has different facultative endosymbionts. Ecol. Entomol. 2013, 38, 433–437. [Google Scholar] [CrossRef]
- Brady, C.M.; Asplen, M.K.; Desneux, N.; Heimpel, G.E.; Hopper, K.R.; Linnen, C.R.; Oliver, K.M.; Wulff, J.A.; White, J.A. Worldwide populations of the aphid Aphis craccivora are infected with diverse facultative bacterial symbionts. Microb. Ecol. 2014, 67, 195–204. [Google Scholar] [CrossRef]
- Xu, S.F.; Jiang, L.Y.; Qiao, G.X.; Chen, J. The bacterial flora associated with the polyphagous aphid Aphis gossypii Glover (Hemiptera: Aphididae) is strongly affected by host plants. Microb. Ecol. 2019. [Google Scholar] [CrossRef]
- Medina, R.F.; Nachappa, P.; Tamborindeguy, C. Differences in bacterial diversity of host-associated populations of Phylloxera notabilis Pergande (Hemiptera: Phylloxeridae) in pecan and water hickory. J. Evol. Biol. 2011, 24, 761–771. [Google Scholar] [CrossRef]
- Najar-Rodriguez, A.J.; McGraw, E.A.; Mensah, R.K.; Pittman, G.W.; Walter, G.H. The microbial flora of Aphis gossypii: Patterns across host plants and geographical space. J. Invertebr. Pathol. 2009, 100, 123–126. [Google Scholar] [CrossRef]
- Jones, R.T.; Bressan, A.; Greenwell, A.M.; Fierer, N. Bacterial communities of two parthenogenetic aphid species cocolonizing two host plants across the Hawaiian islands. Appl. Environ. Microbiol. 2011, 77, 8345–8349. [Google Scholar] [CrossRef]
- Russell, J.A.; Weldon, S.; Smith, A.H.; Kim, K.L.; Hu, Y.; Lukasik, P.; Doll, S.; Anastopoulos, I.; Novin, M.; Oliver, K.M. Uncovering symbiont-driven genetic diversity across North American pea aphids. Mol. Ecol. 2013, 22, 2045–2059. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, S.; Luo, J.Y.; Wang, C.Y.; Lv, L.M.; Cui, J.J. Bacterial communities of the cotton aphid Aphis gossypii associated with Bt cotton in northern China. Sci. Rep. 2016, 6, 22958. [Google Scholar] [CrossRef] [PubMed]
- Gallo-Franco, J.J.; Duque-Gamboa, D.N.; Toro-Perea, N. Bacterial communities of Aphis gossypii and Myzus persicae (Hemiptera: Aphididae) from pepper crops (Capsicum sp.). Sci. Rep. 2019, 9, 5766. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Liu, X.; Poncelet, N.; He, K.; Francis, F.; Wang, Z. Detection and geographic distribution of seven facultative endosymbionts in two Rhopalosiphum aphid species. Microbiologyopen 2019, 8, e00817. [Google Scholar] [CrossRef] [PubMed]
- Zytynska, S.E.; Weisser, W.W. The natural occurrence of secondary bacterial symbionts in aphids. Ecol. Entomol. 2016, 41, 13–26. [Google Scholar] [CrossRef]
- Zytynska, S.E.; Meyer, S.T.; Sturm, S.; Ullmann, W.; Mehrparvar, M.; Weisser, W.W. Secondary bacterial symbiont community in aphids responds to plant diversity. Oecologia 2016, 180, 735–747. [Google Scholar] [CrossRef]
- Fakhour, S.; Ambroise, J.; Renoz, F.; Foray, V.; Gala, J.L.; Hance, T. A large-scale field study of bacterial communities in cereal aphid populations across Morocco. FEMS Microbiol. Ecol. 2018, 94, fiy003. [Google Scholar] [CrossRef]
- Henry, L.M.; Maiden, M.C.; Ferrari, J.; Godfray, H.C. Insect life history and the evolution of bacterial mutualism. Ecol. Lett. 2015, 18, 516–525. [Google Scholar] [CrossRef]
- McLean, A.H.C.; Godfray, H.C.J.; Ellers, J.; Henry, L.M. Host relatedness influences the composition of aphid microbiomes. Environ. Microbiol. Rep. 2019, 11, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Remaudière, G.; Remaudière, M. Catalogue of the World’s Aphididae: Homoptera Aphidoidea; Institut National de la Recherche Agronomique: Paris, Franch, 1997. [Google Scholar]
- Smith, C.F. Keys to and descriptions of the genera of Pemphigini in North America (Homoptera: Aphididae: Pemphiginae). North Carolina Agri. Exp. Station Tech. Bull. 1974, 226, 1–61. [Google Scholar]
- Moran, N.A. The evolution of host-plant alternation in aphids: Evidence for specialization as a dead end. Am. Nat. 1988, 132, 681–706. [Google Scholar] [CrossRef]
- Moran, N.A.; Whitham, T.G. Evolutionary reduction of complex life cycles: Loss of host alternation in Pemphigus (Homoptera: Aphididae). Evolution 1988, 42, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.K. Fauna of India and the Adjacent Countries: Homoptera: Aphidoidea. Part. 3, Subfamilies: Pemphiginae; Zoological Survey of India: Calcutta, India, 1984. [Google Scholar]
- Smith, R.G. Wax glands, wax production and the functional significance of wax use in three aphid species (Homoptera: Aphididae). J. Nat. Hist. 1999, 33, 513–530. [Google Scholar] [CrossRef]
- Zhang, G.X.; Qiao, G.X.; Zhong, T.S.; Zhang, W.Y. Fauna Sinica Insecta, Vol. 14. Homoptera: Mindaridae and Pemphigidae; Science Press: Beijing, China, 1999. [Google Scholar]
- Pike, N.; Foster, W. Fortress repair in the social aphid species Pemphigus spyrothecae. Anim. Behav. 2004, 67, 909–914. [Google Scholar] [CrossRef]
- Pike, N.; Whitfield, J.A.; Foster, W.A. Ecological correlates of sociality in Pemphigus aphids, with a partial phylogeny of the genus. BMC Evol. Biol. 2007, 7, 185. [Google Scholar] [CrossRef]
- Chong, R.A.; Moran, N.A. Evolutionary loss and replacement of Buchnera, the obligate endosymbiont of aphids. ISME J. 2018, 12, 898–908. [Google Scholar] [CrossRef]
- Burke, G.R.; Normark, B.B.; Favret, C.; Moran, N.A. Evolution and diversity of facultative symbionts from the aphid subfamily Lachninae. Appl. Environ. Microbiol. 2009, 75, 5328–5335. [Google Scholar] [CrossRef]
- Jousselin, E.; Coeur d’Acier, A.; Vanlerberghe-Masutti, F.; Duron, O. Evolution and diversity of Arsenophonus endosymbionts in aphids. Mol. Ecol. 2013, 22, 260–270. [Google Scholar] [CrossRef]
- Wang, Z.; Su, X.M.; Wen, J.; Jiang, L.Y.; Qiao, G.X. Widespread infection and diverse infection patterns of Wolbachia in Chinese aphids. Insect Sci. 2014, 21, 313–325. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Magoc, T.; Salzberg, S.L. Flash: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glockner, F.O. The SILVA and “All-Species Living Tree Project (LTP)” taxonomic frameworks. Nucleic Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef]
- Navas-Molina, J.A.; Peralta-Sanchez, J.M.; Gonzalez, A.; McMurdie, P.J.; Vazquez-Baeza, Y.; Xu, Z.; Ursell, L.K.; Lauber, C.; Zhou, H.; Song, S.J.; et al. Advancing our understanding of the human microbiome using QIIME. Methods Enzymol. 2013, 531, 371–444. [Google Scholar] [CrossRef]
- R Core Team. R: A language and environment for statistical computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 15 January 2020).
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Oksanen, F.J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.; O’Hara, R.B.; Simpson, G.; Solymos, P.; et al. Vegan: Community ecology package. R Package Version 2.5-4. 2018. Available online: https://CRAN.R-project.org/package=vegan (accessed on 15 January 2020).
- Paradis, E.; Claude, J.; Strimmer, K. Ape: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate-a practical and powerful approach to multiple testing. J. R. Statist. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Harrell, F.E., Jr. With contributions from Charles Dupont and many others. Hmisc: Harrell Misc. R Package Version 4.2-0. 2019. Available online: https://CRAN.R-project.org/package=Hmisc (accessed on 15 January 2020).
- Tamas, I.; Klasson, L.; Canback, B.; Naslund, A.K.; Eriksson, A.S.; Wernegreen, J.J.; Sandstrom, J.P.; Moran, N.A.; Andersson, S.G. 50 million years of genomic stasis in endosymbiotic bacteria. Science 2002, 296, 2376–2379. [Google Scholar] [CrossRef] [PubMed]
- Oliver, K.M.; Smith, A.H.; Russell, J.A. Defensive symbiosis in the real world-advancing ecological studies of heritable, protective bacteria in aphids and beyond. Funct. Ecol. 2014, 28, 341–355. [Google Scholar] [CrossRef]
- Vorburger, C.; Rouchet, R. Are aphid parasitoids locally adapted to the prevalence of defensive symbionts in their hosts? BMC Evol. Biol. 2016, 16, 271. [Google Scholar] [CrossRef]
- Xu, T.T.; Chen, J.; Jiang, L.Y.; Qiao, G.X. Diversity of bacteria associated with Hormaphidinae aphids (Hemiptera: Aphididae). Insect Sci. 2019. [Google Scholar] [CrossRef]
- Smith, C.F. Pemphiginae associated with the roots of conifers in north America (Homoptera: Aphididae). Ann. Entomol. Soc. Am. 1969, 62, 1128–1152. [Google Scholar] [CrossRef]
- Stroyan, H.L.G. The life cycle and generic position of Aphis tremulae L., 1761 (Aphidoidea: Pemphiginae), with a description of the viviparous morphs and a discussion of spruce root aphids in the British Isles. Biol. J. Linn. Soc. 1975, 7, 45–72. [Google Scholar] [CrossRef]
- Danielsson, R. The root-feeding generations of gall-making aphids of the genera Pachypappa Koch, Pachypappella Baker, and Gootiella Tullgren (Homoptera. Aphidoidea. Pemphigidae). Acta Phytopathol. Entomol. Hung. 1990, 25, 77–88. [Google Scholar]
- Zhang, H.C.; Qiao, G.X. Systematic status of the genus Formosaphis Takahashi and the evolution of galls based on the molecular phylogeny of Pemphigini (Hemiptera: Aphididae: Eriosomatinae). Syst. Entomol. 2007, 32, 690–699. [Google Scholar] [CrossRef]
- Pike, K.S.; Graf, G.; Foottit, R.G.; Maw, H.E.L.; von Dohlen, C.; Harpel, J.; Pantoja, A.; Emmert, S.; Hagerty, A.M. Eriosomatine aphids (Hemiptera: Aphididae: Eriosomatinae) associated with moss and roots of conifer and willow in forests of the Pacific Northwest of North America. Can. Entomol. 2012, 144, 555–576. [Google Scholar] [CrossRef][Green Version]
- Cornell, H.V. The secondary chemistry and complex morphology of galls formed by the Cynipinae (Hymenoptera): Why and how? Am. Midl. Nat. 1983, 110, 225–234. [Google Scholar] [CrossRef]
- Price, P.W.; Waring, G.L.; Fernandes, G.W. Hypotheses on the adaptive nature of galls. Proc. Entomol. Soc. Wash. 1986, 88, 361–363. [Google Scholar]
- Stone, G.N.; Schönrogge, K. The adaptive significance of insect gall morphology. Trends Ecol. Evol. 2003, 18, 512–522. [Google Scholar] [CrossRef]
- Miller, D.G.; Ivey, C.T.; Shedd, J.D. Support for the microenvironment hypothesis for adaptive value of gall induction in the California gall wasp, Andricus quercuscalifornicus. Entomol. Exp. Appl. 2009, 132, 126–133. [Google Scholar] [CrossRef]
- Vorburger, C.; Gouskov, A. Only helpful when required: A longevity cost of harbouring defensive symbionts. J. Evol. Biol. 2011, 24, 1611–1617. [Google Scholar] [CrossRef]
- Polin, S.; Simon, J.C.; Outreman, Y. An ecological cost associated with protective symbionts of aphids. Ecol. Evol. 2014, 4, 826–830. [Google Scholar] [CrossRef]
- Zytynska, S.E.; Thighiouart, K.; Frago, E. A meta-analysis on the benefits and costs of hosting secondary endosymbionts in sap-sucking insects. bioRxiv 2019, 563031. [Google Scholar] [CrossRef]
- Ferrari, J.; Scarborough, C.L.; Godfray, H.C. Genetic variation in the effect of a facultative symbiont on host-plant use by pea aphids. Oecologia 2007, 153, 323–329. [Google Scholar] [CrossRef]
- McLean, A.H.; van Asch, M.; Ferrari, J.; Godfray, H.C. Effects of bacterial secondary symbionts on host plant use in pea aphids. Proc. R. Soc. B. 2011, 278, 760–766. [Google Scholar] [CrossRef]
- Henry, L.M.; Peccoud, J.; Simon, J.C.; Hadfield, J.D.; Maiden, M.J.; Ferrari, J.; Godfray, H.C. Horizontally transmitted symbionts and host colonization of ecological niches. Curr. Biol. 2013, 23, 1713–1717. [Google Scholar] [CrossRef]
- Smith, A.H.; Lukasik, P.; O’Connor, M.P.; Lee, A.; Mayo, G.; Drott, M.T.; Doll, S.; Tuttle, R.; Disciullo, R.A.; Messina, A.; et al. Patterns, causes and consequences of defensive microbiome dynamics across multiple scales. Mol. Ecol. 2015, 24, 1135–1149. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Luo, J.; Wang, L.; Zhang, L.; Zhu, X.; Jiang, W.; Cui, J. Bacterial communities in natural versus pesticide-treated Aphis gossypii populations in north China. Microbiologyopen 2019, 8, e00652. [Google Scholar] [CrossRef] [PubMed]
- Oliver, K.M.; Moran, N.A.; Hunter, M.S. Costs and benefits of a superinfection of facultative symbionts in aphids. Proc. R. Soc. B. 2006, 273, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Guay, J.F.; Boudreault, S.; Michaud, D.; Cloutier, C. Impact of environmental stress on aphid clonal resistance to parasitoids: Role of Hamiltonella defensa bacterial symbiosis in association with a new facultative symbiont of the pea aphid. J. Insect Physiol. 2009, 55, 919–926. [Google Scholar] [CrossRef] [PubMed]
- Ayoubi, A.; Talebi, A.A.; Fathipour, Y.; Mehrabadi, M. Coinfection of the secondary symbionts, Hamiltonella defensa and Arsenophonus sp. contribute to the performance of the major aphid pest, Aphis gossypii (Hemiptera: Aphididae). Insect Sci. 2020, 27, 86–98. [Google Scholar] [CrossRef]
- Leclair, M.; Polin, S.; Jousseaume, T.; Simon, J.C.; Sugio, A.; Morliere, S.; Fukatsu, T.; Tsuchida, T.; Outreman, Y. Consequences of coinfection with protective symbionts on the host phenotype and symbiont titres in the pea aphid system. Insect Sci. 2017, 24, 798–808. [Google Scholar] [CrossRef]
- McLean, A.H.C.; Parker, B.J.; Hrcek, J.; Kavanagh, J.C.; Wellham, P.A.D.; Godfray, H.C.J. Consequences of symbiont co-infections for insect host phenotypes. J. Anim. Ecol. 2018, 87, 478–488. [Google Scholar] [CrossRef]
- Moran, N.A.; Dunbar, H.E. Sexual acquisition of beneficial symbionts in aphids. Proc. Natl. Acad. Sci. USA 2006, 103, 12803–12806. [Google Scholar] [CrossRef]
- Gehrer, L.; Vorburger, C. Parasitoids as vectors of facultative bacterial endosymbionts in aphids. Biol. Lett. 2012, 8, 613–615. [Google Scholar] [CrossRef]
- Chrostek, E.; Pelz-Stelinski, K.; Hurst, G.D.D.; Hughes, G.L. Horizontal transmission of intracellular insect symbionts via plants. Front. Microbiol. 2017, 8, 2237. [Google Scholar] [CrossRef]
- Pons, I.; Renoz, F.; Noel, C.; Hance, T. Circulation of the cultivable symbiont Serratia symbiotica in aphids is mediated by plants. Front. Microbiol. 2019, 10, 764. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Host Plant | Sample Number | Sample Name |
|---|---|---|
| Species across Eriosomatinae | ||
| Anacardiaceae | 3 | Chaetogeoica sp., Kaburagia rhusicola, Schlechtendalia chinensis |
| Magnoliaceae | 1 | Formosaphis micheliae |
| Oleaceae | 2 | Ligustrum lucidum, Prociphilus sp. |
| Rosaceae | 3 | Eriosoma lanigerum, Prociphilus kuwanai, Prociphilus pini |
| Salicaceae | 15 | Epipemphigus imaicus, Epipemphigus niisimae, Epipemphigus yunanensis, Pemphigus borealis, Pemphigus bursarius, Pemphigus matsumurai, Pemphigus populitransversus, Pemphigus sinobursarius, Pemphigus sp. 1, Pemphigus sp. 2, Pemphigus tibetensis, Thecabius beijingensis, Thecabius sp. 1, Thecabius sp. 2, Thecabius sp. 3 |
| Ulmaceae | 9 | Eriosoma sp. 1, Eriosoma sp. 2, Eriosoma sp. 3, Eriosoma sp. 4, Tetraneura sp. 1, Tetraneura sp. 2, Tetraneura sp. 3, Tetraneura sp. 4, Tetraneura sp. 5 |
| Species from Gansu Province | ||
| Salicaceae | 4 | Pemphigus sinobursarius, Thecabius sp. 1, Thecabius sp. 2, Thecabius sp. 3 |
| Ulmaceae | 5 | Tetraneura sp. 3, Tetraneura sp. 4, Tetraneura sp. 5, Eriosoma sp. 2, Eriosoma sp. 3 |
| (a) | ||||
| Bacterial Community | Facultative Symbiont Community | |||
| Host plant | All 4 groups | 2 groups | All 4 groups | 2 groups |
| 0.25, 0.010 | 0.42, 0.008 | 0.27, 0.002 | 0.57, 0.013 | |
| (b) | ||||
| Bacterial Community | Facultative Symbiont Community | |||
| All species | 15 species | All species | 15 species | |
| Geographic distribution | −0.01, 0.50 | 0.14, 0.19 | 0.03, 0.36 | 0.14, 0.20 |
| Aphid relatedness | 0.07, 0.16 | −0.19, 0.96 | 0.01, 0.41 | −0.19, 0.97 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, T.-T.; Jiang, L.-Y.; Chen, J.; Qiao, G.-X. Host Plants Influence the Symbiont Diversity of Eriosomatinae (Hemiptera: Aphididae). Insects 2020, 11, 217. https://doi.org/10.3390/insects11040217
Xu T-T, Jiang L-Y, Chen J, Qiao G-X. Host Plants Influence the Symbiont Diversity of Eriosomatinae (Hemiptera: Aphididae). Insects. 2020; 11(4):217. https://doi.org/10.3390/insects11040217
Chicago/Turabian StyleXu, Ting-Ting, Li-Yun Jiang, Jing Chen, and Ge-Xia Qiao. 2020. "Host Plants Influence the Symbiont Diversity of Eriosomatinae (Hemiptera: Aphididae)" Insects 11, no. 4: 217. https://doi.org/10.3390/insects11040217
APA StyleXu, T.-T., Jiang, L.-Y., Chen, J., & Qiao, G.-X. (2020). Host Plants Influence the Symbiont Diversity of Eriosomatinae (Hemiptera: Aphididae). Insects, 11(4), 217. https://doi.org/10.3390/insects11040217