Disparity of Phoresy in Mesostigmatid Mites upon Their Specific Carrier Ips typographus (Coleoptera: Scolytinae)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Sampling and Laboratory Handling
2.3. Data Analyses
3. Results
3.1. Diversity of Mites and Zoocenological Pattern
3.2. Location of Phoretic Mites on the Beetles’ Body
3.3. Carrier Activity and Dynamics of Phoresy
3.3.1. Ips typographus Flight Activity
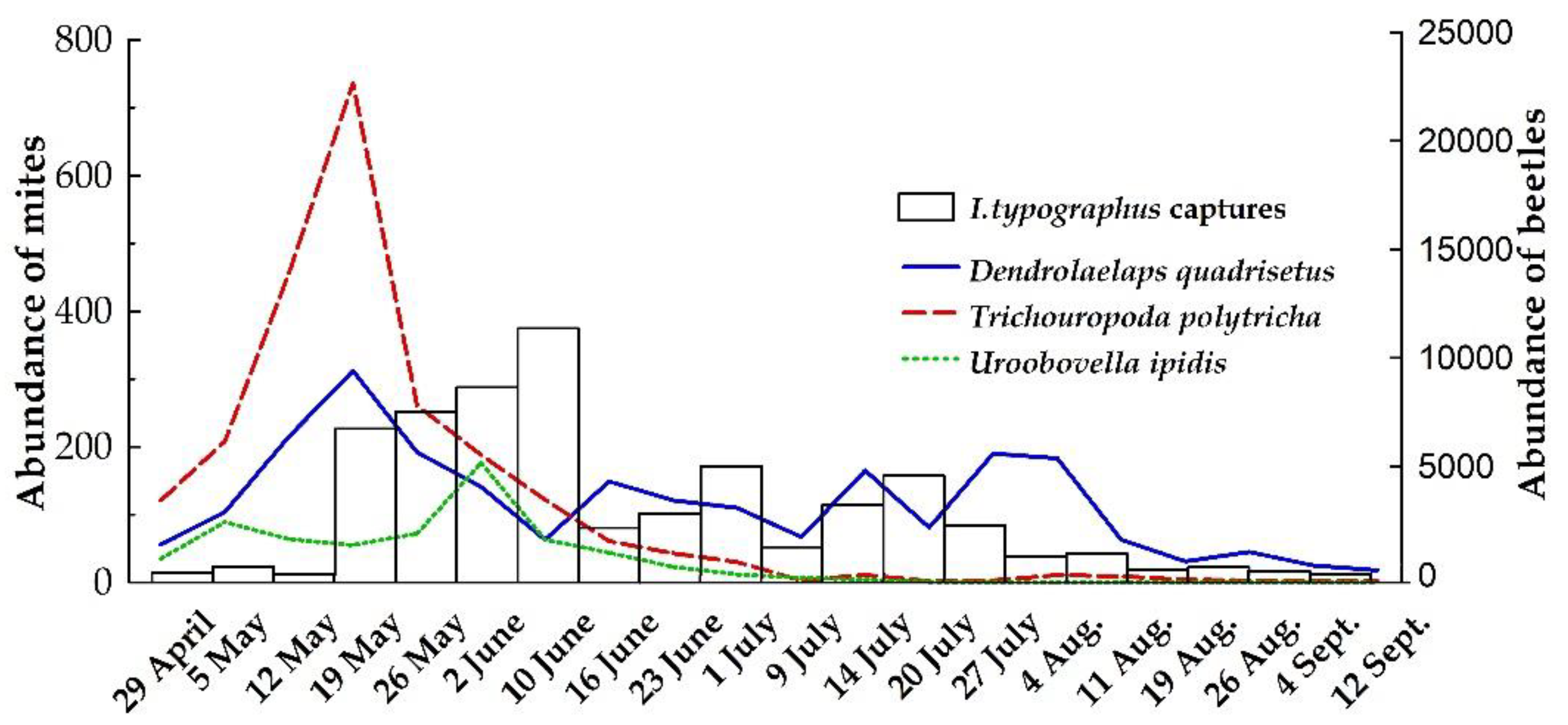
3.3.2. Dynamics of Phoresy
4. Discussion
4.1. Diversity and Zoocenological Pattern
4.2. Attachment Places
4.3. Dynamics of Phoresy
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Șofletea, N.; Curtu, L. Dendrologie, 2nd ed.; Pentru Viață Publishing House: Brasov, Romania, 2008; p. 418. ISBN 978-973-85874-4-1. [Google Scholar]
- Clinovschi, F. Dendrologie; Editura Universității Suceava: Suceava, Romania, 2005; p. 299. [Google Scholar]
- Caudullo, G.; Tinner, W.; De Rigo, D. Piceaabies in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publ. Off. EU: Luxembourg, 2016; p. e012300. [Google Scholar]
- Christiansen, E.; Bakke, A. The spruce bark beetle of Eurasia. In Dynamics of Forest Insect Populations; Berryman, A., Ed.; Plenum Publishing Corporation: New York, NY, USA, 1988; pp. 479–503. [Google Scholar]
- Simionescu, A.; Mihalciuc, V.; Chira, D.; Lupu, D.; Vlăduleasa, A.; Vişoiu, D.; Rang, C.; Mihai, D.; Mihalache, G.; Ciornei, C.; et al. Protecţia Pădurilor; Editura Muşatinii: Suceava, Romania, 2000; p. 867. [Google Scholar]
- Wermelinger, B. Ecology and management of the spruce bark beetle Ips typographus—A review of recent research. For. Ecol. Manag. 2004, 202, 67–82. [Google Scholar] [CrossRef]
- Mihalciuc, V.; Negură, A.; Cucos, V.; Cristoloveanu, G.; Cira, S. Utilizarea feromonilor sintetici în depistarea, prognoza și combaterea dăunătorilor din arboretele de rășinoase din România. Buc. For. 1995, 3, 33–43. [Google Scholar]
- Isaia, G.; Paraschiv, M. Research concerning the effect of synthetic pheromones on Pityogenes chalcographus L. in Brasov County. Bull. Trans. Brasov Series II 2011, 4, 55–60. [Google Scholar]
- Paraschiv, M.; Isaia, G.; Duduman, M.L. The effect of intercept® trap color on Ips typographus captures (preliminary results). Bull. Trans. Brasov Series II 2012, 5, 85–90. [Google Scholar]
- Isaia, G.; Paraschiv, M. Testing the ability of Ips typographus L. females to develop new maternal galleries in lab conditions. Bull. Trans. Brasov Series II 2015, 8, 23–30. [Google Scholar]
- Bentz, B.J.; Jönsson, A.M. Modeling Bark Beetle Responses to Climate Change. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Elsevier Inc.: Cambridge, MA, USA, 2015; pp. 533–553. [Google Scholar] [CrossRef]
