De Novo Transcriptomic Resources in the Brain of Vespa velutina for Invasion Control


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and RNA Extraction
2.2. cDNA Library Construction and Illumina Sequencing
2.3. De novo Assembly and Annotation
2.4. Detection of SSR, SNP Markers
2.5. Phylogenetic Analysis and Quantification of Gene Expression Levels
3. Results
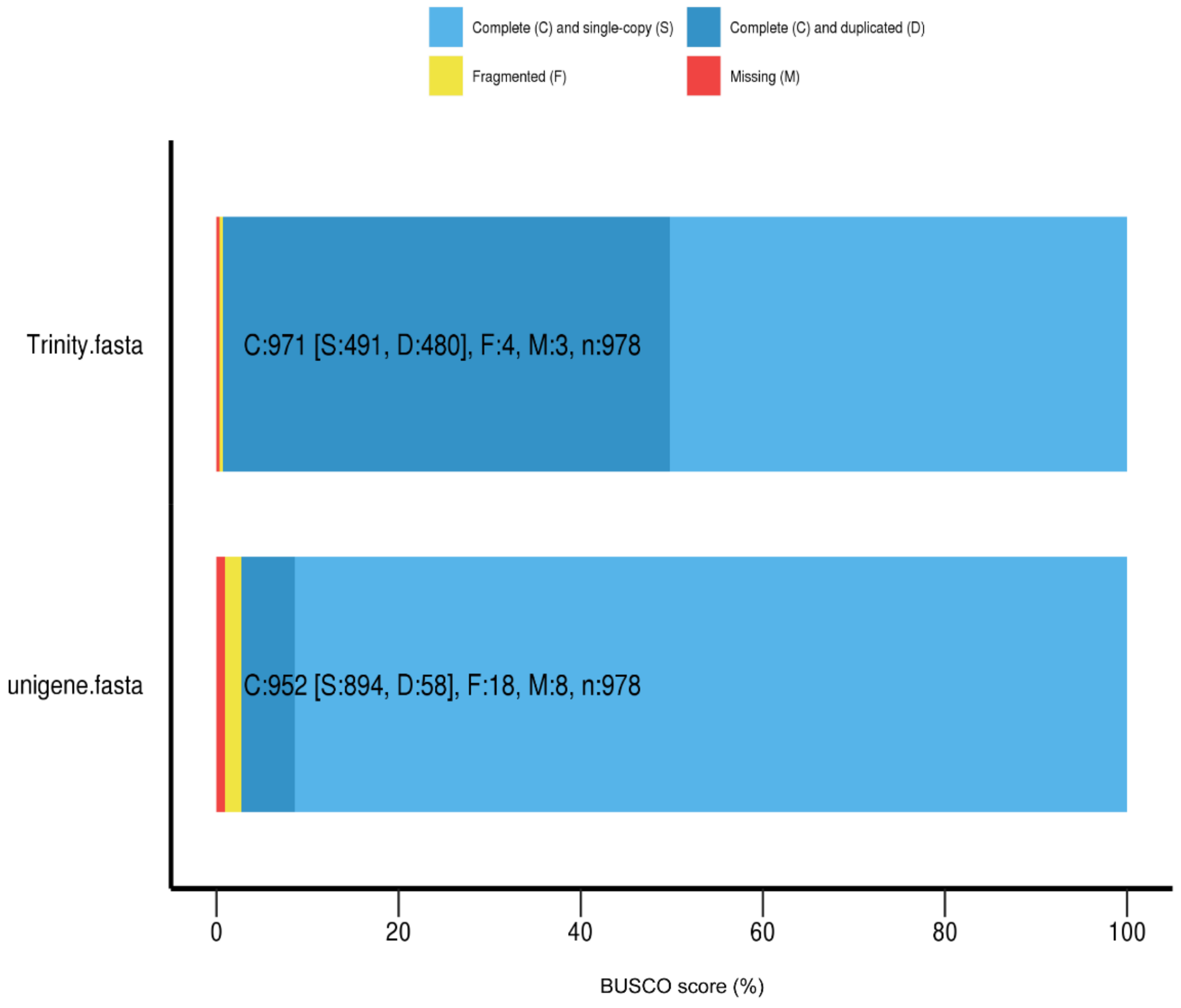
3.1. Sequencing and De Novo Assembly of Vespa Velutina Transcriptomes
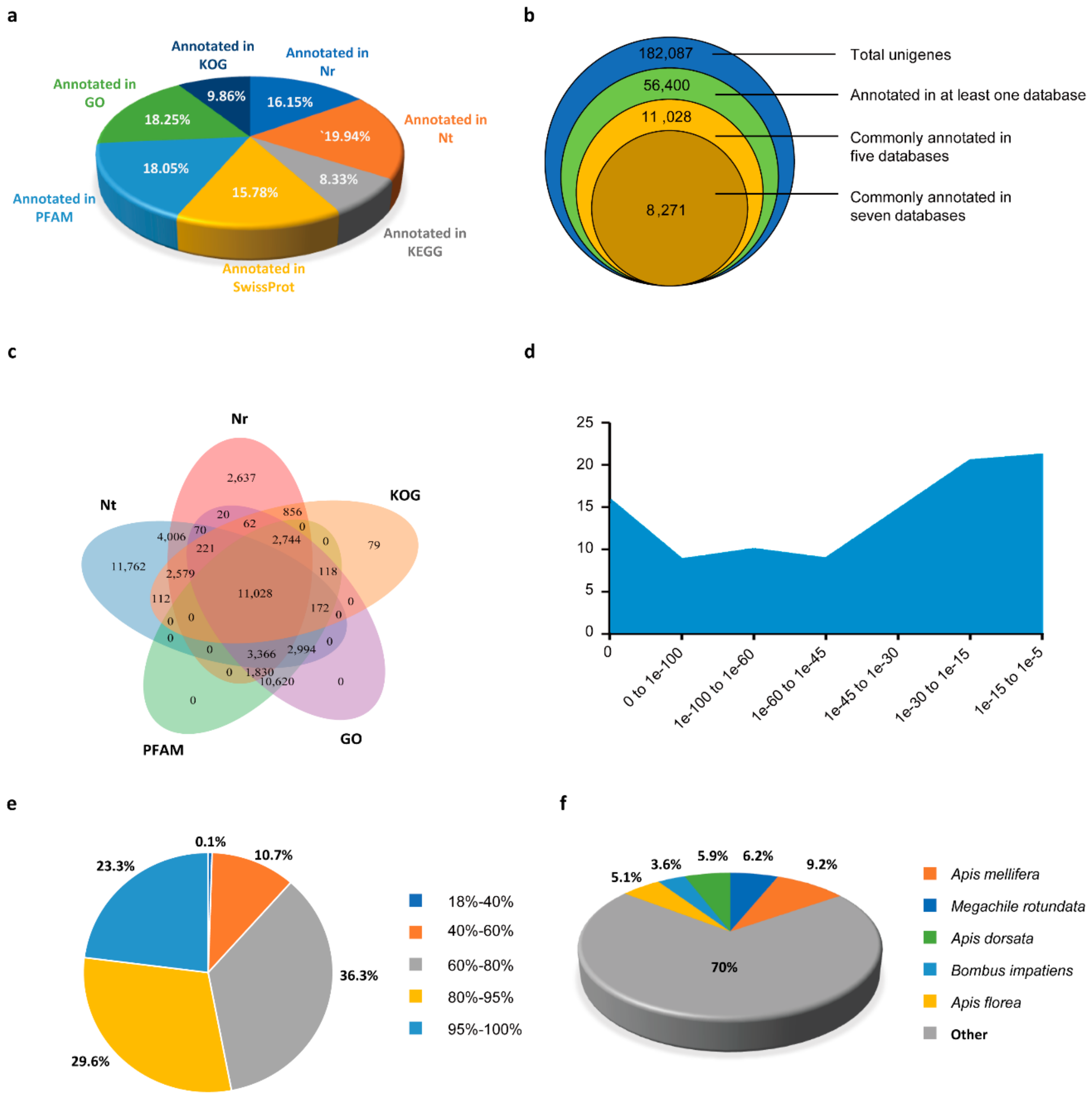
3.2. Functional Annotation
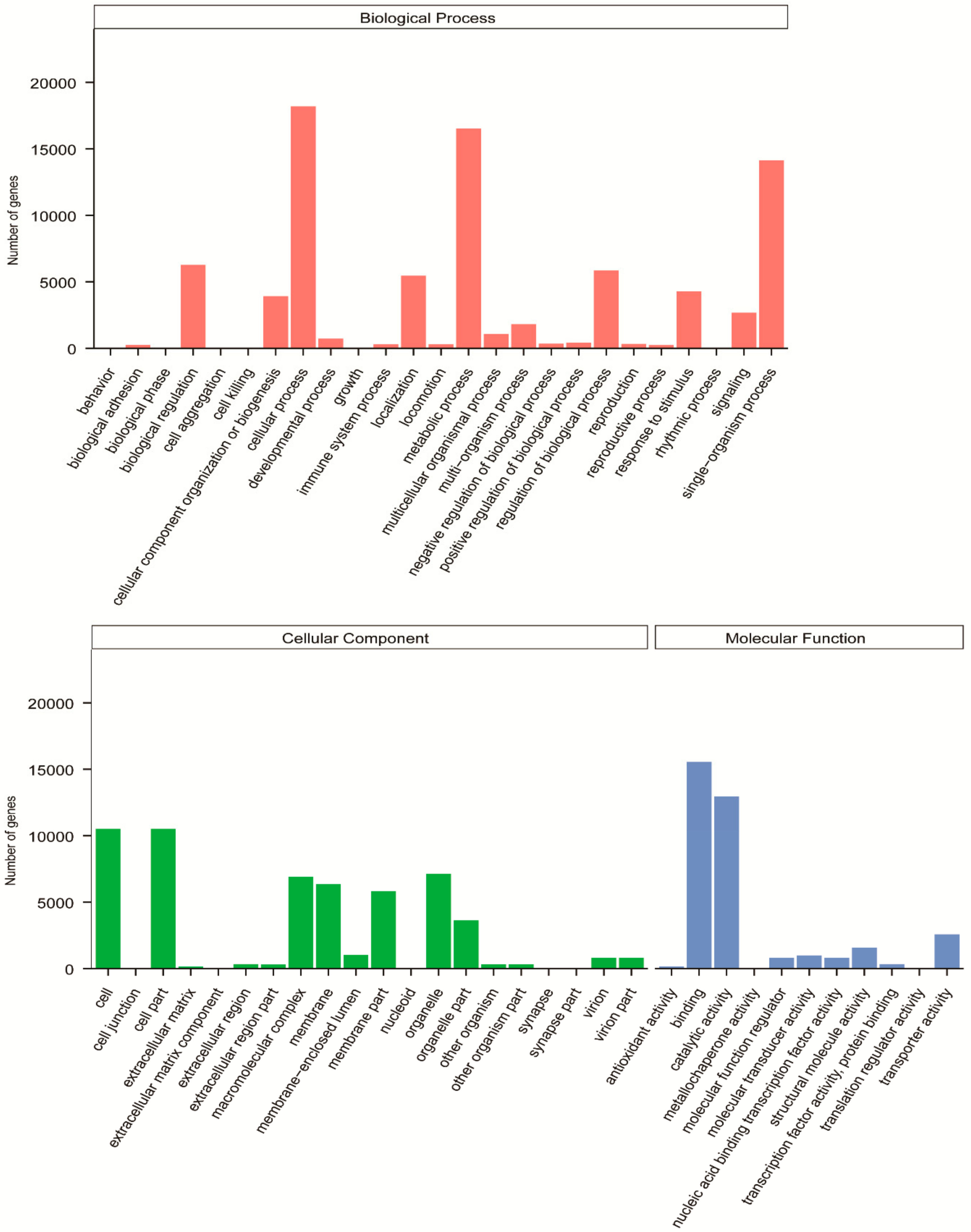
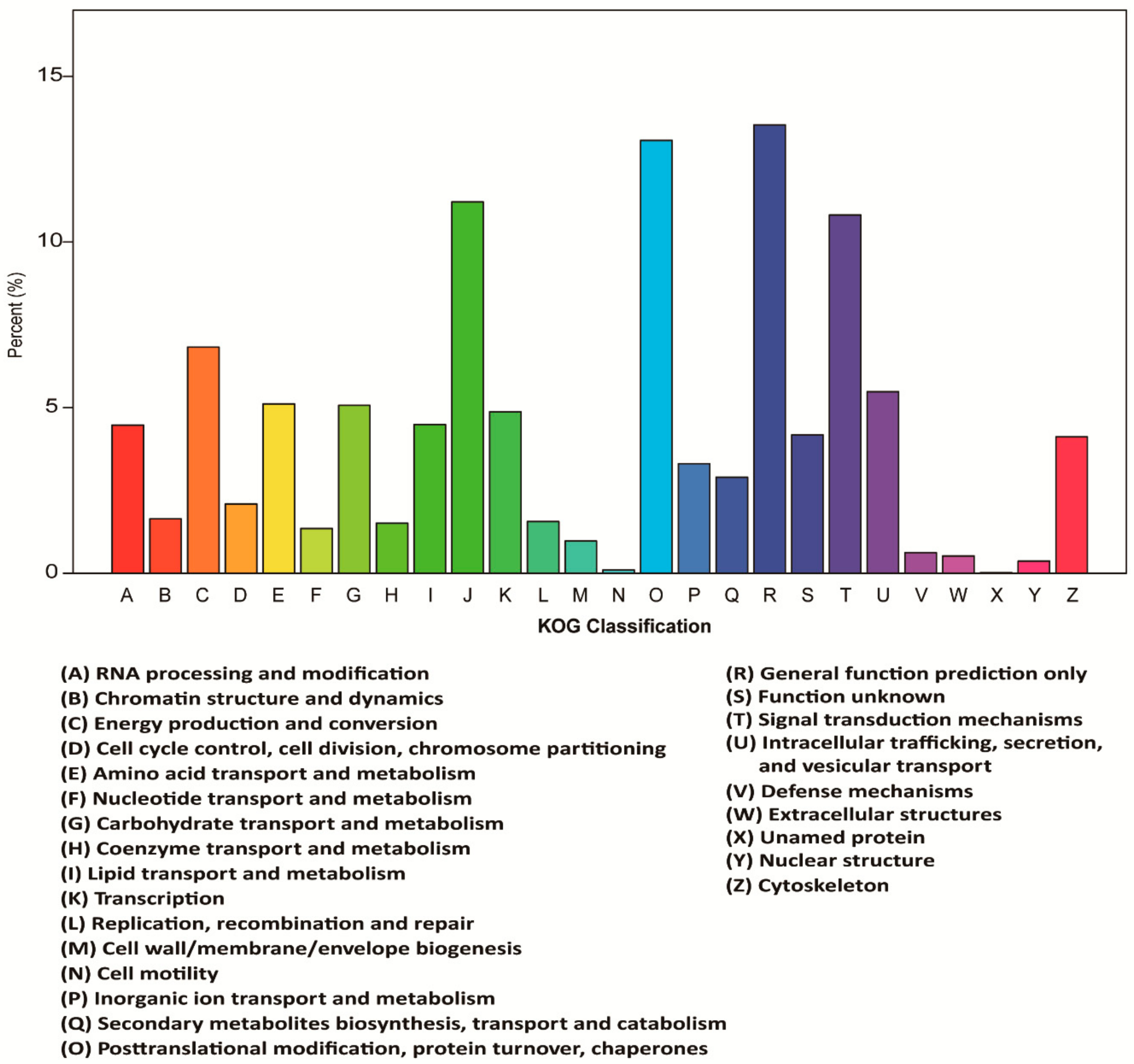
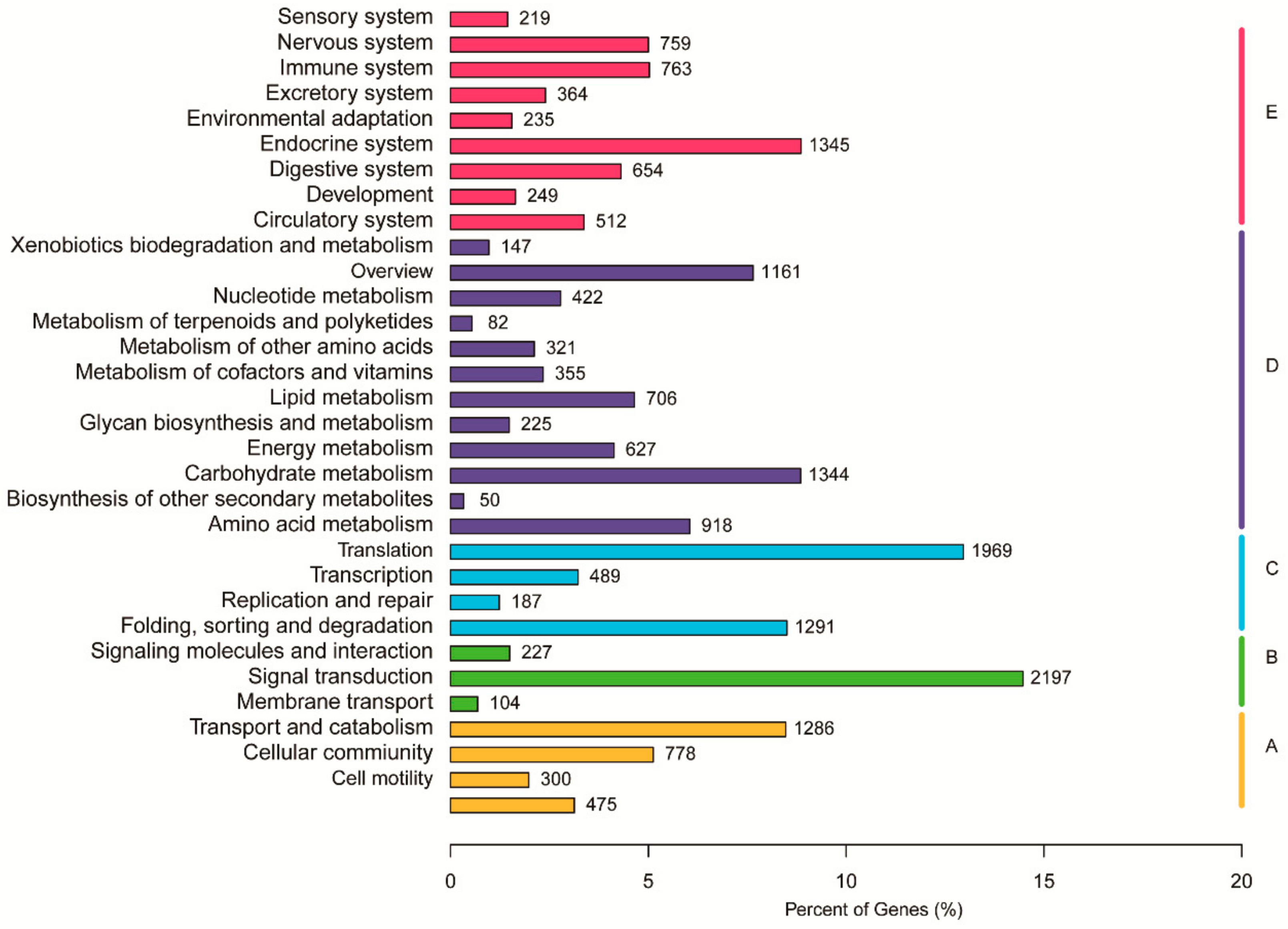
3.3. Functional Classification in GO, KOG, and KEGG Databases
3.4. Development of SSR and SNP Markers
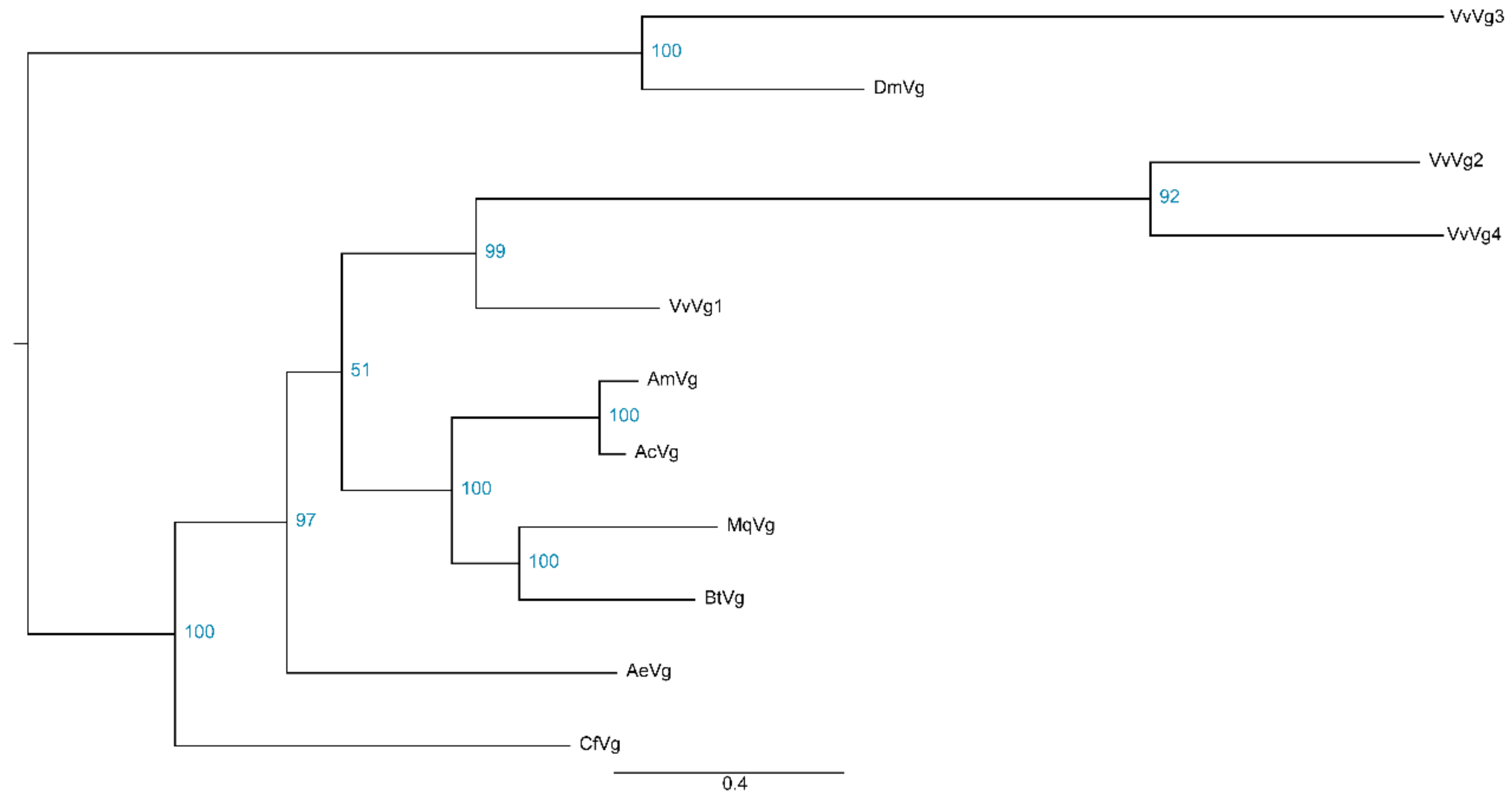
3.5. Identification of Potential Target Genes of OBP and Vitellogenin in the Hornet Brain
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Turchi, L.; Derijard, B. Options for the biological and physical control of Vespa velutina nigrithorax (Hym.: Vespidae) in Europe: A review. J. Appl. Entomol. 2018, 142, 553–562. [Google Scholar] [CrossRef]
