Oviposition Preference of the Cabbage Root Fly towards Some Chinese Cabbage Cultivars: A Search for Future Trap Crop Candidates

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chinese Cabbage Diversity
2.2. Laboratory Choice Experiment
2.3. Field Choice Experiment
2.4. Statistical Analysis
3. Results
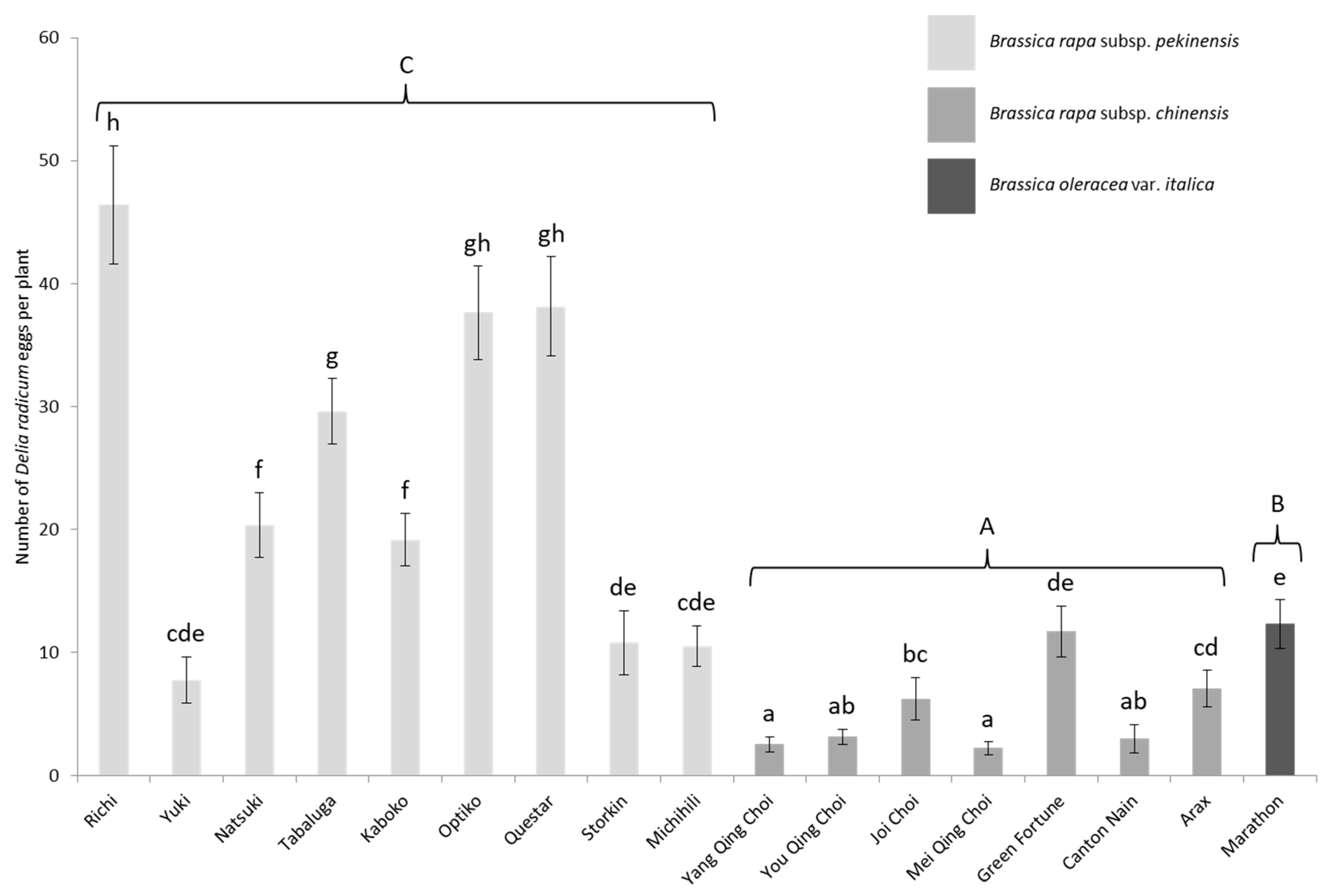
3.1. Laboratory Choice Experiments
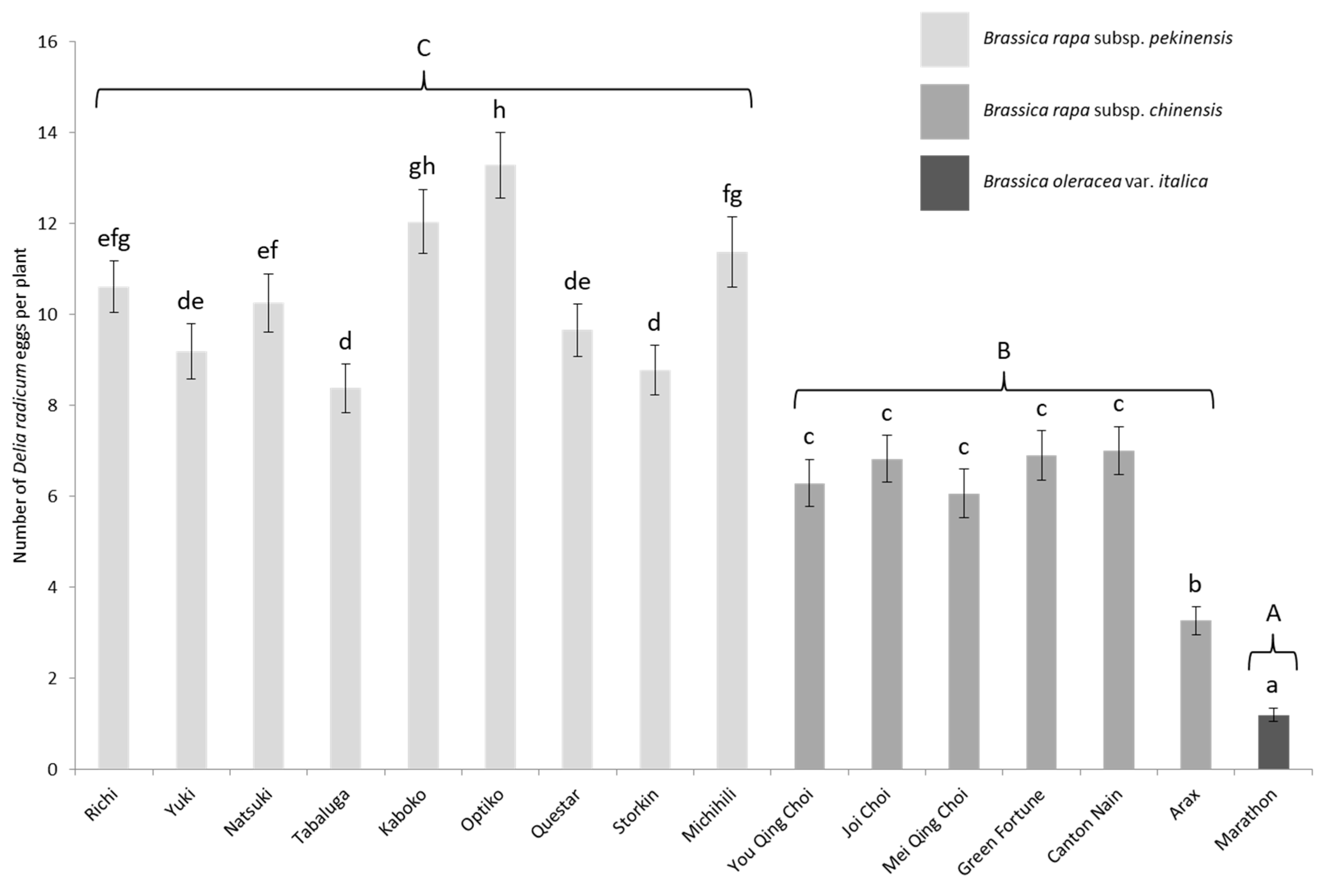
3.2. Field Choice Experiment
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bernays, E.; Graham, M. On the evolution of host specificity in phytophagous arthropods. Ecology 1988, 69, 886–892. [Google Scholar] [CrossRef]
- Thorsteinson, A.J. Host selection in phytophagous insects. Annu. Rev. Entomol. 1960, 5, 193–218. [Google Scholar] [CrossRef]
- Kennedy, J.S. Mechanisms of host plant selection. Ann. Appl. Biol. 1965, 56, 317–322. [Google Scholar] [CrossRef]
- Craig, T.P.; Itami, J.K.; Price, P.W. A strong relationship between oviposition preference and larval performance in a shoot-galling sawfly. Ecology 1989, 70, 1691–1699. [Google Scholar] [CrossRef]
- Åhman, I. Oviposition and larval performance of Rabdophaga terminalis on Salix spp. with special consideration to bud size of host plants. Entomol. Exp. Appl. 1984, 35, 129–136. [Google Scholar] [CrossRef]
- García-Robledo, C.; Horvitz, C.C. Parent–offspring conflicts, “optimal bad motherhood” and the “mother knows best” principles in insect herbivores colonizing novel host plants. Ecol. Evol. 2012, 2, 1446–1457. [Google Scholar] [CrossRef]
- Olazcuaga, L.; Rode, N.O.; Foucaud, J.; Facon, B.; Ravigné, V.; Ausset, A.; Leménager, N.; Loiseau, A.; Gautier, M.; Estoup, A.; et al. Oviposition preference and larval performance of Drosophila suzukii (Diptera: Drosophilidae), spotted-wing Drosophila: Effects of fruit identity and composition. Environ. Entomol. 2019, 48, 867–881. [Google Scholar] [CrossRef]
- Courtney, S.P. Coevolution of pierid butterflies and their cruciferous foodplants. Oecologia 1981, 51, 91–96. [Google Scholar] [CrossRef]