- Ayres, M.P.; Wilkens, R.T.; Ruel, J.J.; Lombardero, M.J.; Erich, V. Nitrogen budgets of phloem-feeding bark beetles with and without symbiotic fungi. Ecology 2000, 81, 2198–2210. [Google Scholar] [CrossRef]
- Bleiker, K.; Six, D.L. Dietary benefits of fungal associates to an eruptive herbivore: Potential implications of multiple associates on host population dynamics. Environ. Entomol. 2007, 36, 1384–1396. [Google Scholar] [CrossRef] [PubMed]
- Viiri, H.; Lieutier, F. Ophiostomatoid fungi associated with the spruce bark beetle, Ips typographus, in three areas in France. Ann. For. Sci. 2004, 61, 215–219. [Google Scholar] [CrossRef] [Green Version]
- Kršiak, B.; Zach, P.; Kulfan, J. The Role of Hylastes Cunicularius Erichson (Coleoptera: Scolytidae) in Transferring Uropodine Mites in a Mountain Spruce Forest. J. For. Sci. 2010, 56, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Moser, J.C. Use of Sporothecae by Phoretic Tarsonemus Mites to Transport Ascospores of Coniferous Bluestain Fungi. Trans. Br. Mycol. Soc. 1985, 84, 750–753. [Google Scholar] [CrossRef]
- O’Connor, B.M. Acarine-Fungal Relationships: The Evolution of Symbiotic Associations. In Fungus-Insect Relationships: Perspectives in Ecology and Evolution; Wheeler, Q., Blackwell, M., Eds.; Columbia University Press: New York, NY, USA, 1984; pp. 354–381. [Google Scholar]
- Klepzig, K.D.; Moser, J.C.; Lombardero, M.J.; Ayres, M.P.; Hofstetter, R.W.; Walkinshaw’, C.J. Mutualism and Antagonism: Ecological Interactions among Bark Beetles, Mites and Fungi. In Biotic Interactions in Plant-Pathogen Associations; Jeger, M.J., Spence, N.J., Eds.; CABI Publishing: Wallingford, UK, 2001; pp. 237–267. [Google Scholar]
- Camerik, A.M. Phoresy revisited. In Trends in Acarology; Sabelis, M., Bruin, J., Eds.; Springer: Dordrecht, The Nederlands, 2010; pp. 333–336. ISBN 978-90-481-9837-5. [Google Scholar]
- Pérez-Martínez, S.; Moraza, M.L.; Saloña-Bordas, M.I. Gamasina Mites (Acari: Mesostigmata) Associated with Animal Remains in the Mediterranean Region of Navarra (Northern Spain). Insects 2019, 10, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houck, M.A.; O’Connor, B.M. Ecological and evolutionary significance of phoresy in the Astigmata. Annu. Rev. Entomol. 1991, 36, 611–636. [Google Scholar] [CrossRef]
- Macchioni, F. Importance of Phoresy in the Transmission of Acarina. Parassitologia 2007, 49, 17–22. [Google Scholar]
- Kinn, D.N.; Witcosky, J.J. Variation in southern pine beetle attack height associated with phoretic uropodid mites. Can. Entomol. 1978, 110, 249–251. [Google Scholar] [CrossRef]