- Rodriguez-Flores, M.S.; Seijo-Rodriguez, A.; Escuredo, O.; Seijo-Coello, M.D. Spreading of Vespa velutina in northwestern Spain: influence of elevation and meteorological factors and effect of bait trapping on target and non-target living organisms. J. Pest Sci. 2019, 92, 557–565. [Google Scholar] [CrossRef]
- Yamasaki, K.; Takahashi, R.; Harada, R.; Matsuo, Y.; Nakamura, M.; Takahashi, J.I. Reproductive interference by alien hornet Vespa velutina threatens the native populations of Vespa simillima in Japan. Naturwissenschaften 2019, 106, 15. [Google Scholar] [CrossRef] [PubMed]
- Poidatz, J.; Bressac, C.; Bonnard, O.; Thiery, D. Comparison of reproductive traits of foundresses in a native and an invasive hornet in Europe. J. Insect Physiol. 2018, 109, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.O.; Kim, C.S.; Lee, Y.S.; Choi, M.B. Abundance of diet-derived polychlorinated dibenzo-p-dioxins and polychlorinated dibenzofurans in the bodies and nests of the yellow-legged hornet Vespa velutina nigrithorax and risks to human health in South Korea. Sci. Total Environ. 2019, 654, 1033–1039. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, S.S.; Rico, M.R.L.; Arrondo, A.P.; Diaz, M.A.R.; Perez, B.V.; Vazquez, C.C. Vespa velutina and its growing relevance in our health area. Allergy 2018, 73, 188. [Google Scholar]