- Martin, A.D.; Stanley-Horn, D.; Hallett, R.H. Adult host preference and larval performance of Liriomyza huidobrensis (Diptera: Agromyzidae) on selected hosts. Environ. Entomol. 2005, 34, 1170–1177. [Google Scholar] [CrossRef][Green Version]
- Smiley, J. Plant chemistry and the evolution of host specificity: New evidence from Heliconius and Passiflora. Science 1978, 201, 745–747. [Google Scholar] [CrossRef]
- Wiklund, C. The evolutionary relationship between adult oviposition preferences and larval host plant range in Papilio machaon L. Oecologia 1975, 18, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, J. Oviposition preference and larval survival of Chilo partellus (Lepidoptera : Pyralidae) on Napier grass (Pennisetum purpureum) trap crops. Int. J. Pest Manag. 2006, 52, 39–44. [Google Scholar] [CrossRef]
- Mayhew, P.J. Herbivore host choice and optimal bad motherhood. Trends Ecol. Evol. 2001, 16, 165–167. [Google Scholar] [CrossRef]
- Gripenberg, S.; Mayhew, P.J.; Parnell, M.; Roslin, T. A meta-analysis of preference–performance relationships in phytophagous insects. Ecol. Lett. 2010, 13, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Charlery de la Masselière, M.; Facon, B.; Hafsi, A.; Duyck, P.F. Diet breadth modulates preference—performance relationships in a phytophagous insect community. Sci. Rep. UK 2017, 7, 16934. [Google Scholar] [CrossRef] [PubMed]
- Hokkanen, H.M.T. Trap cropping in pest-management. Annu. Rev. Entomol. 1991, 36, 119–138. [Google Scholar] [CrossRef]
- Javaid, I.; Joshi, J.M. Trap cropping in insect pest-management. J. Sustain. Agric. 1995, 5, 117–136. [Google Scholar] [CrossRef]
- Veromann, E.; Kaasik, R.; Kovacs, G.; Metspalu, L.; Williams, I.H.; Mand, M. Fatal attraction: Search for a dead-end trap crop for the pollen beetle (Meligethes aeneus). Arthropod-Plant Int. 2014, 8, 373–381. [Google Scholar] [CrossRef]
- Khan, Z.R.; Midega, C.A.O.; Hutter, N.J.; Wilkins, R.M.; Wadhams, L.J. Assessment of the potential of Napier grass (Pennisetum purpureum) varieties as trap plants for management of Chilo partellus. Entomol. Exp. Appl. 2006, 119, 15–22. [Google Scholar] [CrossRef]
