Marking Drosophila suzukii (Diptera: Drosophilidae) with Fluorescent Dusts

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Pigment Application Methods
2.1.1. Effect on Survival
2.1.2. Marking Efficiency and Persistence
2.2. Effects of Colour on Survival
2.3. Effects on Behaviour
2.3.1. Olfaction of Marked Flies in Laboratory Trap Assays
2.3.2. Orientation of Marked Flies by Trap Recapture in the Field
2.4. Persistence of Marks during Trapping or Storage
2.5. Statistical Analysis
3. Results
3.1. Pigment Application Methods
3.1.1. Effect on Survival
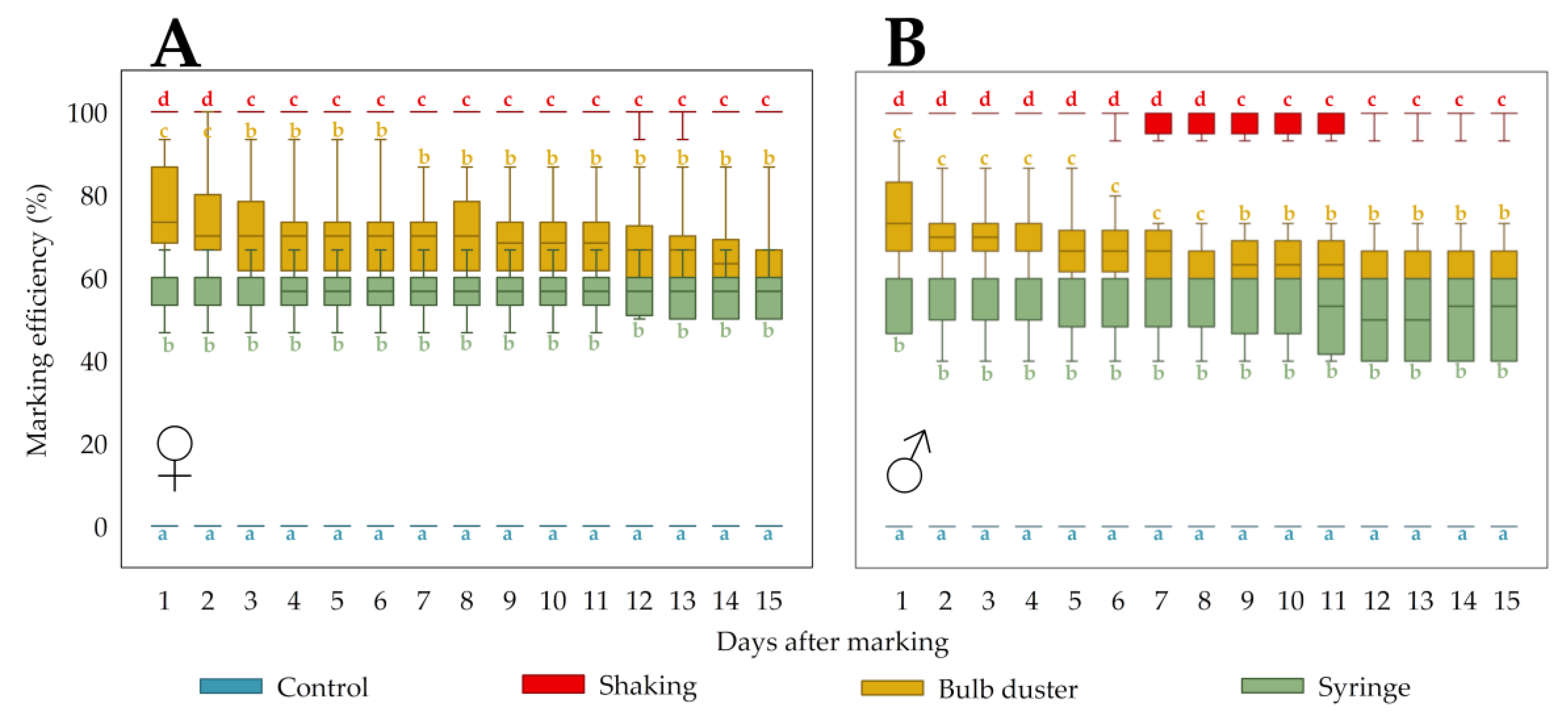
3.1.2. Marking Efficiency and Persistence
3.2. Effects of Colour (Dye) on Survival
3.3. Effects on Behaviour
3.3.1. Olfaction of Marked Flies in Laboratory Trap Assays
3.3.2. Orientation of Marked Flies by Trap Recapture in the Field
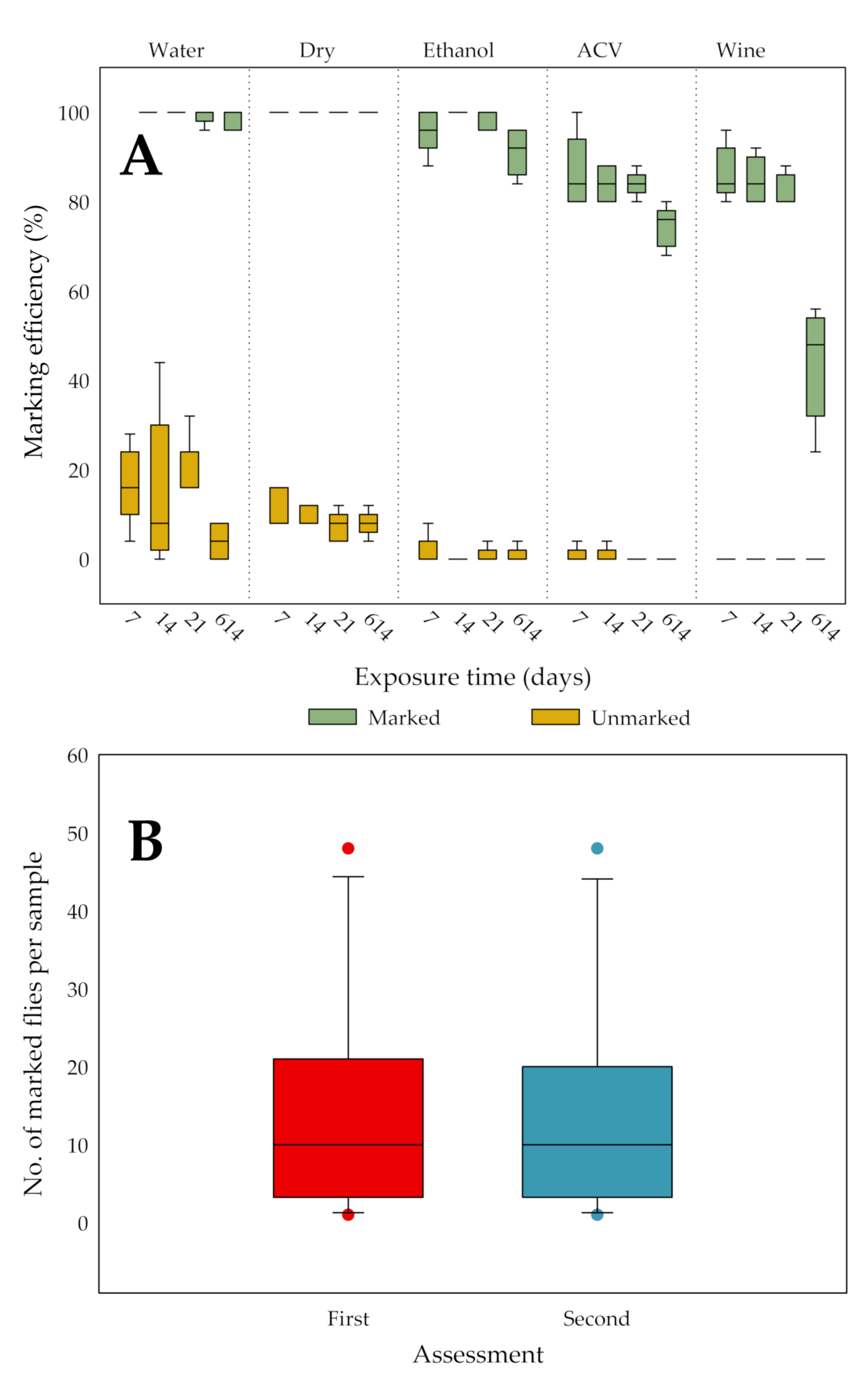
3.4. Persistence of Marks during Trapping or Storage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zetek, J. Determining the flight of mosquitos. Ann. Entomol. Soc. Am. 1913, 6, 5–21. [Google Scholar] [CrossRef]
- Geiger, J.C.; Purdy, W.C.; Tarbett, R.E. Effective malaria control in a ricefield district, with observations on experimental mosquito flights. J. Am. Med. Assoc. 1919, 72, 844–847. [Google Scholar] [CrossRef]
- Dudley, J.E.; Searles, E.M. Color marking of the striped cucumber beetle (Diabrotica vittata Fab.) and preliminary experiments to determine its flight. J. Econ. Entomol. 1923, 16, 363–368. [Google Scholar] [CrossRef]
- Hagler, J.R.; Jackson, C.G. Methods for marking insects: Current techniques and future prospects. Annu. Rev. Entomol. 2001, 46, 511–543. [Google Scholar] [CrossRef] [PubMed]
- Hamby, K.A.; Bolda, M.P.; Sheehan, M.E.; Zalom, F.G. Seasonal monitoring for Drosophila suzukii (Diptera: Drosophilidae) in California commercial raspberries. Environ. Entomol. 2014, 43, 1008–1018. [Google Scholar] [CrossRef]
- Ørsted, I.V.; Ørsted, M. Species distribution models of the Spotted Wing Drosophila (Drosophila suzukii, Diptera: Drosophilidae) in its native and invasive range reveal an ecological niche shift. J. Appl. Ecol. 2019, 56, 423–435. [Google Scholar] [CrossRef]
- Walsh, D.B.; Bolda, M.P.; Goodhue, R.E.; Dreves, A.J.; Lee, J.; Bruck, D.J.; Walton, V.M.; O’Neal, S.D.; Zalom, F.G. Drosophila suzukii (Diptera: Drosophilidae): Invasive pest of ripening soft fruit expanding its geographic range and damage potential. J. Integr. Pest Manag. 2011, 2, G1–G7. [Google Scholar] [CrossRef]
- Andreazza, F.; Bernardi, D.; Baronio, C.A.; Pasinato, J.; Nava, D.E.; Botton, M. Toxicities and effects of insecticidal toxic baits to control Drosophila suzukii and Zaprionus indianus (Diptera: Drosophilidae). Pest Manag. Sci. 2017, 73, 146–152. [Google Scholar] [CrossRef]
- Belien, T.; Thys, T.; Fassotte, C.; Walrant, C.; Tomme, M.; Bolen, M.; Bylemans, D. Population dynamics of Drosophila suzukii (Diptera: Drosophilidae) in Belgium in 2013. Commun. Agric. Appl. Biol. Sci. 2014, 79, 169–175. [Google Scholar]
- Dalton, D.T.; Walton, V.M.; Shearer, P.W.; Walsh, D.B.; Caprile, J.; Isaacs, R. Laboratory survival of Drosophila suzukii under simulated winter conditions of the Pacific Northwest and seasonal field trapping in five primary regions of small and stone fruit production in the United States. Pest Manag. Sci. 2011, 67, 1368–1374. [Google Scholar] [CrossRef]