- Milanesio, D.; Saccani, M.; Maggiora, R.; Laurino, D.; Porporato, M. Recent upgrades of the harmonic radar for the tracking of the Asian yellow-legged hornet. Ecol. Evol. 2017, 7, 4599–4606. [Google Scholar] [CrossRef] [Green Version]
- Maggiora, R.; Saccani, M.; Milanesio, D.; Porporato, M. An Innovative Harmonic Radar to Track Flying Insects: the Case of Vespa velutina. Sci. Rep. 2019, 9, 11964. [Google Scholar] [CrossRef]
- Milanesio, D.; Saccani, M.; Maggiora, R.; Laurino, D.; Porporato, M. Design of an harmonic radar for the tracking of the Asian yellow-legged hornet. Ecol. Evol. 2016, 6, 2170–2178. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, P.J.; Ford, S.M.; Poidatz, J.; Thiery, D.; Osborne, J.L. Searching for nests of the invasive Asian hornet (Vespa velutina) using radio-telemetry. Commun. Biol. 2018, 1, 88. [Google Scholar] [CrossRef] [Green Version]
- Wen, P.; Cheng, Y.N.; Dong, S.H.; Wang, Z.W.; Tan, K.; Nieh, J.C. The sex pheromone of a globally invasive honey bee predator, the Asian eusocial hornet, Vespa velutina. Sci. Rep. 2017, 7, 12956. [Google Scholar] [CrossRef] [PubMed]