- Newman, K.; You, M.; Vasseur, L. Diamondback moth (Lepidoptera: Plutellidae) exhibits oviposition and larval feeding preferences among crops, wild plants, and ornamentals as host plants. J. Econ. Entomol. 2016, 109, 644–648. [Google Scholar] [CrossRef]
- Kumar, S. Potential of Ethiopian mustard, Brassica carinata as a trap crop for large white butterfly, Pieris brassicae infesting Indian mustard, Brassica juncea. J. Pest Sci. 2017, 90, 129–137. [Google Scholar] [CrossRef]
- Finch, S.; Ackley, C.M. Cultivated and wild host plants supporting populations of the cabbage root fly. Ann. Appl. Biol. 1977, 85, 13–22. [Google Scholar] [CrossRef]
- Shuhang, W.; Voorrips, R.E.; Steenhuis-Broers, G.; Vosman, B.; van Loon, J.J.A. Antibiosis resistance against larval cabbage root fly, Delia radicum, in wild Brassica-species. Euphytica 2016, 211, 139–155. [Google Scholar] [CrossRef]
- King, K.M.; Forbes, A.R. Control of root maggots in rutabagas. J. Econ. Entomol. 1954, 47, 607–615. [Google Scholar] [CrossRef]
- Coaker, T.H.; Finch, S. The cabbage root fly, Erioischia brassicae (Bouché). Rep. Natl. Veg. Res. Stn. 1970 1971, 71, 23–42. [Google Scholar]
- Brühl, C.A.; Zaller, J.G. Biodiversity decline as a consequence of an inappropriate environmental risk assessment of pesticides. Front. Environ. Sci. 2019, 7. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Kesavachandran, C. Health Effects of Pesticides; CRC Press: London, UK, 2019. [Google Scholar] [CrossRef]
- Radcliffe, E.B.; Keith Chapman, R. Varietal resistance to insect attack in various cruciferous crops. J. Econ. Entomol. 1966, 59, 120–125. [Google Scholar] [CrossRef]
- Doane, J.F.; Chapman, R.K. Oviposition Preference of the Cabbage Maggot, Hylemya brassicae (Bouché). J. Econ. Entomol. 1962, 55, 137–138. [Google Scholar] [CrossRef]
- Mukerji, M.K. Oviposition preference and survival of Hylemya brassicae on some cruciferous crops. Can. Entomol. 1969, 101, 153–158. [Google Scholar] [CrossRef]