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Lee, J.C.; Bruck, D.J.; Dreves, A.J.; Ioriatti, C.; Vogt, H.; Baufeld, P. In focus: Spotted wing Drosophila, Drosophila suzukii, across perspectives. Pest Manag. Sci. 2011, 67, 1349–1351. [Google Scholar] [CrossRef]
- Mazzi, D.; Bravin, E.; Meraner, M.; Finger, R.; Kuske, S. Economic impact of the introduction and establishment of Drosophila suzukii on sweet cherry production in Switzerland. Insects 2017, 8, 18. [Google Scholar] [CrossRef] [PubMed]
- Mortelmans, J.; Casteels, H.; Beliën, T. Drosophila suzukii (Diptera: Drosophilidae): A pest species new to Belgium. Belgian J. Zool. 2012, 142, 143–146. [Google Scholar]
- Jones, V.P.; Hagler, J.R.; Brunner, J.F.; Baker, C.C.; Wilburn, T.D. An inexpensive immunomarking technique for studying movement patterns of naturally occurring insect populations. Environ. Entomol. 2006, 35, 827–836. [Google Scholar] [CrossRef]
- Klick, J.; Lee, J.C.; Hagler, J.R.; Bruck, D.J.; Yang, W.Q. Evaluating Drosophila suzukii immunomarking for mark-capture research. Entomol. Exp. Appl. 2014, 152, 31–41. [Google Scholar] [CrossRef]
- Jones, V.P.; Melton, T.D.; Baker, C.C. Optimizing immunomarking systems and development of a new marking system based on wheat. J. Insect Sci. 2011, 11, 1–16. [Google Scholar] [CrossRef][Green Version]
- Klick, J.; Yang, W.Q.; Bruck, D.J. Marking Drosophila suzukii (Diptera: Drosophilidae) with Rubidium or 15N. J. Econ. Entomol. 2015, 108, 1447–1451. [Google Scholar] [CrossRef]
- Verhulst, N.O.; Loonen, J.A.C.M.; Takken, W. Advances in methods for colour marking of mosquitoes. Parasit. Vectors 2013, 6, 1–7. [Google Scholar] [CrossRef]
- Dickens, B.L.; Brant, H.L. Effects of marking methods and fluorescent dusts on Aedes aegypti survival. Parasit. Vectors 2014, 7, 1–9. [Google Scholar] [CrossRef]
- Rojas-Araya, D.; Alto, B.W.; Burkett-Cadena, N.; Cummings, D.A. Detection of fluorescent powders and their effect on survival and recapture of Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2020, 57, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Van Keymeulen, M.; Hertveldt, L.; Pelerents, C. Methods for improving both the quantitative and qualitative aspects of rearing Delia brassicae for sterile release programmes. Entomol. Exp. Appl. 1981, 30, 231–240. [Google Scholar] [CrossRef]
- Weldon, C. Marking Queensland fruit fly, Bactrocera tryoni (Froggatt) (Diptera: Tephritidae) with fluorescent pigments: Pupal emergence, adult mortality, and visibility and persistence of marks. Gen. Appl. Entomol. 2005, 34, 7–13. [Google Scholar]
- Dominiak, B.C.; Sundaralingam, S.; Jiang, L.; Jessup, A.J.; Barchia, I.M. Impact of marker dye on adult eclosion and flight ability of mass produced Queensland fruit fly Bactrocera tryoni (Froggatt) (Diptera: Tephritidae). Aust. J. Entomol. 2010, 49, 166–169. [Google Scholar] [CrossRef]
- Makumbe, L.D.M.; Manrakhan, A.; Weldon, C.W. Marking Bactrocera dorsalis (Diptera: Tephritidae) with fluorescent pigments: Effects of pigment colour and concentration. African Entomol. 2017, 25, 220–234. [Google Scholar] [CrossRef]
- Christie, R.M. Colour Chemistry., 2nd ed.; The Royal Society of Chemistry: Cambridge, UK, 2015; ISBN 978-1-84973-328-1. [Google Scholar]