- Bajerlein, D.; Błoszyk, J. Phoresy of Uropoda orbicularis (Acari: Mesostigmata) by beetles (Coleoptera) associated with cattle dung in Poland. Eur. J. Entomol. 2004, 101, 185–188. [Google Scholar] [CrossRef]
- Stephen, F.M.; Berisford, C.W.; Dahlsten, D.L.; Fenn, P.; Moser, J.C. Invertebrate and microbial associates. In Beetle-Pathogen Interactions in Conifer Forests; Schowalter, T.D., Filip, G.M., Eds.; Academic Press: San Diego, CA, USA, 1993; pp. 129–153. [Google Scholar]
- Lindquist, E.E. Mites and Regulations of Bark Beetle Populations. In Proceedings of the 2nd International Congress of Acarology, Sutton Bonington, UK, 19–25 July 1967; Evans, G.O., Ed.; Section VIII, Biological Control. Publishing House of the Hungarian Academy of Sciences Akademiai Kiado: Budapest, Hungary, 1969; pp. 389–399. [Google Scholar]
- Maslov, A.D. Mites Associated with Ips typographus, L. in Russia. In Proceedings of the IOBC/EPRS Conference Biological Methods in Integrated Plant Protection and Products, Poznan, Poland, 15–19 May 2006; pp. 15–19. [Google Scholar]
- Moser, J.C.; Kiełczewski, B.; Wiśniewski, J.; Bałazy, S. Evaluating Pyemotes Dryas (Vitzthum 1923) (Acari: Pyemotidae) as a Parasite of the Southern Pine Beetle. Int. J. Acaro. 1978, 4, 67–70. [Google Scholar] [CrossRef]
- Hofstetter, R.W.; Moser, J.C.; Blomquist, S.R. Mites associated with bark beetles and their hyperphoretic ophiostomatoid fungi. In Ophiostomatoid Fungi: Expanding Frontiers; Seifert, K.A., de Beer, Z.W., Wingfield, M.J., Eds.; Publisher CBS Biodiversity: Utrecht, The Netherlands, 2013; Series 12; pp. 165–176. [Google Scholar]
- Vissa, S.; Hofstetter, R.W. The role of mites in bark and ambrosia beetle-fungal interactions. In Insect Physiology and Ecology; Shields, V., Ed.; InTech: Rijeka, Croatia, 2017; pp. 135–156. [Google Scholar]
- Hofstetter, R.W.; Cronin, J.T.; Klepzig, K.D.; Moser, J.C. Antagonisms, mutualisms and commensalisms affect outbreak dynamics of the southern pine beetle. Oecologia 2006, 147, 679–691. [Google Scholar] [CrossRef]
- Hofstetter, R.W.; Dinkins-Bookwalter, J.; Davies, T.S.; Klepzig, K.D. Symbiotic associations of bark beetles. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 209–245. [Google Scholar]
- Lombardero, M.J.; Ayres, M.P.; Hofstetter, R.W.; Moser, J.C.; Lepzig, K.D. Strong Indirect Interactions of Tarsonemus Mites (Acarina: Tarsonemidae) and Dendroctonus Frontalis (Coleoptera: Scolytidae). Oikos 2003, 102, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Walter, D.E.; Behan-Pelletier, V. Mites in Forest Canopies: Filling the Size Distribution Shortfall? Ann. Rev. Entomol. 1999, 44, 1–19. Available online: http://www.ncbi.nlm.nih.gov/pubmed/15012369 (accessed on 12 September 2020). [CrossRef]