- Rousse, P.; Fournet, S.; Porteneuve, C.; Brunel, E. Trap cropping to control Delia radicum populations in cruciferous crops: First results and future applications. Entomol. Exp. Appl. 2003, 109, 133–138. [Google Scholar] [CrossRef]
- Kergunteuil, A. Des odeurs pour protéger les cultures : Utilisation de composés volatils pour modifier le comportement de la mouche du chou, Delia radicum et de ses Ennemis Naturels. Ph.D. Thesis, Biology, Université de Rennes 1, Rennes, France, 2013. [Google Scholar]
- Lamy, F.; Dugravot, S.; Cortesero, A.M.; Chaminade, V.; Faloya, V.; Poinsot, D. One more step toward a push-pull strategy combining both a trap crop and plant volatile organic compounds against the cabbage root fly Delia radicum. Environ. Sci. Pollut. R 2017, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lamy, F.; Poinsot, D.; Cortesero, A.M.; Dugravot, S. Artificially applied plant volatile organic compounds modify the behavior of a pest with no adverse effect on its natural enemies in the field. J. Pest Sci. 2016, 1–11. [Google Scholar] [CrossRef]
- Kergunteuil, A.; Cortesero, A.M.; Chaminade, V.; Dourlot, S.; Paty, C.; Le Ralec, A.; Dugravot, S. Field and laboratory selection of brassicaceous plants that differentially affect infestation levels by Delia radicum. J. Appl. Entomol. 2015, 139, 487–495. [Google Scholar] [CrossRef]
- Mesmin, X.; Vincent, M.; Tricault, Y.; Estorgues, V.; Daniel, L.; Cortesero, A.-M.; Faloya, V.; Le Ralec, A. Assessing the relationship between pest density and plant damage: A case study with the belowground herbivore Delia radicum (Diptera: Anthomyiidae) on broccoli. Appl. Entomol. Zool. 2019. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics With S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; SAGE: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Lenth, R. Emmeans: Estimated Marginal Means, aka Least-Squares Means. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 5 February 2020).