- Newbacher, C.; Jatla, A. Fluorescence. In Coloring of Plastics: Fundamentals; Charvat, R.A., Ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2004; pp. 242–257. ISBN 0-471-13906-8. [Google Scholar]
- Streitel, S.G. Fluorescent pigments (daylight). In Kirk-Othmer Encyclopedia of Chemical Technology; John Wiley and Sons: New York, NY, USA, 2009; Volume 15, pp. 584–607. [Google Scholar]
- Rice, K.B.; Jones, S.K.; Morrison, W.; Leskey, T.C. Spotted wing Drosophila prefer low hanging fruit: Insights into foraging behavior and management strategies. J. Insect Behav. 2017, 30, 645–661. [Google Scholar] [CrossRef]
- Kirkpatrick, D.M.; Gut, L.J.; Miller, J.R. Estimating monitoring trap plume reach and trapping area for Drosophila suzukii (Diptera: Drosophilidae) in Michigan tart cherry. J. Econ. Entomol. 2018, 111, 1285–1289. [Google Scholar] [CrossRef]
- Drummond, F.A.; Ballman, E.; Collins, J.A. Spotted-wing Drosophila (Diptera: Drosophilidae) adult movement, activity, and oviposition behavior in Maine wild blueberry (Vaccinium angustifolium; Ericales: Ericaceae). J. Econ. Entomol. 2019, 112, 1623–1633. [Google Scholar] [CrossRef]
- Crumpacker, D.W. The use of micronized fluorescent dusts to mark adult Drosophila pseudoobscura. Am. Midl. Nat. 1974, 91, 118–129. [Google Scholar] [CrossRef]
- Kilgour, R.J.; McAdam, A.G.; Betini, G.S.; Norris, D.R. Experimental evidence that density mediates negative frequency-dependent selection on aggression. J. Anim. Ecol. 2018, 87, 1091–1101. [Google Scholar] [CrossRef]
- Shenoi, V.N.; Prasad, N.G. Local adaptation to developmental density does not lead to higher mating success in Drosophila melanogaster. J. Evol. Biol. 2016, 29, 2036–2042. [Google Scholar] [CrossRef] [PubMed]
- Cooper, B.S.; Sedghifar, A.; Nash, W.T.; Comeault, A.A.; Matute, D.R. A maladaptive combination of traits contributes to the maintenance of a Drosophila hybrid zone. Curr. Biol. 2018, 28, 2940–2947. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A.; Ratna, E.; Sgrò, C.M.; Barton, M.; Blacket, M.; Hallas, R.; De Garis, S.; Weeks, A.R. Antagonistic selection between adult thorax and wing size in field released Drosophila melanogaster independent of thermal conditions. J. Evol. Biol. 2007, 20, 2219–2227. [Google Scholar] [CrossRef] [PubMed]
- Etges, W.J. Premating isolation is determined by larval substrates in cactophilic Drosophila mojavensis. Evolution (N. Y.) 1992, 46, 1945. [Google Scholar]
- Etges, W.J. Premating isolation is determined by larval rearing substrates in cactophilic Drosophila mojavensis. IV. Correlated responses in behavioral isolation to artificial selection on a life-history trait. Am. Nat. 1998, 152, 129–144. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A.; Parsons, P.A. Inter- and intraspecific variation in the response of Drosophila melanogaster and D. simulans to larval cues. Behav. Genet. 1986, 16, 295–306. [Google Scholar] [CrossRef]
- Feder, M.E.; Roberts, S.P.; Bordelon, A.C. Molecular thermal telemetry of free-ranging adult Drosophila melanogaster. Oecologia 2000, 123, 460–465. [Google Scholar] [CrossRef]
- Coyne, J.A.; Bryant, S.H.; Turelli, M. Long-distance migration of Drosophila. 2. Presence in desolate sites and dispersal near a desert oasis. Am. Nat. 1987, 129, 847–861. [Google Scholar] [CrossRef]
- Markow, T.A.; Castrezana, S. Dispersal in cactophilic Drosophila. Oikos 2000, 89, 378–386. [Google Scholar] [CrossRef]
- Begon, M. Dispersal, density and microdistribution in Drosophila subobscura Collin. J. Anim. Ecol. 1976, 45, 441–456. [Google Scholar] [CrossRef]
- Begon, M.; Milburn, O.; Turner, D. Density estimates of Drosophila in Southern England. J. Nat. Hist. 1975, 9, 315–320. [Google Scholar] [CrossRef]
- Johnston, J.S.; Heed, W.B. Dispersal of Drosophila: The effect of baiting on the behavior and distribution of natural populations. Am. Nat. 1975, 109, 207–216. [Google Scholar] [CrossRef]
- Turelli, M.; Burkhard, C.; Fong, V.; Moore, J.; Van Horn, S.; Prout, T. Does dusting distort Drosophila dispersal? Drosoph. Inf. Serv. 1986, 63, 131–132. [Google Scholar]
- McKenzie, J.A. The distribution of vineyard populations of Drosophila melanogaster and Drosophila simulans during vintage and non-vintage periods. Oecologia 1974, 15, 1–16. [Google Scholar] [CrossRef]
- Crumpacker, D.W.; Williams, J.S. Density, dispersion, and population structure in Drosophila pseudoobscura. Ecol. Monogr. 1973, 43, 499–538. [Google Scholar] [CrossRef]
- Powell, J.R.; Dobzhansky, T.; Hook, J.E.; Wistrand, H.E. Genetics of natural populations. XLIII. Further studies on rates of dispersal of Drosophila pseudoobscura and its relatives. Genetics 1976, 82, 493–506. [Google Scholar]
- Dobzhansky, T.; Powell, J.R.; Taylor, C.E.; Andregg, M. Ecological variables affecting the dispersal behavior of Drosophila pseudoobscura and its relatives. Am. Nat. 1979, 114, 325–334. [Google Scholar] [CrossRef]
- Dobzhansky, T.; Powell, J.R. Rates of dispersal of Drosophila pseudoobscura and its relatives. Proc. R. Soc. London - Biol. Sci. 1974, 187, 281–298. [Google Scholar]
- Schirra, K.-J.; Alexander, S.; Griebeler, E.M.; Johannesen, J. Forschungsring des Deutschen Weinbaus bei der DLG e.V.: Jahresbericht 2012; DLG e.V. Fachzentrum Land- und Ernährungswirtschaft Frankfurt: Frankfurt am Main, 2012. [Google Scholar]
- Cardoso Pereira, R.; (Joint FAO/IAEA Division of Nuclear Techniques in Food and Agriculture, Vienna, Austria). Personal communication, 2019.
- Taning, C.N.T.; Christiaens, O.; Berkvens, N.; Casteels, H.; Maes, M.; Smagghe, G. Oral RNAi to control Drosophila suzukii: Laboratory testing against larval and adult stages. J. Pest Sci. 2016, 89, 803–814. [Google Scholar] [CrossRef]
- Townsend, G.R.; Heuberger, J.W. Methods for estimating losses caused by diseases in fungicide experiments. Plant Dis. Report. 1943, 27, 340–343. [Google Scholar]
- Clymans, R.; Van Kerckvoorde, V.; Bangels, E.; Akkermans, W.; Alhmedi, A.; De Clercq, P.; Beliën, T.; Bylemans, D. Olfactory preference of Drosophila suzukii shifts between fruit and fermentation cues over the season: Effects of physiological status. Insects 2019, 10, 200. [Google Scholar] [CrossRef] [PubMed]
- Panel, A.D.C.; Zeeman, L.; van der Sluis, B.J.; van Elk, P.; Pannebakker, B.A.; Wertheim, B.; Helsen, H.H.M. Overwintered Drosophila suzukii are the main source for infestations of the first fruit crops of the season. Insects 2018, 9, 145. [Google Scholar] [CrossRef] [PubMed]
- Van Kerckvoorde, V.; Clymans, R.; Bangels, E.; Alhmedi, A.; De Ketelaere, B.; De Clercq, P.; Bylemans, D.; Belien, T. Improved trapping efficacy, by tunnel entries and an insecticide-coated inner surface as killing agent, uncovers the importance of fly retention in Drosophila suzukii traps. Pest Manag. Sci. 2020. (under review). [Google Scholar]
- Rossi-Stacconi, M.V.; Kaur, R.; Mazzoni, V.; Ometto, L.; Grassi, A.; Gottardello, A.; Rota-Stabelli, O.; Anfora, G. Multiple lines of evidence for reproductive winter diapause in the invasive pest Drosophila suzukii: Useful clues for control strategies. J. Pest Sci. 2016, 89, 689–700. [Google Scholar] [CrossRef]
- Harris, D.W.; Hamby, K.A.; Wilson, H.E.; Zalom, F.G. Seasonal monitoring of Drosophila suzukii (Diptera: Drosophilidae) in a mixed fruit production system. J. Asia. Pac. Entomol. 2014, 17, 857–864. [Google Scholar] [CrossRef]
- Cha, D.H.; Hesler, S.P.; Wallingford, A.K.; Zaman, F.; Jentsch, P.; Nyrop, J.; Loeb, G.M. Comparison of commercial lures and food baits for early detection of fruit infestation risk by Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2018, 111, 645–652. [Google Scholar] [CrossRef]
- Cha, D.H.; Adams, T.; Werle, C.T.; Sampson, B.J.; Adamczyk, J.J.; Rogg, H.; Landolt, P.J. A four-component synthetic attractant for Drosophila suzukii (Diptera: Drosophilidae) isolated from fermented bait headspace. Pest Manag. Sci. 2014, 70, 324–331. [Google Scholar] [CrossRef]
- Burrack, H.J.; Asplen, M.; Bahder, L.; Collins, J.; Drummond, F.A.; Guédot, C.; Isaacs, R.; Johnson, D.; Blanton, A.; Lee, J.C.; et al. Multistate comparison of attractants for monitoring Drosophila suzukii (Diptera: Drosophilidae) in blueberries and caneberries. Environ. Entomol. 2015, 44, 704–712. [Google Scholar] [CrossRef]
- Tonina, L.; Grassi, A.; Caruso, S.; Mori, N.; Gottardello, A.; Anfora, G.; Giomi, F.; Vaccari, G.; Ioriatti, C. Comparison of attractants for monitoring Drosophila suzukii in sweet cherry orchards in Italy. J. Appl. Entomol. 2018, 142, 18–25. [Google Scholar] [CrossRef]
- Baroffio, C.A.; Richoz, P.; Fischer, S.; Kuske, S.; Linder, C.; Kehrli, P. Monitoring Drosophila suzukii in Switzerland in 2012. J. Berry Res. 2014, 4, 47–51. [Google Scholar] [CrossRef]
- Arnó, J.; Solà, M.; Riudavets, J.; Gabarra, R. Population dynamics, non-crop hosts, and fruit susceptibility of Drosophila suzukii in Northeast Spain. J. Pest Sci. 2016, 89, 713–723. [Google Scholar] [CrossRef]
- Briem, F.; Dominic, A.R.; Golla, B.; Hoffmann, C.; Englert, C.; Herz, A.; Vogt, H. Explorative data analysis of Drosophila suzukii trap catches from a seven-year monitoring program in Southwest Germany. Insects 2018, 9, 125. [Google Scholar] [CrossRef] [PubMed]
- Mazzetto, F.; Pansa, M.G.; Ingegno, B.L.; Tavella, L.; Alma, A. Monitoring of the exotic fly Drosophila suzukii in stone, pome and soft fruit orchards in NW Italy. J. Asia. Pac. Entomol. 2015, 18, 321–329. [Google Scholar] [CrossRef]
- Ioriatti, C.; Walton, V.; Dalton, D.; Anfora, G.; Grassi, A.; Maistri, S.; Mazzoni, V. Drosophila suzukii (Diptera: Drosophilidae) and its potential impact to wine grapes during harvest in two cool climate wine grape production regions. J. Econ. Entomol. 2015, 108, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Tait, G.; Grassi, A.; Pfab, F.; Crava, C.M.; Dalton, D.T.; Magarey, R.; Ometto, L.; Vezzulli, S.; Rossi-Stacconi, M.V.; Gottardello, A.; et al. Large-scale spatial dynamics of Drosophila suzukii in Trentino, Italy. J. Pest Sci. 2018, 91, 1213–1224. [Google Scholar] [CrossRef]
- Tochen, S.; Dalton, D.T.; Wiman, N.; Hamm, C.; Shearer, P.W.; Walton, V.M. Temperature-related development and population parameters for Drosophila suzukii (Diptera: Drosophilidae) on cherry and blueberry. Environ. Entomol. 2014, 43, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.G.; Stewart, T.J.; Biondi, A.; Chavez, B.A.; Ingels, C.; Caprile, J.; Grant, J.A.; Walton, V.M.; Daane, K.M. Population dynamics and ecology of Drosophila suzukii in Central California. J. Pest Sci. 2016, 89, 701–712. [Google Scholar] [CrossRef]
- Vacas, S.; Primo, J.; Manclús, J.J.; Montoya, Á.; Navarro-Llopis, V. Survey on Drosophila suzukii natural short-term dispersal capacities using the mark−release−recapture technique. Insects 2019, 10, 268. [Google Scholar] [CrossRef]
- Kirkpatrick, D.M.; McGhee, P.S.; Gut, L.J.; Miller, J.R. Improving monitoring tools for spotted wing drosophila, Drosophila suzukii. Entomol. Exp. Appl. 2017, 164, 87–93. [Google Scholar] [CrossRef]
- Lasa, R.; Tadeo, E.; Toledo-Hérnandez, R.A.; Carmona, L.; Lima, I.; Williams, T. Improved capture of Drosophila suzukii by a trap baited with two attractants in the same device. PLoS One 2017, 12, e0188350. [Google Scholar] [CrossRef]
- Ebbenga, D.N.; Burkness, E.C.; Hutchison, W.D.; Rodriguez-Saona, C. Evaluation of exclusion netting for spotted-wing Drosophila (Diptera: Drosophilidae) management in Minnesota wine grapes. J. Econ. Entomol. 2019, 112, 2287–2294. [Google Scholar] [CrossRef] [PubMed]
- Wiman, N.G.; Dalton, D.T.; Anfora, G.; Biondi, A.; Chiu, J.C.; Daane, K.M.; Gerdeman, B.; Gottardello, A.; Hamby, K.A.; Isaacs, R.; et al. Drosophila suzukii population response to environment and management strategies. J. Pest Sci. 2016, 89, 653–665. [Google Scholar] [CrossRef]
- Van Timmeren, S.; Isaacs, R. Control of spotted wing Drosophila, Drosophila suzukii, by specific insecticides and by conventional and organic crop protection programs. Crop Prot. 2013, 54, 126–133. [Google Scholar] [CrossRef]
- Klick, J.; Yang, W.Q.; Walton, V.M.; Dalton, D.T.; Hagler, J.R.; Dreves, A.J.; Lee, J.C.; Bruck, D.J. Distribution and activity of Drosophila suzukii in cultivated raspberry and surrounding vegetation. J. Appl. Entomol. 2016, 140, 37–46. [Google Scholar] [CrossRef]
- Leach, H.; Hagler, J.R.; Machtley, S.A.; Isaacs, R. Spotted wing Drosophila (Drosophila suzukii) utilization and dispersal from the wild host Asian bush honeysuckle (Lonicera spp.). Agric. For. Entomol. 2019, 21, 149–158. [Google Scholar] [CrossRef]
- Betz, D.; Rauleder, H.; Köppler, K.; Eben, A.; Vogt, H. Markierungsversuche zur Ermittlung des Ausbreitungsverhaltens der Kirschessigfliege (Drosophila suzukii) in Abhängigkeit der Habitatstruktur im Oberrheingebiet 1. Available online: https://www.julius-kuehn.de/media/Institute/OW/KrankheitenSchaedlinge/PDF/1_Migration_InterregInvaprotect_Markierungsversuche_Ausbreitungsverhalten_Dsuzukii_2017_Versuchsbericht1_.pdf (accessed on 30 October 2019).
- Haag, N.; Betz, D.; Hauck, S.; Rauleder, H.; Köppler, K.; Eben, A.; Braun, T.; Vogt, H. Markierungsversuche zur Ermittlung des Ausbreitungsverhaltens der Kirschessigfliege in Abhängigkeit der Habitatstruktur im Oberrheingebiet 2. Available online: https://www.julius-kuehn.de/media/Institute/OW/KrankheitenSchaedlinge/PDF/2_Migration_InterregInvaprotect_Markierungsversuchezumausbreitungsverhalten_Dsuzukii__2018_Versuchsbericht2_.pdf (accessed on 30 October 2019).
- Epopa, P.S.; Millogo, A.A.; Collins, C.M.; North, A.; Tripet, F.; Benedict, M.Q.; Diabate, A. The use of sequential mark-release-recapture experiments to estimate population size, survival and dispersal of male mosquitoes of the Anopheles gambiae complex in Bana, a west African humid savannah village. Parasit. Vectors 2017, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Gouagna, L.C.; Dehecq, J.S.; Fontenille, D.; Dumont, Y.; Boyer, S. Seasonal variation in size estimates of Aedes albopictus population based on standard mark-release-recapture experiments in an urban area on Reunion Island. Acta Trop. 2015, 143, 89–96. [Google Scholar] [CrossRef]
- Lux, S.A.; Wnuk, A.; Vogt, H.; Belien, T.; Spornberger, A.; Studnicki, M. Validation of individual-based Markov-like stochastic process model of insect behavior and a “virtual farm” concept for enhancement of site-specific IPM. Front. Physiol. 2016, 7, 1–20. [Google Scholar] [CrossRef]
- Kirkpatrick, D.M.; Acebes-Doria, A.L.; Rice, K.B.; Short, B.D.; Adams, C.G.; Gut, L.J.; Leskey, T.C.; Stelinski, L. Estimating monitoring trap plume reach and trapping area for nymphal and adult Halyomorpha halys (Hemiptera: Pentatomidae) in crop and non-crop habitats. Environ. Entomol. 2019, 48, 1104–1112. [Google Scholar] [CrossRef]
- Lance, D.R.; Gates, D.B. Sensitivity of detection trapping systems for Mediterranean fruit flies (Diptera: Tephritidae) in southern California. J. Econ. Entomol. 1994, 87, 1377–1383. [Google Scholar] [CrossRef]
- Manoukis, N.C.; Hall, B.; Geib, S.M. A computer model of insect traps in a landscape. Sci. Rep. 2014, 4, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, R.T.; Couey, H.M. Mediterranean fruit fly (Diptera: Tephritidae): Distance/response curves to trimedlure to measure trapping efficiency. Environ. Entomol. 1986, 15, 71–74. [Google Scholar] [CrossRef]
- Parker, A.G. Mass-Rearing for Sterile Insect Release; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; ISBN 9781402040511. [Google Scholar]
- Krüger, A.P.; Schlesener, D.C.H.; Martins, L.N.; Wollmann, J.; Deprá, M.; Garcia, F.R.M. Effects of irradiation dose on sterility induction and quality parameters of Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2018, 111, 741–746. [Google Scholar] [CrossRef] [PubMed]
- Krüger, A.P.; Schlesener, D.C.H.; Martins, L.N.; Wollmann, J.; Deprá, M.; Garcia, F.R.M. Radiation effects on Drosophila suzukii (Diptera: Drosophilidae) reproductive behaviour. J. Appl. Entomol. 2019, 143, 88–94. [Google Scholar] [CrossRef]
- Lanouette, G.; Brodeur, J.; Fournier, F.; Martel, V.; Vreysen, M.; Cáceres, C.; Firlej, A. The sterile insect technique for the management of the spotted wing Drosophila, Drosophila suzukii: Establishing the optimum irradiation dose. PLoS One 2017, 12, e0180821. [Google Scholar] [CrossRef]
- Nikolouli, K.; Colinet, H.; Renault, D.; Enriquez, T.; Mouton, L.; Gibert, P.; Sassu, F.; Cáceres, C.; Stauffer, C.; Pereira, R.; et al. Sterile insect technique and Wolbachia symbiosis as potential tools for the control of the invasive species Drosophila suzukii. J. Pest Sci. 2018, 91, 489–503. [Google Scholar] [CrossRef]
- RADIANT COLOR NV Technical Data Sheet Radglo® TP; RADIANT COLOR NV: Houthalen-Helchteren, Belgium, 2017.
- RADIANT COLOR NV Production Promotion Sheet Radglo® TP; RADIANT COLOR NV: Houthalen-Helchteren, Belgium, 2017.
- RADIANT COLOR NV Personal Communication; RADIANT COLOR NV: Houthalen-Helchteren, Belgium, 2018.
- SWADA Technical data sheets. Available online: http://www.swada.co.uk/ (accessed on 25 November 2019).
- RADIANT COLOR NV Safety Data Sheet TP-40; RADIANT COLOR NV: Houthalen-Helchteren, Belgium, 2017.
- Southwood, T.R.E. Ecological Methods with Particular Reference to the Study of Insect Populations; Chapman and Hall: New York, NY, USA, 1987; ISBN 9780412307102. [Google Scholar]
- Moffitt, H.R.; Albano, D.J. Codling moths: Fluorescent powders as markers. Environ. Entomol. 1972, 1, 750–753. [Google Scholar] [CrossRef]
- Wong, T.T.Y.; Cleveland, M.L. Fluorescent powder for marking deciduous fruit moths for studies of dispersal. J. Econ. Entomol. 1970, 63, 339–340. [Google Scholar] [CrossRef]
- Dowdy, A.K.; McGaughey, W.H. Fluorescent pigments for marking lesser grain borers (Coleoptera: Bostrichidae). J. Econ. Entomol. 1992, 85, 567–569. [Google Scholar] [CrossRef]
- Lin, A.Q.; Zhai, Y.; Zhang, A.; Men, X.; Zhang, X. Comparative developmental times and laboratory life tables for Drosophila suzukii and Drosophila melanogaster (Diptera: Drosophilidae). Florida Entomol. 2014, 97, 1434–1442. [Google Scholar] [CrossRef]
- Tochen, S.; Woltz, J.M.; Dalton, D.T.; Lee, J.C.; Wiman, N.G.; Walton, V.M. Humidity affects populations of Drosophila suzukii (Diptera: Drosophilidae) in blueberry. J. Appl. Entomol. 2016, 140, 47–57. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, J.S.; Park, J.S.; Choi, D.S.; Park, J.; Kim, I. Oviposition and development potential of the spotted-wing Drosophila, Drosophila suzukii (Diptera: Drosophilidae), on uninjured Campbell Early grape. Entomol. Res. 2015, 45, 354–359. [Google Scholar] [CrossRef]
- Bennett, S.R.; McClelland, G.A.H.; Smilanick, J.M. A versatile system of fluorescent marks for studies of large populations of mosquitoes (Diptera: Culicidae). J. Med. Entomol. 1981, 18, 173–174. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clymans, R.; Van Kerckvoorde, V.; Beliën, T.; Bylemans, D.; De Clercq, P. Marking Drosophila suzukii (Diptera: Drosophilidae) with Fluorescent Dusts. Insects 2020, 11, 152. https://doi.org/10.3390/insects11030152
Clymans R, Van Kerckvoorde V, Beliën T, Bylemans D, De Clercq P. Marking Drosophila suzukii (Diptera: Drosophilidae) with Fluorescent Dusts. Insects. 2020; 11(3):152. https://doi.org/10.3390/insects11030152
Chicago/Turabian StyleClymans, Rik, Vincent Van Kerckvoorde, Tim Beliën, Dany Bylemans, and Patrick De Clercq. 2020. "Marking Drosophila suzukii (Diptera: Drosophilidae) with Fluorescent Dusts" Insects 11, no. 3: 152. https://doi.org/10.3390/insects11030152
APA StyleClymans, R., Van Kerckvoorde, V., Beliën, T., Bylemans, D., & De Clercq, P. (2020). Marking Drosophila suzukii (Diptera: Drosophilidae) with Fluorescent Dusts. Insects, 11(3), 152. https://doi.org/10.3390/insects11030152