- Zhang, Z.Q. Animal biodiversity: An introduction to higher-level classification and taxonomic richness. Zootaxa 2011, 3148, 7–12. [Google Scholar] [CrossRef]
- Ochoa, R.; Erbe, E.F.; Pettis, J.S.; Wergin, W.P. Examination of frozen, hydrated mites using low temperature field emission scanning electron microscopy. Microsc. Microanal. 2000, 6, 874–875. [Google Scholar] [CrossRef]
- Wergin, W.P.; Ochoa, R.; Erbe, E.F.; Craemer, C.; Raina, A.K. Use of Low-Temperature Field Emission Scanning Electron Microscopy to Examine Mites. Scanning 2000, 22, 145–155. [Google Scholar] [CrossRef]
- Zhang, Z.Q. Accelerating studies on the ontogeny and morphological diversity in immature mites. Zootaxa 2018, 4540, 5–6. [Google Scholar] [CrossRef]
- Moser, J.C. Mites Associated with Forest Insects; For Willamette Institute for Biological Control. Inc.: Monroe, LA, USA, 1995; pp. 1–52. Available online: https://www.fs.fed.us/r6/icbemp/science/moser.pdf (accessed on 12 September 2020).
- Bajerlein, D.; Witaliński, W.; Adamski, Z. Morphological diversity of pedicels in phoretic deutonymphs of Uropodina mites (Acari: Mesostigmata). Arthropod Struct. Dev. 2013, 42, 185–196. [Google Scholar] [CrossRef]
- Behdran, A.K.; Ramani, N. Relationship between phoretic mites and their carrier, the banana pseudostem weevil Odoiporus longicollis Oliver (Coleoptera: Curculionidae). Int. J. Acarol. 2019, 45, 361–365. [Google Scholar] [CrossRef]
- Paraschiv, M.; Martinez-Ruiz, C.; Fernández, M.M. Dynamic associations between Ips sexdentatus (Coleoptera: Scolytinae) and its phoretic mites in a Pinus pinaster forest in northwest Spain. Exp. Appl. Acarol. 2018, 75, 369–381. [Google Scholar] [CrossRef]
- Linnakoski, R.; Mahilainen, S.; Harrington, A.; Vanhanen, H.; Eriksson, M.; Mehtätalo, L.; Pappinen, A.; Wingfield, M.J. Seasonal Succession of Fungi Associated with Ips typographus Beetles and Their Phoretic Mites in an Outbreak Region of Finland. PLoS ONE 2016, 11, e0155622. [Google Scholar] [CrossRef] [Green Version]
- Moser, J.C.; Konrad, H.; Kirisits, T.; Carta, L.K. Phoretic Mites and Nematode Associates of Scolytus Multistriatus and Scolytus Pygmaeus (Coleoptera: Scolytidae) in Austria. Agric. For. Entomol. 2005, 7, 169–177. [Google Scholar] [CrossRef]
- Moser, J.C.; Burjanadze, M.S.; Klimov, P.; Carta, L.K. Phoretic Mite and Nematode Associates of the Spruce Bark Beetle, Ips typographus (Coleptera: Scolytidae) in Georgia. 2009. Available online: http://www.srs.fs.usda.gov/idip/spb_ii/Scientist_Docs/Moser_et_al_WFIWC_2009_poster.pdf (accessed on 12 September 2020).