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. Roy. Stat. Soc. B Met. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Pond, D.D.; Moore, C.A. Oviposition of Hylemya brassicae and H. Cilicrura around several brassica crops. Can. J. Plant Sci. 1965, 45, 392–394. [Google Scholar] [CrossRef]
- Ellis, P.R.; Taylor, J.D.; Littlejohn, I.H. The role of microorganisms colonising radish seedlings in the oviposition behaviour of cabbage root fly, Delia radicum. In Proceedings of the 5th International Symposium on Insect Plant Relationships, Wageningen, the Netherlands, 1–4 March 1982; pp. 131–137. [Google Scholar]
- Dosdall, L.M.; Herbut, M.J.; Cowle, N.T. Susceptibilities of species and cultivars of canola and mustard to infestation by root maggots (Delia spp.) (Diptera: Anthomyiidae). Can. Entomol. 1994, 126, 251–260. [Google Scholar] [CrossRef]
- Kostal, V. Physical and chemical factors influencing landing and oviposition by the cabbage root fly on host-plant models. Entomol. Exp. Appl. 1993, 66, 109–118. [Google Scholar] [CrossRef]
- Finch, S. Volatile plant-chemicals and their effect on host plant finding by the cabbage root fly (Delia brassicae). Entomol. Exp. Appl. 1978, 24, 350–359. [Google Scholar] [CrossRef]
- Hawkes, C.; Coaker, T.H. Behavioural responses to host-plant odours in adult cabbage root fly (Erioischia brassicae (Bouché)). In The Host-Plant in Relation to Insect Behaviour and Reproduction; Jermy, T., Ed.; Springer US: Boston, MA, USA, 1976; pp. 85–89. [Google Scholar] [CrossRef]
- Prokopy, R.J.; Collier, R.H.; Finch, S. Leaf color used by cabbage root flies to distinguish among host plants. Science 1983, 221, 190–192. [Google Scholar] [CrossRef]
- Roessingh, P.; Stadler, E.; Fenwick, G.R.; Lewis, J.A.; Nielsen, J.K.; Hurter, J.; Ramp, T. Oviposition and tarsal chemoreceptors of the cabbage root fly are stimulated by glucosinolates and host plant extracts. Entomol. Exp. Appl. 1992, 65, 267–282. [Google Scholar] [CrossRef]
- Roessingh, P.; Stadler, E.; Baur, R.; Hurter, J.; Ramp, T. Tarsal chemoreceptors and oviposition behaviour of the cabbage root fly (Delia radicum) sensitive to fractions and new compounds of host-leaf surface extracts. Physiol. Entomol. 1997, 22, 140–148. [Google Scholar] [CrossRef]
- Nottingham, S.F. Anemotactic strategy and optimum flight conditions for host-plant finding in the cabbage root fly, Delia radicum (L.) (Diptera: Anthomyiidae). B Entomol. Res. 1987, 77, 271–277. [Google Scholar] [CrossRef]
- Hawkes, C. Dispersal of adult cabbage root fly (Erioischia brassicae (Bouché)) in relation to a Barassica crop. J. Appl. Ecol. 1974, 11, 83–93. [Google Scholar] [CrossRef]
- Gouinguene, S.P.D.; Poiger, T.; Stadler, E. Eggs of cabbage root fly stimulate conspecific oviposition: Evaluation of the activity and determination of an egg-associated compound. Chemoecology 2006, 16, 107–113. [Google Scholar] [CrossRef]
- Wallbank, B.E.; Wheatley, G.A. Some responses of cabbage root fly (Delia brassicae) to ally isothiocyanate and other volatile constituents of crucifers. Ann. Appl. Biol. 1979, 91, 1–12. [Google Scholar] [CrossRef]
- Nottingham, S.F.; Coaker, T.H. Changes in flight track angles of cabbage root fly, Delia radicum, in diffuse clouds and discrete plumes of the host-plant volatile allylisothiocyanate. Entomol. Exp. Appl. 1987, 43, 275–278. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, H.; Umashankar, S.; Liang, X.; Lee, H.; Swarup, S.; Ong, C. Characterization of plant volatiles reveals distinct metabolic profiles and pathways among 12 Brassicaceae vegetables. Metabolites 2018, 8, 94. [Google Scholar] [CrossRef]
- Kergunteuil, A.; Dugravot, S.; Mortreuil, A.; Le Ralec, A.; Cortesero, A.M. Selecting volatiles to protect brassicaceous crops against the cabbage root fly, Delia radicum. Entomol. Exp. Appl. 2012, 144, 69–77. [Google Scholar] [CrossRef]
- Roessingh, P.; Stadler, E. Foliar form, colour and surface characteristics influence oviposition behaviour in the cabbage root fly Delia radicum. Entomol. Exp. Appl. 1990, 57, 93–100. [Google Scholar] [CrossRef]
- Zohren, E. Laboruntersuchungen zu massenanzucht, lebensweise, eiablage und eiablageverhalten der Kohlfliege, Chortophila brassicae Bouché (Diptera, Anthomyiidae). J. Appl. Entomol. 1968, 62, 139–188. [Google Scholar] [CrossRef]
- Hurter, J.; Ramp, T.; Patrian, B.; Stadler, E.; Roessingh, P.; Baur, R.; de Jong, R.; Nielsen, J.K.; Winkler, T.; Richter, W.J.; et al. Oviposition stimulants for the cabbage root fly: Isolation from cabbage leaves. Phytochemistry 1999, 51, 377–382. [Google Scholar] [CrossRef]
- Baur, R.; Birch, A.N.E.; Hopkins, R.J.; Griffiths, D.W.; Simmonds, M.S.J.; Stadler, E. Oviposition and chemosensory stimulation of the root flies Delia radicum and D. floralis in response to plants and leaf surface extracts from resistant and susceptible Brassica genotypes. Entomol. Exp. Appl. 1996, 78, 61–75. [Google Scholar] [CrossRef]
- Bhandari, S.R.; Jo, J.S.; Lee, J.G. Comparison of glucosinolate profiles in different tissues of nine Brassica crops. Molecules 2015, 20, 15827–15841. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.; Fenwick, G.R. Glucosinolate content of Brassica vegetables—Chinese cabbages pe-tsai (Brassica-pekinensis) and pak-choi (Brassica-chinensis). J. Sci. Food Agric. 1988, 45, 379–386. [Google Scholar] [CrossRef]
- Kim, J.K.; Chu, S.M.; Kim, S.J.; Lee, D.J.; Lee, S.Y.; Lim, S.H.; Ha, S.H.; Kweon, S.J.; Cho, H.S. Variation of glucosinolates in vegetable crops of Brassica rapa L. ssp. pekinensis. Food Chem. 2010, 119, 423–428. [Google Scholar] [CrossRef]
- Lee, M.K.; Chun, J.H.; Byeon, D.H.; Chung, S.O.; Park, S.U.; Park, S.; Arasu, M.V.; Al-Dhabi, N.A.; Lim, Y.P.; Kim, S.J. Variation of glucosinolates in 62 varieties of Chinese cabbage (Brassica rapa L. ssp pekinensis) and their antioxidant activity. LWT- Food Sci. Technol. 2014, 58, 93–101. [Google Scholar] [CrossRef]
- Hong, E.; Kim, G.H. Variation of glucosinolate composition during seedling and growth stages of Brassica rapa L. ssp pekinensis. Korean J. Hortic. Sci. 2014, 32, 730–738. [Google Scholar] [CrossRef]