- Gevar, J.; Bagneres, A.G.; Christides, J.P.; Darrouzet, E. Chemical Heterogeneity in Inbred European Population of the Invasive Hornet Vespa velutina nigrithorax. J. Chem. Ecol. 2017, 43, 763–777. [Google Scholar] [CrossRef] [PubMed]
- Couto, A.; Monceau, K.; Bonnard, O.; Thiery, D.; Sandoz, J.C. Olfactory attraction of the hornet Vespa velutina to honeybee colony odors and pheromones. PLoS ONE 2014, 9, e115943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishi, S.; Goka, K. Review of the invasive yellow-legged hornet, Vespa velutina nigrithorax (Hymenoptera: Vespidae), in Japan and its possible chemical control. Appl. Entomol. Zool. 2017, 52, 361–368. [Google Scholar] [CrossRef]
- Sauvard, D.; Imbault, V.; Darrouzet, E. Flight capacities of yellow-legged hornet (Vespa velutina nigrithorax, Hymenoptera: Vespidae) workers from an invasive population in Europe. PLoS ONE 2018, 13, e0198597. [Google Scholar] [CrossRef] [Green Version]
- Poidatz, J.; Monceau, K.; Bonnard, O.; Thiery, D. Activity rhythm and action range of workers of the invasive hornet predator of honeybees Vespa velutina, measured by radio frequency identification tags. Ecol. Evol. 2018, 8, 7588–7598. [Google Scholar] [CrossRef] [Green Version]