- Moser, J.C.; Eidmann, H.H.; Regnander, J.R. The mites associated with Ips typographus in Sweeden. Ann. Zool. Fenn. 1989, 55, 23–27. [Google Scholar]
- Penttinen, R.; Viiri, H.; Moser, J.C. The Mites (Acari) Associated with Bark Beetles in the Koli National Park in Finland. Acarologia 2013, 53, 3–15. [Google Scholar] [CrossRef]
- Schäffer, S.; Kerschbaumer, M. Borkenkäfer und ihr Zusammenleben mit Hornmilben (Acari, Oribatida)—Bark beetles and their associations with oribatid mites (Acari, Oribatida). Forstsch. Aktuell. 2017, 62, 13–18. [Google Scholar]
- Moser, J.C.; Bogenschűtz, H. A Key to the Mites Associated with Flying Ips typographus in South Germany. Z. Angew. Entomol. 1984, 97, 437–450. [Google Scholar] [CrossRef]
- Gwiazdowicz, D.J.; Kamczyc, J.; Bloszyk, J. The diversity of phoretic Mesostigmata on Ips typographus (Coleoptera: Scolytinae) caught in the Karkonosze forest. Eur. J. Entomol. 2011, 108, 489–491. [Google Scholar] [CrossRef] [Green Version]
- Fernández, M.; Diez, J.; Moraza, M.L. Acarofauna associated with Ips sexdentatus in northwest Spain. Scand. J. For. Res. 2013, 28, 358–362. [Google Scholar] [CrossRef]
- Moraza, M.L.; Bueno, A.; Diez, J.J.; Fernández, M.M. Short note: First European record of the family Cercomegistidae Trägardh, 1937 (Acari:Mesostigmata) in Spain. Int. J. Acarol. 2008, 34, 217–218. [Google Scholar] [CrossRef]
- Moraza, M.L.; Fernández, M.M.; Jurc, M. Phoretic mites of the six-spined engraver beetle, Ips sexdentatus (Böerner, 1776) (Coleoptera, Scolytinae), on Pinus halepensis in Slovenia. Int. J. Acarol. 2013, 39, 577–579. [Google Scholar] [CrossRef]
- Pernek, M.; Hrasovec, B.; Matosevic, D.; Pilas, I.; Kirisits, T.; Moser, J.C. Phoretic mites of three bark beetles (Pityokteines spp.) on Silver fir. J. Pest Sci. 2008, 81, 35–42. [Google Scholar] [CrossRef]
- Takov, D.; Pilarska, D.; Moser, J. Phoretic mites associated with spruce bark beetle Ips typographus L. (Curculionidae: Scolytinae) from Bulgaria. Acta Zool. Bulg. 2009, 61, 293–296. [Google Scholar]
- Čejka, M.; Holuša, J. Phoretic mites in uni- and bivoltine populations of Ips typographus: A 1-year case study. Turk. J. Zool. 2014, 38, 569–574. [Google Scholar] [CrossRef]
- Čejka, M.; Holuša, J. Phoretic mites (Mesostigmata) on double-spined spruce bark beetle Ips duplicatus (Coleoptera: Curculionidae: Scolytinae) in recent outbreak area in the central Europe. For. J. 2014, 60, 240–243. [Google Scholar] [CrossRef]
- Poliță, D.; Manu, M.; Marcu, V.M. Relationship among phoretic mites and Norway spruce bark beetles—Ips typographus and Pityogenes chalcographus. Rev. Păd. 2016, 131, 57–65. [Google Scholar]
- Manu, M.; Poliză, D.; Onete, M. Comparative analysis of the phoretic mites communities (Acari: Mesostigmata) associated with Ips typographus from natural and planted Norway spruce stands-Romania. Rom. Biotech. Lett. 2018, 23, 39–46. [Google Scholar] [CrossRef]
- Duduman, M.L. Răspunsul Adulților de Ips typographus L. și Hylobius abietis L. la Diferite Combinații de Stimuli Olfactivi Sintetici; Editura Universității Ștefan cel Mare: Suceava, Romania, 2019; p. 151. [Google Scholar]