- Roessingh, P.; Städler, E.; Hurter, J.; Ramp, T. Oviposition stimulant for the cabbage root fly: Important new cabbage leaf surface compound and specific tarsal receptors. In Proceedings of the 8th International Symposium on Insect-Plant Relationships ; Menken, S.B.J., Visser, J.H., Harrewijn, P., Eds.; Springer: Dordrecht, The Netherlands, 1992; pp. 141–142. [Google Scholar] [CrossRef]
- Kergunteuil, A.; Dugravot, S.; Danner, H.; van Dam, N.M.; Cortesero, A.M. Characterizing volatiles and attractiveness of five brassicaceous plants with potential for a ‘push-pull’ strategy toward the cabbage root fly, Delia radicum. J. Chem. Ecol. 2015, 41, 330–339. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Binomial Name | Subspecies/Variety | Cultivar (Genetics-Seed Provider) | Seed Coating | Number (Figure 1) |
|---|---|---|---|---|
| Brassica rapa | pekinensis | Richi (F1–SV) | Thiram | 1 |
| Yuki (F1–SV) | Thiram | 2 | ||
| Natsuki (F1–SV) | Thiram | 3 | ||
| Tabaluga (F1–SV) | Thiram | 4 | ||
| Kaboko (F1–GB) | None | 5 | ||
| Optiko (F1–GB) | Iprodione | 6 | ||
| Questar (F1–GB) | None | 7 | ||
| Storkin (F1–GB) | None | 8 | ||
| Michihili (Pop–LBG) | None | 9 | ||
| chinensis | Yang Qing Choi (F1–SV) * | Thiram | – | |
| You Qing Choi (F1–SV) | Thiram | 10 | ||
| Joi Choi(F1–SV) | Thiram | 11 | ||
| Mei Qing Choi (F1–SV) | Thiram | 12 | ||
| Green fortune (F1–GB) | Thiram | 13 | ||
| Canton Nain (Pop–GB) | None | 14 | ||
| Arax (Pop–GB) | None | 15 | ||
| Brassica oleracea | italica | Marathon (F1–SV) | None | 16 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lamy, F.; Bellec, L.; Rusu-Stievenard, A.; Clin, P.; Ricono, C.; Olivier, D.; Mauger, S.; Poinsot, D.; Faloya, V.; Daniel, L.; et al. Oviposition Preference of the Cabbage Root Fly towards Some Chinese Cabbage Cultivars: A Search for Future Trap Crop Candidates. Insects 2020, 11, 127. https://doi.org/10.3390/insects11020127
Lamy F, Bellec L, Rusu-Stievenard A, Clin P, Ricono C, Olivier D, Mauger S, Poinsot D, Faloya V, Daniel L, et al. Oviposition Preference of the Cabbage Root Fly towards Some Chinese Cabbage Cultivars: A Search for Future Trap Crop Candidates. Insects. 2020; 11(2):127. https://doi.org/10.3390/insects11020127
Chicago/Turabian StyleLamy, Fabrice, Laura Bellec, Amélie Rusu-Stievenard, Pauline Clin, Claire Ricono, Diane Olivier, Solène Mauger, Denis Poinsot, Vincent Faloya, Loïc Daniel, and et al. 2020. "Oviposition Preference of the Cabbage Root Fly towards Some Chinese Cabbage Cultivars: A Search for Future Trap Crop Candidates" Insects 11, no. 2: 127. https://doi.org/10.3390/insects11020127
APA StyleLamy, F., Bellec, L., Rusu-Stievenard, A., Clin, P., Ricono, C., Olivier, D., Mauger, S., Poinsot, D., Faloya, V., Daniel, L., & Cortesero, A. M. (2020). Oviposition Preference of the Cabbage Root Fly towards Some Chinese Cabbage Cultivars: A Search for Future Trap Crop Candidates. Insects, 11(2), 127. https://doi.org/10.3390/insects11020127