- Poidatz, J.; Bressac, C.; Bonnard, O.; Thiery, D. Delayed sexual maturity in males of Vespa velutina. Insect Sci. 2018, 25, 679–689. [Google Scholar] [CrossRef]
- Gong, Z.; Tan, K.; Nieh, J.C. Hornets possess long-lasting olfactory memories. J. Exp. Biol. 2019, 222. [Google Scholar] [CrossRef] [Green Version]
- Couto, A.; Mitra, A.; Thiery, D.; Marion-Poll, F.; Sandoz, J.C. Hornets Have It: A Conserved Olfactory Subsystem for Social Recognition in Hymenoptera? Front. Neuroanat. 2017, 11, 48. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, R.; Okuyama, H.; Minoshima, Y.N.; Takahashi, J.I. Complete mitochondrial DNA sequence of the alien hornet Vespa velutina (Insecta: Hymenoptera) invading Kyushu Island, Japan. Mitochondrial DNA B 2018, 3, 179–181. [Google Scholar] [CrossRef]
- Takahashi, R.; Okuyama, H.; Kiyoshi, T.; Takahashi, J. Complete mitochondrial DNA sequence of the invasive hornet Vespa velutina (Insecta, Hymenoptera) found in Japan. Mitochondrial DNA B 2017, 2, 143–144. [Google Scholar] [CrossRef] [Green Version]
- Okuyama, H.; Martin, S.J.; Takahashi, J.I. Complete mitochondrial DNA sequence of the tropical hornet Vespa affinis (Insecta, Hymenoptera). Mitochondrial DNA B 2017, 2, 776–777. [Google Scholar] [CrossRef] [Green Version]
- Arca, M.; Capdevielle-Dulac, C.; Villemant, C.; Mougel, F.; Arnold, G.; Silvain, J.F. Development of microsatellite markers for the yellow-legged Asian hornet, Vespa velutina, a major threat for European bees. Conserv. Genet. Resour. 2012, 4, 283–286. [Google Scholar] [CrossRef]
- Stainton, K.; Hall, J.; Budge, G.E.; Boonham, N.; Hodgetts, J. Rapid molecular methods for in-field and laboratory identification of the yellow-legged Asian hornet (Vespa velutina nigrithorax). J. Appl. Entomol. 2018, 142, 610–616. [Google Scholar] [CrossRef]
- Granato, A.; Negrisolo, E.; Bonomi, J.; Zulian, L.; Cappa, F.; Bortolotti, L.; Mutinelli, F. Recent confirmation of a single haplotype in the Italian population of Vespa velutina. Biol. Invasions 2019, 21, 2811–2817. [Google Scholar] [CrossRef]
- Takahashi, J.; Okuyama, H.; Kiyoshi, T.; Takeuchi, T.; Martin, S.J. Origins of Vespa velutina hornets that recently invaded Iki Island, Japan and Jersey Island, UK. Mitochondrial DNA DNA Mapp. Seq. Anal. 2019, 30, 434–439. [Google Scholar] [CrossRef]
- Scott, J.G.; Michel, K.; Bartholomay, L.C.; Siegfried, B.D.; Hunter, W.B.; Smagghe, G.; Zhu, K.Y.; Douglas, A.E. Towards the elements of successful insect RNAi. J. Insect Physiol. 2013, 59, 1212–1221. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.W.; Zhu, X.Y.; Chao, Q.J.; Zhang, Y.J.; Yang, Y.X.; Wang, R.R.; Zhang, Y.; Xie, M.Z.; Ge, Y.T.; Wu, X.L.; et al. Chemosensory Gene Families in the Oligophagous Pear Pest Cacopsylla chinensis (Hemiptera: Psyllidae). Insects 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Munch, D.; Ihle, K.E.; Salmela, H.; Amdam, G.V. Vitellogenin in the honey bee brain: Atypical localization of a reproductive protein that promotes longevity. Exp. Gerontol. 2015, 71, 103–108. [Google Scholar] [CrossRef]
- Ihle, K.E.; Page, R.E.; Frederick, K.; Fondrk, M.K.; Amdam, G.V. Genotype effect on regulation of behaviour by vitellogenin supports reproductive origin of honeybee foraging bias. Anim. Behav. 2010, 79, 1001–1006. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Simao, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- BUSCO Software. Available online: http://busco.ezlab.org/ (accessed on 2 February 2020).
- MISA Software. Available online: http://pgrc.ipk-gatersleben.de/misa/misa.html (accessed on 2 February 2020).
- Primer3 Software. Available online: http://primer3.sourceforge.net/releases.php (accessed on 2 February 2020).
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FigTree v1.4.4 Software. Available online: https://github.com/rambaut/figtree (accessed on 2 February 2020).
- Joga, M.R.; Zotti, M.J.; Smagghe, G.; Christiaens, O. RNAi Efficiency, Systemic Properties, and Novel Delivery Methods for Pest Insect Control: What We Know So Far. Front. Physiol. 2016, 7, 553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behura, S.K. Molecular marker systems in insects: current trends and future avenues. Mol. Ecol. 2006, 15, 3087–3113. [Google Scholar] [CrossRef]
- Hurst, G.D.D.; Jiggins, F.M. Problems with mitochondrial DNA as a marker in population, phylogeographic and phylogenetic studies: the effects of inherited symbionts. Proc. Biol. Sci. 2005, 272, 1525–1534. [Google Scholar] [CrossRef]
- Mehta, B.; Daniel, R.; Phillips, C.; McNevin, D. Forensically relevant SNaPshot(A (R)) assays for human DNA SNP analysis: a review. Int. J. Legal. Med. 2017, 131, 21–37. [Google Scholar] [CrossRef]
- Henriques, D.; Parejo, M.; Vignal, A.; Wragg, D.; Wallberg, A.; Webster, M.T.; Pinto, M.A. Developing reduced SNP assays from whole-genome sequence data to estimate introgression in an organism with complex genetic patterns, the Iberian honeybee (Apis mellifera iberiensis). Evol. Appl. 2018, 11, 1270–1282. [Google Scholar] [CrossRef] [Green Version]
- Tarver, M.R.; Huang, Q.; de Guzman, L.; Rinderer, T.; Holloway, B.; Reese, J.; Weaver, D.; Evans, J.D. Transcriptomic and functional resources for the small hive beetle Aethina tumida, a worldwide parasite of honey bees. Genom. Data 2016, 9, 97–99. [Google Scholar] [CrossRef] [Green Version]
- Guirao-Rico, S.; Gonzalez, J. Evolutionary insights from large scale resequencing datasets in Drosophila melanogaster. Curr. Opin. Insect Sci. 2019, 31, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Choudhury, D.R.; Singh, A.K.; Kumar, S.; Srinivasan, K.; Tyagi, R.K.; Singh, N.K.; Singh, R. Comparison of SSR and SNP Markers in Estimation of Genetic Diversity and Population Structure of Indian Rice Varieties. PLoS ONE 2013, 8, e84136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismail, N.A.; Rafii, M.Y.; Mahmud, T.M.M.; Hanafi, M.M.; Miah, G. Molecular markers: a potential resource for ginger genetic diversity studies. Mol. Biol. Rep. 2016, 43, 1347–1358. [Google Scholar] [CrossRef] [PubMed]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Li, H.; Zheng, H.; Zhao, L.; Xue, X.; Wu, L. De Novo Transcriptomic Resources in the Brain of Vespa velutina for Invasion Control. Insects 2020, 11, 101. https://doi.org/10.3390/insects11020101
Wang M, Li H, Zheng H, Zhao L, Xue X, Wu L. De Novo Transcriptomic Resources in the Brain of Vespa velutina for Invasion Control. Insects. 2020; 11(2):101. https://doi.org/10.3390/insects11020101
Chicago/Turabian StyleWang, Miao, Hanyu Li, Huoqing Zheng, Liuwei Zhao, Xiaofeng Xue, and Liming Wu. 2020. "De Novo Transcriptomic Resources in the Brain of Vespa velutina for Invasion Control" Insects 11, no. 2: 101. https://doi.org/10.3390/insects11020101
APA StyleWang, M., Li, H., Zheng, H., Zhao, L., Xue, X., & Wu, L. (2020). De Novo Transcriptomic Resources in the Brain of Vespa velutina for Invasion Control. Insects, 11(2), 101. https://doi.org/10.3390/insects11020101