- Blaženec, M.; Jakuš, R. Effect of (+)-limonene and 1-methoxy-2-propanol on Ips typographus response to pheromone blends. J. For. Res. 2009, 20, 37–44. [Google Scholar] [CrossRef]
- Schlyter, F.; Cederholm, I. Separation of the sexes of living spruce bark beetles, Ips typographus (L.), (Coleoptera: Scolytidae). Z. Angew. Entomol. 1981, 92, 42–47. [Google Scholar] [CrossRef]
- Bednarz, B.; Kacprzyk, M. An Innovative Method for Sex Determination of the European Spruce Bark Beetle Ips typographus (Coleoptera: Scolytinae). Entomol. Gen. 2012, 34, 111–118. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication. Urbana; University of Illinois Press: Champaign, IL, USA, 1963; p. 117. [Google Scholar]
- Kielczewski, B.; Moser, J.C.; Wisniewski, J. Surveying the Acarofauna Associated with Polish Scolytidae. Bull. Soc. Amis Sci. Lett. Poz. Serie D. 1983, 22, 151–159. [Google Scholar]
- Khaustov, A.A.; Klimov, P.B.; Trach, A.V.; Bobylev, A.N.; Salavatulin, V.M.; Khaustov, A.V.; Tolstikov, A.V. Review of mites (Acari) associatedwith the european spruce barkbeetle, Ips typographus (Coleoptera: Curculionidae: Scolytinae) in Asian Russia. Acarina 2018, 26, 137–151. [Google Scholar] [CrossRef]
- Kiełczewski, B.; Bałazy, S. Zagadnienie drapieżnictwa roztoczy (Acarina) na jajach korników (Scolytidae, Coleoptera) (The problem of predation of bark beetle eggs by mites—Translated from polish). Ekol. Polsk. 1966, 12, 161–163. [Google Scholar]
- Michalski, J.; Kaczmarek, S.; Ratajczak, E. Mites (Acari, Mesostigmata) in the Bark beetle Galleries (Coleoptera, Scolytida) of the Gorczanski National Park. Pol. J. Entomol. 1992, 61, 137–142. [Google Scholar]
- Khaustov, A.A.; Trach, V.A.; Bobylev, A.N. Mites (Acari) phoretic on six-toothed spruce bark beetle, Pityogenes chalcographus Linnaeus (Coleoptera: Curculionidae: Scolytinae), in western Siberia, Russia. Acarina 2016, 24, 137–151. [Google Scholar] [CrossRef] [Green Version]
- Pernek, M.; Wirth, S.; Blomquist, S.R.; Avtzis, D.N.; Moser, J.C. New Associations of Phoretic Mites on Pityokteines Curvidens (Coleoptera, Curculionidae, Scolytinae). Cent. Eur. J. Biol. 2012, 7, 63–68. [Google Scholar] [CrossRef]
- Pfammatter, J.A.; Moser, J.C.; Raffa, K.F. Mites Phoretic on Ips pini (Coleoptera: Curculionidae: Scolytinae) in Wisconsin Red Pine Stands. Ann. Entomol. Soc. Am. 2013, 106, 204–213. [Google Scholar] [CrossRef]
- Kielczewski, B.; Wisniewski, J. Bark Beetle Acarofauna in Different Types of Forest Habitat. Part III. Tarsonemini, Prostigmata, Acaridiae. Bull. Soc. Amis Sci. Lett. Poz. 1980, 20, 161–175. [Google Scholar]
- Moser, J.C.; Parry, T.J.; Furuta, K. Phoretic mites and their hyperphoretic fungi associated with flying Ips typographus japonicus Niijima (Col., Scolytidae) in Japan. J. Appl. Ent. 1997, 121, 425–428. [Google Scholar] [CrossRef]
- Wirth, S.F.; Weis, O.; Pernek, M. Comparison of phoretic mites associated with bark beetles Ips typographus and Ips cembrae from central Croatia. Sumar. List. 2016, 140, 549–560. [Google Scholar] [CrossRef]
- Turk, E.; Turk, F. Systematik und Okologie der Tyroglyphiden Mitteleuropas. In Beitrage zur Systematikund Okologie Mitteleuropaischer Acarina; Stammer, H.J., Ed.; Akademische Verlagsgesllschaft, Geest & Portig K-G: Leipzig, Germany, 1957; pp. 1–123. [Google Scholar]
- Bajerlein, D.; Witaliński, W. Localization and Density of Phoretic Deutonymphs of the Mite Uropoda Orbicularis (Parasitiformes: Mesostigmata) on Aphodius Beetles (Aphodiidae) Affect Pedicel Length. Naturwissenschaften 2014, 101, 265–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajerlein, D. Seasonal abundance and infestation of deutonymphs of Uropoda orbicularis (Muller, 1776) (Acari:Mesostigmata) phoretic on coprophilous beetles (Scarabeidae, Geotrupidae, Aphodiidae, Hydrophilidae, Histeridae). Int. J. Acarol. 2011, 37, 216–227. [Google Scholar] [CrossRef]
- Belozerov, V. Diapause and quiescence as two main kinds of dormancy and their significance in life cycles of mites and ticks (Chelicerata: Arachnida: Acari). Part 1. Acariformes. Acarina 2009, 16, 79–130. [Google Scholar]
- Barton, P.S.; Weaver, H.J.; Manning, A.D. Contrasting diversity dynamics of phoretic mites and beetles associated with vertebrate carrion. Exp. Appl. Acarol. 2014, 63, 1–13. [Google Scholar] [CrossRef]
- Vázquez, G.H.P. Ecology of Mite Phoresy on Mountain Pine Beetles. Ph.D. Thesis, Univeristyof Calgary, Calgary, AB, Canada, 2018. [Google Scholar]
- Annila, E. Influence of temperature upon the development and voltinism of Ips typographus L. (Coleoptera, Scolytidae). Ann. Zool. Fennici. 1969, 6, 161–208. [Google Scholar]
- Faccoli, M. Winter mortality in sub-corticolous populations of Ips typographus (Coleoptera Scolytidae) and its parasitoids in the South-eastern Alps. J. Pest Sci. 2002, 75, 62–68. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species | Number of Phoretic Mites | Dominance (%) | Frequency a (%) |
|---|---|---|---|---|
| Digamasellidae | Dendrolaelaps quadrisetus | 1896 | 45.38 Eudominant | 84.18 Euconstant |
| Ascidae | Proctolaelaps fiseri | 59 | 1.41 Subresident | 12.55 Accesory |
| Trematuridae | Trychouropoda polytricha | 1652 | 39.54 Eudominant | 53.02 Euconstant |
| Histiomatidae | Histiostoma piceae | 14 | 0.34 Subresident | 6.51 Accessory |
| Urodinychidae | Uroobovella ipidis | 523 | 12.52 Dominant | 37.20 Constant |
| Urodinychidae | Uroobovella vinicolora | 34 | 0.81 Subresident | 3.72 Accidental |
| Total | 4178 | 100 | 19.45 |
| 9Species | Developmental Stage | Under Elytra | Thorax | Elytral Declivity | Abdomen | Total |
|---|---|---|---|---|---|---|
| Dendrolaelaps quadrisetus | deutonymphs adults | 1878 | 13 | 5 | - | 1896 |
| Proctolaelaps fiseri | adults | 59 | - | - | - | 59 |
| Trychouropoda polytricha | deutonymphs | 5 | 880 | 743 | 24 | 1652 |
| Histiostoma piceae | deutonymphs * | 14 | - | - | - | 14 |
| Uroobovella ipidis | deutonymphs | - | 205 | 313 | 5 | 523 |
| