Temperature Tolerance and Thermal Environment of European Seed Bugs

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Critical Thermal Minimum (CTmin)
2.3. Critical Thermal Maximum (CTmax)
2.4. Thermal Tolerance Breadth (TTB), Heat Tolerance (TH), Cold Tolerance (TC)
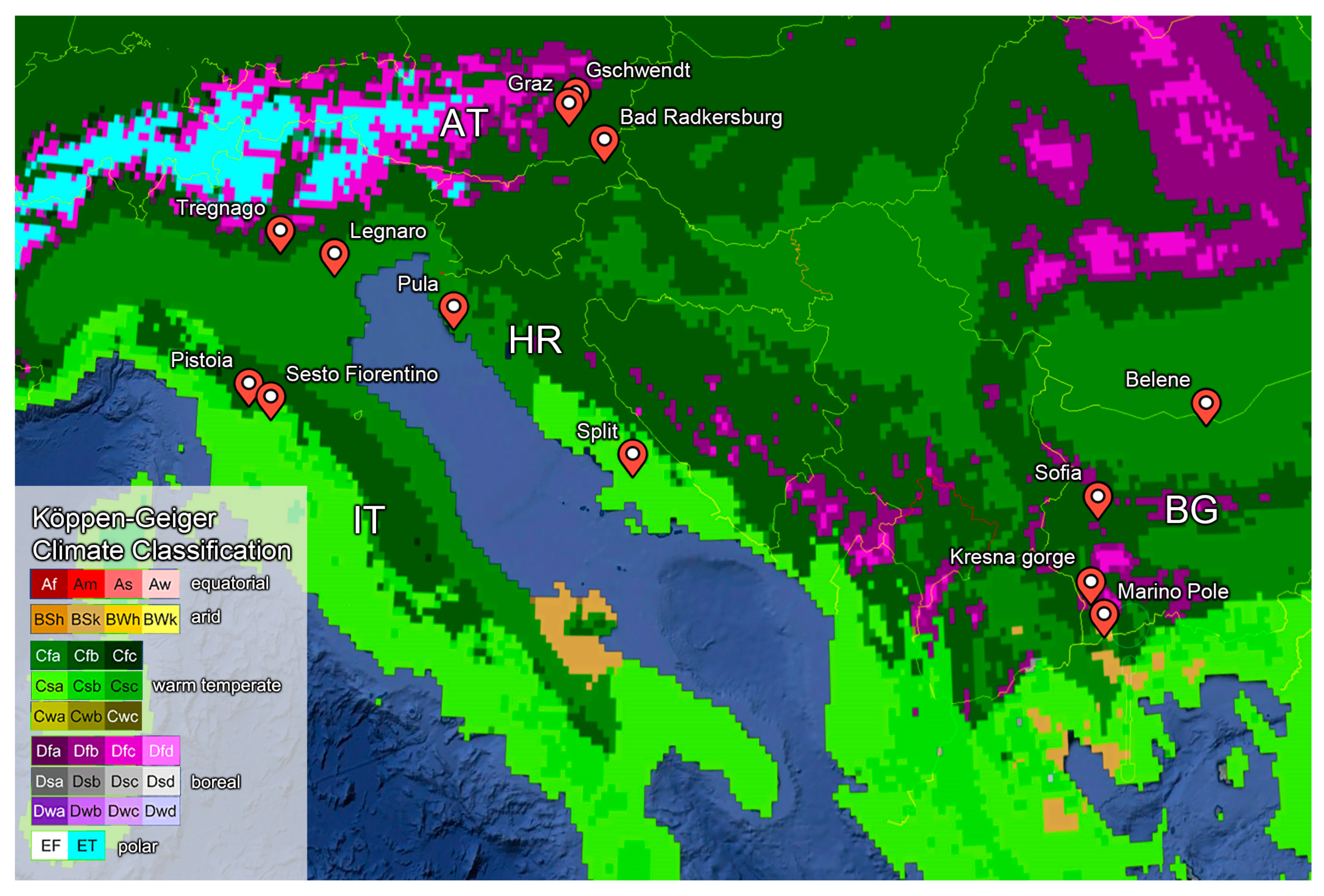
2.5. Climate Data, Bioclimatic Variables, and Data Analysis
2.6. Testing for a Phylogenetic Signal
3. Results
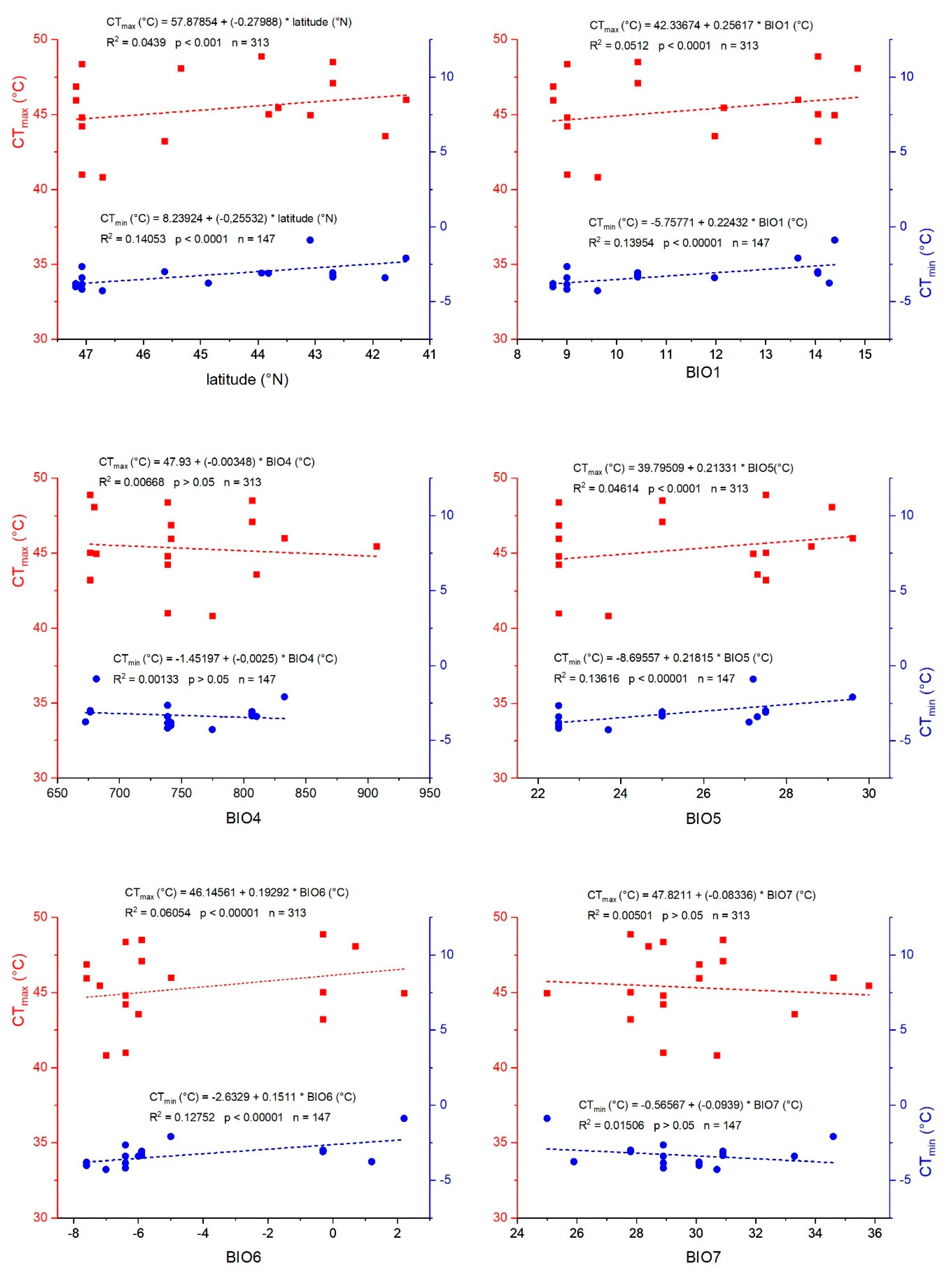
3.1. Critical Thermal Limits (CTmin, CTmax)
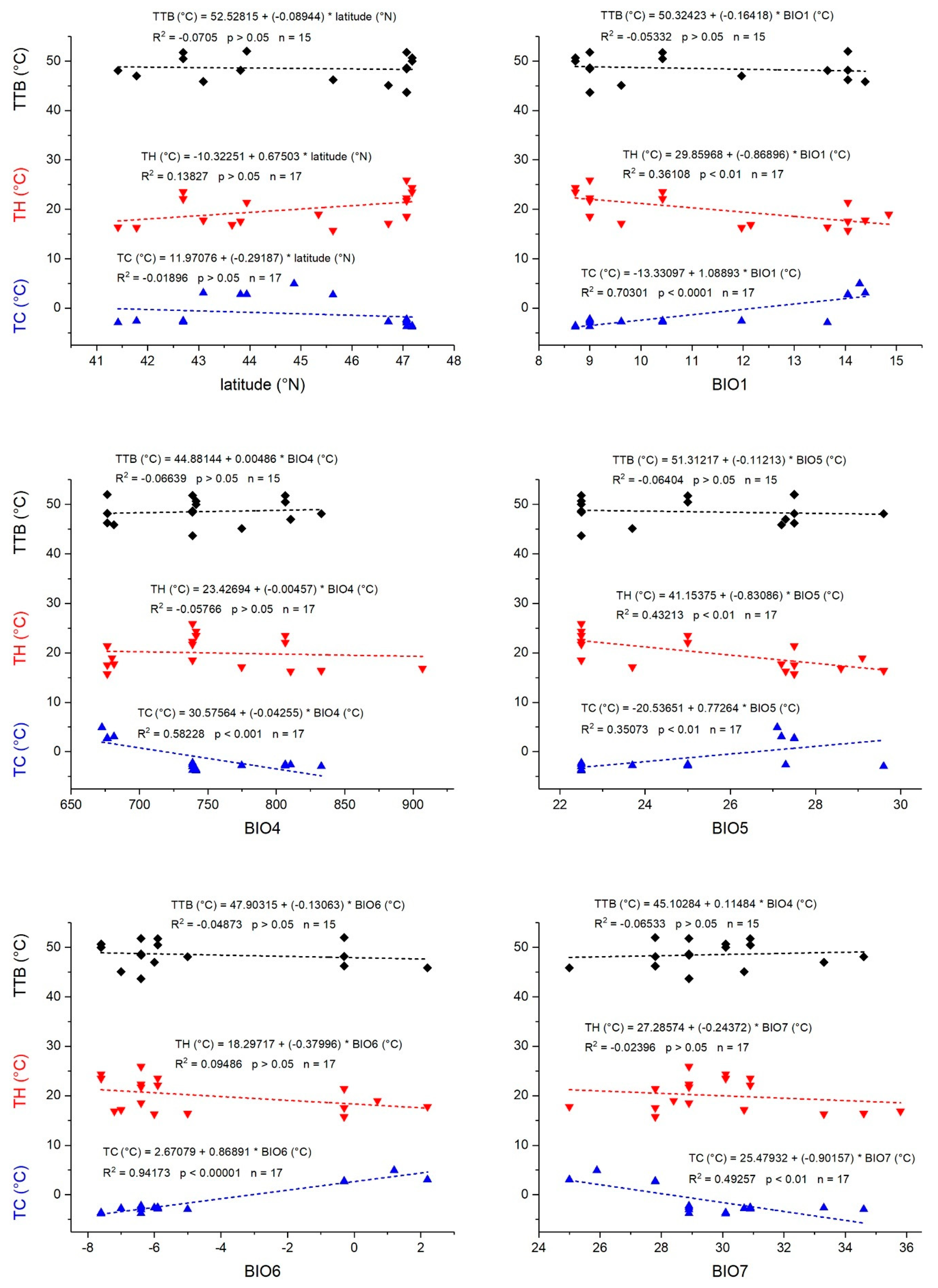
3.2. Correlation with Derived Physiological Thermal Tolerance Parameters (TTB, TC, TH)
3.3. Testing for Phylogenetic Signal
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| KCC Köppen–Geiger climate classification (climate, precipitation, temperature) | |
| Bsk | arid, summer dry, cold arid |
| Cfa | warm temperate, fully humid, hot summer |
| Cfb | warm temperate, fully humid, warm summer |
| Csa | warm temperate, summer dry, hot summer |
| Bioclimatic variables (mean values 1970–2000) | |
| BIO1 | annual mean temperature |
| BIO2 | mean diurnal range (mean of monthly (max temp − min temp)) |
| BIO3 | isothermality (BIO2/BIO7 × 100) |
| BIO4 | temperature seasonality (standard deviation × 100) |
| BIO5 | max temperature of warmest month |
| BIO6 | min temperature of coldest month |
| BIO7 | temperature annual range (BIO5 − BIO6) |
| BIO12 | annual precipitation |
| Physiological parameters | |
| CTmin | critical thermal minimum |
| CTmax | critical thermal maximum |
| TTB | thermal tolerance breadth (CTmax − CTmin) |
| TC | cold tolerance (BIO6 − CTmin) |
| TH | heat tolerance (CTmax − BIO5) |
References
- Henry, T.J. Biodiversity of Heteroptera. In Insect Biodiversity; Foottit, R.G., Adler, P.H., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 279–335. ISBN 9781118945568. [Google Scholar]
- Weber, N.A. A survey of the insects and related arthropods of arctic Alaska. Part I. Trans. Am. Entomol. Soc. (1890-) 1950, 76, 147–206. [Google Scholar]
- Makarova, O.L.; Makarov, K.V. True bugs (Heteroptera) from the Arctic Dolgii Island, the Barents sea. Entomol. Rev. 2006, 86, 423–432. [Google Scholar] [CrossRef]
- Schuh, R.T.; Weirauch, C. True Bugs of the World (Hemiptera, Heteroptera). Classification and Natural History, 2nd ed.; Siri Scientific Press: Manchester, UK, 2020; ISBN 978-0-9957496-9-6. [Google Scholar]
- Morrison, R. Invasive Insects: The Top 4 “Most Wanted” List. Available online: https://entomologytoday.org/2018/06/21/invasive-insects-the-top-4-most-wanted-list/ (accessed on 26 August 2019).
- Aukema, B. Fauna Europaea: Hemiptera, Heteroptera, Pentatomorpha, Lygaeoidea: Fauna Europaea Version 2017.06. Available online: https://fauna-eu.org (accessed on 29 November 2019).
- Rabitsch, W.; Adlbauer, K. Erstnachweis und bekannte Verbreitung von Oxycarenus lavaterae (Fabricius, 1787) in Österreich (Heteroptera: Lygaeidae). Beitr. Entomofaunist. 2001, 2, 49–54. [Google Scholar]
- Hoffmann, H.-J. Oxycarenus lavaterae (Fabricius, 1787) nun auch im Norden Frankreichs, und im SW Deutschlands. In Heteropteron: Mitteilungsblatt der Arbeitsgruppe Mitteleuropäischer Heteropterologen; Hoffmann, H.C., Ed.; Arbeitsgruppe Mitteleuropäischer Heteropterologen, Hrsg.: Köln, Germany, 2005; pp. 25–26. [Google Scholar]
- Kalushkov, P.; Simov, N.; Tzankova, R. Biology and Acclimatization of Oxycarenus lavaterae (Heteroptera: Lygaeidae) A new invasive Mediterranean Species in Bulgarian Fauna. In Proceedings of the International Conference “Alien Arthropods in South East Europe—Crossroad of Three Continents”, Sofia, Bulgaria, 19–21 September 2007; University of Forestry: Sofia, Bulgaria, 2007; pp. 44–47. [Google Scholar]
- Schneider, A.; Dorow, W.H. Erstnachweis von Oxycarenus lavaterae (Fabricius, 1787) für Hessen. In Heteropteron: Mitteilungsblatt der Arbeitsgruppe Mitteleuropäischer Heteropterologen; Hoffmann, H.C., Ed.; Arbeitsgruppe Mitteleuropäischer Heteropterologen, Hrsg.: Köln, Germany, 2016; pp. 23–24. [Google Scholar]
- Musolin, D.L.; Fujisaki, K. Changes in ranges: Trends in distribution of true bugs (Heteroptera) under conditions of the current climate warning. Russ. Entomol. J. 2006, 15, 175–179. [Google Scholar]
- Taylor, F. Ecology and evolution of physiological time in insects. Am. Nat. 1981, 117, 1–23. [Google Scholar] [CrossRef]
- Chown, S.L.; Nicolson, S.W. Insect Physiological Ecology: Mechanisms and Patterns; Oxford University Press: New York, NY, USA, 2004; ISBN 978-0198515494. [Google Scholar]
- Dixon, A.F.G.; Honěk, A.; Keil, P.; Kotela, M.A.A.; Šizling, A.L.; Jarošík, V. Relationship between the minimum and maximum temperature thresholds for development in insects. Funct. Ecol. 2009, 23, 257–264. [Google Scholar] [CrossRef]
- Bale, J.S. Insects at low temperature: A predictable relationship? Funct. Ecol. 1991, 5, 291. [Google Scholar] [CrossRef]
- Stevens, G.C. The latitudinal gradient in geographical range: How so many species coexist in the tropics. Am. Nat. 1989, 133, 240–256. [Google Scholar] [CrossRef]
- Chown, S.L. Physiological variation in insects: Hierarchical levels and implications. J. Insect Physiol. 2001, 47, 649–660. [Google Scholar] [CrossRef]
- Sunday, J.M.; Bates, A.E.; Dulvy, N.K. Global analysis of thermal tolerance and latitude in ectotherms. Proc. Biol. Sci. 2011, 278, 1823–1830. [Google Scholar] [CrossRef]
- Gaston, K.J.; Chown, S.L. Elevation and climatic colerance: A test using dung beetles. Oikos 1999, 86, 584. [Google Scholar] [CrossRef]
- Addo-Bediako, A.; Chown, S.L.; Gaston, K.J. Thermal tolerance, climatic variability and latitude. Proc. Biol. Sci. 2000, 267, 739–745. [Google Scholar] [CrossRef]
- Sheldon, K.S.; Tewksbury, J.J. The impact of seasonality in temperature on thermal tolerance and elevational range size. Ecology 2014, 95, 2134–2143. [Google Scholar] [CrossRef] [PubMed]
- Sunday, J.; Bennett, J.M.; Calosi, P.; Clusella-Trullas, S.; Gravel, S.; Hargreaves, A.L.; Leiva, F.P.; Verberk, W.C.E.P.; Olalla-Tárraga, M.Á.; Morales-Castilla, I. Thermal tolerance patterns across latitude and elevation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20190036. [Google Scholar] [CrossRef] [PubMed]
- Oyen, K.J.; Giri, S.; Dillon, M.E. Altitudinal variation in bumble bee (Bombus) critical thermal limits. J. Therm. Biol. 2016, 59, 52–57. [Google Scholar] [CrossRef]
- Sunday, J.M.; Bates, A.E.; Kearney, M.R.; Colwell, R.K.; Dulvy, N.K.; Longino, J.T.; Huey, R.B. Thermal-safety margins and the necessity of thermoregulatory behavior across latitude and elevation. Proc. Natl. Acad. Sci. USA 2014, 111, 5610–5615. [Google Scholar] [CrossRef]
- Winwood-Smith, H.S.; Alton, L.A.; Franklin, C.E.; White, C.R. Does greater thermal plasticity facilitate range expansion of an invasive terrestrial anuran into higher latitudes? Conserv. Physiol. 2015, 3, cov010. [Google Scholar] [CrossRef]
- Ducatez, S.; Baguette, M.; Trochet, A.; Chaput-Bardy, A.; Legrand, D.; Stevens, V.; Fréville, H. Flight endurance and heating rate vary with both latitude and habitat connectivity in a butterfly species. Oikos 2013, 122, 601–611. [Google Scholar] [CrossRef]
- Alford, L.; Blackburn, T.M.; Bale, J.S. Effect of latitude and acclimation on the lethal temperatures of the peach-potato aphid Myzus persicae. Agric. Forest Entomol. 2012, 14, 69–79. [Google Scholar] [CrossRef]
- Buckley, L.B.; Huey, R.B. Temperature extremes: Geographic patterns, recent changes, and implications for organismal vulnerabilities. Glob. Chang. Biol. 2016, 22, 3829–3842. [Google Scholar] [CrossRef]
- Potter, K.A.; Arthur Woods, H.; Pincebourde, S. Microclimatic challenges in global change biology. Glob. Chang. Biol. 2013, 19, 2932–2939. [Google Scholar] [CrossRef]
- Deutsch, C.A.; Tewksbury, J.J.; Huey, R.B.; Sheldon, K.S.; Ghalambor, C.K.; Haak, D.C.; Martin, P.R. Impacts of climate warming on terrestrial ectotherms across latitude. Proc. Natl. Acad. Sci. USA 2008, 105, 6668–6672. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A. Physiological climatic limits in Drosophila: Patterns and implications. J. Exp. Biol. 2010, 213, 870–880. [Google Scholar] [CrossRef] [PubMed]
- Pearson, O.P.; Bradford, D.F. Thermoregulation of lizards and toads at high altitudes in Peru. Copeia 1976, 1976, 155–170. [Google Scholar] [CrossRef]
- Pincebourde, S.; Casas, J. Narrow safety margin in the phyllosphere during thermal extremes. Proc. Natl. Acad. Sci. USA 2019, 116, 5588–5596. [Google Scholar] [CrossRef]
- Hadley, N.F. Water relations of the desert scorpion, Hadrurus arizonensis. J. Exp. Biol. 1970, 53, 547–558. [Google Scholar]
- Hadley, N.F.; Quinlan, M.C.; Kennedy, M.L. Evaporative cooling in the desert cicada. J. Exp. Biol. 1991, 159, 269–283. [Google Scholar]
- Buckley, L.B.; Miller, E.F.; Kingsolver, J.G. Ectotherm thermal stress and specialization across altitude and latitude. Integr. Comp. Biol. 2013, 53, 571–581. [Google Scholar] [CrossRef]
- Kearney, M.R.; Shine, R.; Porter, W.P. The potential for behavioral thermoregulation to buffer “cold-blooded” animals against climate warming. Proc. Natl. Acad. Sci. USA 2009, 106, 3835–3840. [Google Scholar] [CrossRef]
- Woods, H.A.; Dillon, M.E.; Pincebourde, S. The roles of microclimatic diversity and of behavior in mediating the responses of ectotherms to climate change. J. Therm. Biol. 2015, 54, 86–97. [Google Scholar] [CrossRef]
- Vorhees, A.S.; Gray, E.M.; Bradley, T.J. Thermal resistance and performance correlate with climate in populations of a widespread mosquito. Physiol. Biochem. Zool. 2013, 86, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Hazell, S.P.; Pedersen, B.P.; Worland, M.R.; Blackburn, T.M.; Bale, J.S. A method for the rapid measurement of thermal tolerance traits in studies of small insects. Physiol. Entomol. 2008, 33, 389–394. [Google Scholar] [CrossRef]
- Klok, C.J.; Chown, S.L. Critical thermal limits, temperature tolerance and water balance of a sub-Antarctic caterpillar, Pringleophaga marioni (Lepidoptera: Tineidae). J. Insect Physiol. 1997, 43, 685–694. [Google Scholar] [CrossRef]
- Lutterschmidt, W.I.; Hutchison, V.H. The critical thermal maximum: Data to support the onset of spasms as the definitive end point. Can. J. Zool. 1997, 75, 1553–1560. [Google Scholar] [CrossRef]
- Aukema, B.; Rieger, C. Catalogue of the Heteroptera of the Palaearctic Region: Pentatomomorpha I; Netherlands Entomological Society: Amsterdam, The Netherlands, 2001; Volume 4, ISBN 90-71912-21-3. [Google Scholar]
- Andersen, J.L.; Manenti, T.; Sørensen, J.G.; MacMillan, H.A.; Loeschcke, V.; Overgaard, J. How to assess Drosophila cold tolerance: Chill coma temperature and lower lethal temperature are the best predictors of cold distribution limits. Funct. Ecol. 2015, 29, 55–65. [Google Scholar] [CrossRef]
- Chown, S.L.; Jumbam, K.R.; Sørensen, J.G.; Terblanche, J.S. Phenotypic variance, plasticity and heritability estimates of critical thermal limits depend on methodological context. Funct. Ecol. 2009, 23, 133–140. [Google Scholar] [CrossRef]
- Terblanche, J.S.; Deere, J.A.; Clusella-Trullas, S.; Janion, C.; Chown, S.L. Critical thermal limits depend on methodological context. Proc. R. Soc. Lond. B 2007, 274, 2935–2942. [Google Scholar] [CrossRef] [PubMed]
- Kovac, H.; Käfer, H.; Stabentheiner, A. The respiratory metabolism of Polistes biglumis, a paper wasp from mountainous regions. Insects 2020, 11, 165. [Google Scholar] [CrossRef]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Freckleton, R.P. The seven deadly sins of comparative analysis. J. Evol. Biol. 2009, 22, 1367–1375. [Google Scholar] [CrossRef] [PubMed]
- Gordon, E.R.L.; McFrederick, Q.; Weirauch, C. Phylogenetic evidence for ancient and persistent environmental symbiont reacquisition in Largidae (Hemiptera: Heteroptera). Appl. Environ. Microbiol. 2016, 82, 7123–7133. [Google Scholar] [CrossRef] [PubMed]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis, Version 3.6; Available online: http://www.mesquiteproject.org (accessed on 24 September 2019).
- Erregger, B.; Kovac, H.; Stabentheiner, A.; Hartbauer, M.; Römer, H.; Schmidt, A.K.D. Cranking up the heat: Relationships between energetically costly song features and the increase in thorax temperature in male crickets and katydids. J. Exp. Biol. 2017, 220, 2635–2644. [Google Scholar] [CrossRef]
- Pagel, M. Inferring the historical patterns of biological evolution. Nature 1999, 401, 877–884. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing. Available online: http://www.R-project.org/ (accessed on 24 September 2019).
- Orme, D.; Freckleton, R.P.; Thomas, G.H.; Petzoldt, T.; Fritz, S.A.; Isaac, N. CAPER: Comparative analyses of phylogenetics and evolution in R. Methods Ecol. Evol. 2013, 3, 145–151. [Google Scholar]
- Kellermann, V.; Overgaard, J.; Hoffmann, A.A.; Flojgaard, C.; Svenning, J.-C.; Loeschcke, V. Upper thermal limits of Drosophila are linked to species distributions and strongly constrained phylogenetically. Proc. Natl. Acad. Sci. USA 2012, 109, 16228–16233. [Google Scholar] [CrossRef]
- Gallego, B.; Verdú, J.R.; Carrascal, L.M.; Lobo, J.M. A protocol for analysing thermal stress in insects using infrared thermography. J. Therm. Biol. 2016. [Google Scholar] [CrossRef]
- Nyamukondiwa, C.; Chidawanyika, F.; Machekano, H.; Mutamiswa, R.; Sands, B.; Mgidiswa, N.; Wall, R. Climate variability differentially impacts thermal fitness traits in three coprophagic beetle species. PLoS ONE 2018, 13, e0198610. [Google Scholar] [CrossRef]
- Baudier, K.M.; Mudd, A.E.; Erickson, S.C.; O’Donnell, S. Microhabitat and body size effects on heat tolerance: Implications for responses to climate change (army ants: Formicidae, Ecitoninae). J. Anim. Ecol. 2015, 84, 1322–1330. [Google Scholar] [CrossRef]
- Ribeiro, P.L.; Camacho, A.; Navas, C.A. Considerations for assessing maximum critical temperatures in small ectothermic animals: Insights from leaf-cutting Ants. PLoS ONE 2012, 7, e32083. [Google Scholar] [CrossRef]
- Verble-Pearson, R.M.; Gifford, M.E.; Yanoviak, S.P. Variation in thermal tolerance of North American ants. J. Therm. Biol. 2015, 48, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Hemmings, Z.; Andrew, N.R. Effects of microclimate and species identity on body temperature and thermal tolerance of ants (Hymenoptera: Formicidae). Austral Entomol. 2017, 56, 104–114. [Google Scholar] [CrossRef]
- Janowiecki, M.; Clifton, E.; Avalos, A.; Vargo, E.L. Upper thermal tolerance of tropical and temperate termite species (Isoptera: Rhinotermitidae, Termitidae): A test of the climate variability hypothesis in termites. Insectes Soc. 2019, 16, 1206. [Google Scholar] [CrossRef]
- Hu, X.P.; Appel, A.G. Seasonal variation of critical thermal limits and temperature tolerance in formosan and eastern subterranean termites (Isoptera: Rhinotermitidae). Environ. Entomol. 2004, 33, 197–205. [Google Scholar] [CrossRef]
- Raschmanová, N.; Šustr, V.; Kováč, Ľ.; Parimuchová, A.; Devetter, M. Testing the climatic variability hypothesis in edaphic and subterranean Collembola (Hexapoda). J. Therm. Biol. 2018, 78, 391–400. [Google Scholar] [CrossRef]
- Chown, S.L.; Addo-Bediako, A.; Gaston, K.J. Physiological variation in insects: Large-scale patterns and their implications. Comp. Biochem. Physiol. B 2002, 131, 587–602. [Google Scholar] [CrossRef]
- Rozsypal, J.; Koštál, V. Supercooling and freezing as eco-physiological alternatives rather than mutually exclusive strategies: A case study in Pyrrhocoris apterus. J. Insect Physiol. 2018, 111, 53–62. [Google Scholar] [CrossRef]
- Mumladze, L.; Asanidze, Z.; Walther, F.; Hausdorf, B. Beyond elevation: Testing the climatic variability hypothesis vs. Rapoport’s rule in vascular plant and snail species in the Caucasus. Biol. J. Linn. Soc. 2017, 121, 753–763. [Google Scholar] [CrossRef]
- Koštál, V.; Šimek, P. Overwintering strategy in Pyrrhocoris apterus (Heteroptera): The relations between life-cycle, chill tolerance and physiological adjustments. J. Insect Physiol. 2000, 46, 1321–1329. [Google Scholar] [CrossRef]
- Honek, A.; Martinkova, Z. Behavioural thermoregulation hastens spring mating activity in Pyrrhocoris apterus (Heteroptera: Pyrrhocoridae). J. Therm. Biol. 2019, 84, 185–189. [Google Scholar] [CrossRef]
- Baudier, K.M.; D’Amelio, C.L.; Malhotra, R.; O’Connor, M.P.; O’Donnell, S. Extreme insolation: Climatic variation shapes the evolution of thermal tolerance at multiple Scales. Am. Nat. 2018, 192, 347–359. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Chown, S.L.; Clusella-Trullas, S.; Fox, C. Upper thermal limits in terrestrial ectotherms: How constrained are they? Funct. Ecol. 2013, 27, 934–949. [Google Scholar] [CrossRef]
- Hansen, T.F.; Pienaar, J.; Orzack, S.H. A comparative method for studying adaptation to a randomly evolving environment. Evolution 2008, 62, 1965–1977. [Google Scholar] [CrossRef]
- Losos, J.B. Phylogenetic niche conservatism, phylogenetic signal and the relationship between phylogenetic relatedness and ecological similarity among species. Ecol. Lett. 2008, 11, 995–1003. [Google Scholar] [CrossRef]
- Teets, N.M.; Hahn, D.A. Genetic variation in the shape of cold-survival curves in a single fly population suggests potential for selection from climate variability. J. Evol. Biol. 2018, 31, 543–555. [Google Scholar] [CrossRef]
- Ayrinhac, A.; Debat, V.; Gibert, P.; Kister, A.-G.; Legout, H.; Moreteau, B.; Vergilino, R.; David, J.R. Cold adaptation in geographical populations of Drosophila melanogaster: Phenotypic plasticity is more important than genetic variability. Funct. Ecol. 2004, 18, 700–706. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Shirriffs, J.; Scott, M. Relative importance of plastic vs genetic factors in adaptive differentiation: Geographical variation for stress resistance in Drosophila melanogaster from eastern Australia. Funct. Ecol. 2005, 19, 222–227. [Google Scholar] [CrossRef]
- Terblanche, J.S.; Klok, C.J.; Krafsur, E.S.; Chown, S.L. Phenotypic Plasticity and Geographic Variation in Thermal Tolerance and Water Loss of the Tsetse Glossina pallidipes (Diptera: Glossinidae): Implications for Distribution Modelling. Am. J. Trop. Med. Hyg. 2006, 74, 786–794. [Google Scholar] [CrossRef][Green Version]
- Hamblin, A.L.; Youngsteadt, E.; López-Uribe, M.M.; Frank, S.D. Physiological thermal limits predict differential responses of bees to urban heat-island effects. Biol. Lett. 2017, 13. [Google Scholar] [CrossRef]
- Rabitsch, W. The times they are a-changin’: Driving forces of recent additions to the Heteroptera fauna of Austria. In Advances in Heteroptera Research. Festschrift in Honor of 80th Anniversary of Michail Josifov; Grozeva, S., Simov, N., Eds.; Pensoft Publishers: Sofia, Moscow, 2008; pp. 309–326. ISBN 9789546423115. [Google Scholar]
- Musolin, D.L.; Saulich, A.K. Responses of insects to the current climate changes: From physiology and behavior to range shifts. Entmol. Rev. 2012, 92, 715–740. [Google Scholar] [CrossRef]
- Musolin, D.L. Insects in a warmer world: Ecological, physiological and life-history responses of true bugs (Heteroptera) to climate change. Glob. Chang. Biol. 2007, 13, 1565–1585. [Google Scholar] [CrossRef]
- Musolin, D.L.; Fujisaki, K.; Numata, H. Photoperiodic control of diapause termination, colour change and postdiapause reproduction in the southern green stink bug, Nezara viridula. Physiol. Entomol. 2007, 32, 64–72. [Google Scholar] [CrossRef]
- Saulich, A.K.; Musolin, D.L. Diapause in the seasonal cycle of stink bugs (Heteroptera, Pentatomidae) from the Temperate Zone. Entmol. Rev. 2012, 92, 1–26. [Google Scholar] [CrossRef]
- Kiritani, K. Factors affecting the winter mortality in the southern green stink bug, Nezara viridula L. Ann. Entomol. Soc. Fr. 1966, 2, 199–207. [Google Scholar]
- Kiritani, K. The impact of global warming and land-use change on the pest status of rice and fruit bugs (Heteroptera) in Japan. Glob. Chang. Biol. 2007, 13, 1586–1595. [Google Scholar] [CrossRef]
- Kiritani, K.; Hókyo, N.; Kimura, K. Differential Winter Mortality Relative to Sex in the Population of the Southern Green Stink Bug, Nezara viridula (Pentatomidae, Hemiptera). Jpn. J. Appl. Entomol. Zool. 1962, 6, 242–246. [Google Scholar] [CrossRef]
- Yukawa, J.; Kiritani, K.; Gyoutoku, N.; Uechi, N.; Yamaguchi, D.; Kamitani, S. Distribution range shift of two allied species, Nezara viridula and N. antennata (Hemiptera: Pentatomidae), in Japan, possibly due to global warming. Appl. Entomol. Zool. 2007, 42, 205–215. [Google Scholar] [CrossRef][Green Version]
- Yukawa, J.; Kiritani, K.; Kawasawa, T.; Higashiura, Y.; Sawamura, N.; Nakada, K.; Gyotoku, N.; Tanaka, A.; Kamitani, S.; Matsuo, K.; et al. Northward range expansion by Nezara viridula (Hemiptera: Pentatomidae) in Shikoku and Chugoku Districts, Japan, possibly due to global warming. Appl. Entomol. Zool. 2009, 44, 429–437. [Google Scholar] [CrossRef]
- Kiritani, K. Different effects of climate change on the population dynamics of insects. Appl. Entomol. Zool. 2013, 48, 97–104. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Subfamily | Family | Suborder | Order |
|---|---|---|---|---|
| Arocatus longiceps | Lygaeinae | Lygaeidae | Heteroptera | Hemiptera |
| Melanocoryphus albomaculatus | ||||
| Orsillus depressus | Orsillinae | |||
| Orsillus maculatus | ||||
| Oxycarenus lavaterae | Oxycareninae | |||
| Rhyparochromus vulgaris | Rhyparochrominae | |||
| Scolopostethus pictus | ||||
| Pyrrhocoris apterus | Pyrrhocoridae |
| Species | Location - KCC | Lat (°N)/Long (°E) | Mass | CTmin ± SD | Mass | CTmax ± SD | TTB | TH | TC | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| (mg) | (°C) | n | (mg) | (°C) | n | (°C) | (°C) | (°C) | |||
| Arocatus longiceps | Graz (AT) - Cfb | 47.071/15.44 | 6.6 ± 1.1 | −4.2 ± 0.4 | 8 | 6.7 ± 1.3 | 44.2 ± 0.5 | 30 | 48.4 | 21.7 | −2.2 |
| Sofia (BG) - Cfa | 42.696/23.334 | 6.8 ± 0.9 | −3.1 ± 0.8 | 6 | - | - | - | - | - | −2.8 | |
| Kresna gorge (BG) - Cfa | 41.783/23.155 | 3.6 ± 0.6 | −3.4 ± 1.6 | 2 | 3.6 ± 1 | 43.6 ± 0.6 | 25 | 47.0 | 16.3 | −2.6 | |
| Melanocoryphus albomaculatus | Pistoia (IT) - Cfa | 43.939/10.849 | 14.4 ± 3.4 | −3.1 ± 0.3 | 9 | 14.6 ± 2.6 | 48.9 ± 0.5 | 14 | 52.0 | 21.4 | 2.8 |
| Orsillus depressus | Graz (AT) - Cfb | 47.071/15.44 | 9.7 ± 1.8 | −3.8 ± 0.4 | 9 | 9.2 ± 1.6 | 44.8 ± 0.8 | 10 | 48.6 | 22.3 | −2.6 |
| Orsillus maculatus | Tregnago (IT) - Cfb | 45.629/11.095 | 15.2 ± 4.6 | −3.0 ± 0.7 | 5 | 10.7 ± 2.4 | 43.2 ± 1.2 | 25 | 46.2 | 15.7 | 2.7 |
| Marino Pole (BG) - Bsk | 41.419/23.331 | 11.5 ± 2.5 | −2.1 ± 2.6 | 7 | 11.4 ± 2.7 | 46.0 ± 0.4 | 10 | 48.1 | 16.4 | −2.9 | |
| Oxycarenus lavaterae | Sofia (BG) - Cfa | 42.696/23.334 | 2.5 ± 0.8 | −3.2 ± 2.0 | 8 | 3.1 ± 0.5 | 48.5 ± 0.3 | 19 | 51.7 | 23.5 | −2.7 |
| Graz (AT) - Cfb | 47.071/15.44 | 4.0 ± 0.7 | −3.4 ± 2.1 | 8 | 3.4 ± 0.8 | 48.4 ± 0.2 | 19 | 51.8 | 25.9 | −3.0 | |
| Legnaro (IT) - Cfa | 45.346/11.964 | - | - | - | 4.7 ± 0.7 | 48.1 ± 0.4 | 32 | - | 19.0 | - | |
| Rhyparochromus vulgaris | Gschwendt (AT) - Cfb | 47.179/15.573 | 15.1 ± 2.5 | −3.8 ± 0.5 | 10 | 17.1 ± 3.5 | 46.9 ± 0.4 | 10 | 50.7 | 24.4 | −3.8 |
| Scolopostethus pictus | Radkersburg (AT) - Cfb | 46.714/15.998 | 2.8 ± 0.7 | −4.3 ± 0.2 | 9 | 3.5 ± 0.6 | 40.8 ± 0.6 | 29 | 45.1 | 17.1 | −2.7 |
| Graz (AT) – Cfb | 47.071/15.44 | 3.9 ± 0.8 | −2.7 ± 0.4 | 9 | 3.3 ± 0.6 | 41 ± 0.4.0 | 9 | 43.7 | 18.5 | −3.7 | |
| Pyrrhocoris apterus | Graz (AT) - Cfb | 47.179/15.573 | 55.9 ± 9.3 | −4.0 ± 0.6 | 21 | 53.2 ± 13.4 | 46.0 ± 0.5 | 10 | 50.0 | 23.5 | −3.6 |
| SestoFno/FI (IT) - Csa | 43.818/11.204 | 24.5 ± 2.8 | −3.1 ± 1.4 | 8 | 39.0 ± 8.5 | 45.0 ± 0.5 | 19 | 48.1 | 17.5 | 2.8 | |
| Belene (BG) - Cfa | 43.652/25.129 | - | - | - | 25.4 ± 5 | 45.5 ± 0.6 | 32 | - | 16.9 | - | |
| Sofia (BG) - Cfa | 42.696/23.334 | 29.5 ± 6.7 | −3.4 ± 1.6 | 9 | 47.3 ± 8.4 | 47.1 ± 0.2 | 10 | 50.4 | 22.1 | −2.5 | |
| Split (HR) - Csa | 43.09/16.752 | 39.0 ± 6.4 | −0.9 ± 0.6 | 10 | 29.8 ± 2.9 | 45.0 ± 0.4 | 10 | 45.9 | 17.8 | 3.1 | |
| Pula (HR) - Cfa | 44.867/13.85 | 31.6 ± 3.4 | −3.8 ± 0.7 | 9 | - | - | - | - | - | 5.0 | |
| CTmin | CTmax | |||||
|---|---|---|---|---|---|---|
| df | F-Quotient | p-Value | df | F-Quotient | p-Value | |
| KCC | 3 | 10.46 | <0.0001 | 4 | 46.7 | <0.0001 |
| Species | 7 | 0.8 | 0.5903 | 7 | 508.36 | <0.0001 |
| Mass | 1 | 1.08 | 0.3002 | 1 | 26.43 | <0.0001 |
| GLM for all bioclimatic parameters | 7 | 10.37 | <0.0001 | 7 | 36.01 | <0.0001 |
| BIO1 = annual mean temperature | 1 | 6.43 | 0.0123 | 1 | 48.27 | <0.0001 |
| BIO2 = mean diurnal range | 1 | 15.23 | <0.001 | 1 | 40.57 | <0.0001 |
| BIO3 = isothermality ((BIO2/BIO7) × 100) | 1 | 17.60 | <0.0001 | 1 | 48.69 | <0.0001 |
| BIO4 = temperature seasonality (SD × 100) | 1 | 0.76 | 0.3863 | 1 | 9.32 | <0.005 |
| BIO5 = max temperature of warmest month | 1 | 1.14 | 0.2875 | 1 | 50.04 | <0.0001 |
| BIO6 = min temperature of coldest month | 1 | 20.43 | <0.0001 | 1 | 16.32 | <0.001 |
| BIO12 = annual precipitation | 1 | 2.50 | 0.1163 | 1 | 46.84 | <0.0001 |
| TTB | TC | TH | |||||||
|---|---|---|---|---|---|---|---|---|---|
| df | F-Quotient | p-Value | df | F-Quotient | p-Value | df | F-Quotient | p-Value | |
| KCC | 3 | 0.86 | 0.6225 | 3 | 1.22 | 0.3939 | 3 | 2.07 | 0.2223 |
| Species | 7 | 11.37 | 0.0357 | 7 | 1.09 | 0.4788 | 7 | 3.87 | 0.0776 |
| Mass | 1 | 2.24 | 0.2316 | 1 | 0.7 | 0.4398 | 1 | 1.36 | 0.2959 |
| GLM | 4 | 1.47 | 0.2836 | 4 | 224.56 | <0.0001 | 7 | 1.65 | 0.2379 |
| BIO1 | 1 | 5.42 | 0.0421 | 1 | 23.97 | <0.0005 | - | - | - |
| BIO2 | - | - | - | 1 | 26.00 | <0.0005 | - | - | - |
| BIO3 | - | - | - | 1 | 28.40 | <0.0005 | - | - | - |
| BIO4 | 1 | 5.21 | 0.0456 | - | - | - | - | - | - |
| BIO5 | 1 | 5.44 | 0.0419 | - | - | - | - | - | - |
| BIO6 | 1 | 5.00 | 0.0494 | 1 | 18.93 | <0.001 | - | - | - |
| BIO12 | - | - | - | - | - | - | - | - | - |
| BIO7 | 1 | 0.14 | 0.7135 | - | - | - | - | - | - |
| Pagel’s λ | p(λ=0) | p(λ=1) | |
|---|---|---|---|
| Body mass (mg) | 1 | <0.001 | 1 |
| CTmax (°C) | 1 | 0.235 | 1 |
| CTmin (°C) | 0 | 1 | 0.008 |
| TTB (°C) | 0 | 1 | 0.197 |
| TH (°C) | 0 | 1 | 0.011 |
| TC (°C) | 0 | 1 | 0.069 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Käfer, H.; Kovac, H.; Simov, N.; Battisti, A.; Erregger, B.; Schmidt, A.K.D.; Stabentheiner, A. Temperature Tolerance and Thermal Environment of European Seed Bugs. Insects 2020, 11, 197. https://doi.org/10.3390/insects11030197
Käfer H, Kovac H, Simov N, Battisti A, Erregger B, Schmidt AKD, Stabentheiner A. Temperature Tolerance and Thermal Environment of European Seed Bugs. Insects. 2020; 11(3):197. https://doi.org/10.3390/insects11030197
Chicago/Turabian StyleKäfer, Helmut, Helmut Kovac, Nikolay Simov, Andrea Battisti, Bettina Erregger, Arne K. D. Schmidt, and Anton Stabentheiner. 2020. "Temperature Tolerance and Thermal Environment of European Seed Bugs" Insects 11, no. 3: 197. https://doi.org/10.3390/insects11030197
APA StyleKäfer, H., Kovac, H., Simov, N., Battisti, A., Erregger, B., Schmidt, A. K. D., & Stabentheiner, A. (2020). Temperature Tolerance and Thermal Environment of European Seed Bugs. Insects, 11(3), 197. https://doi.org/10.3390/insects11030197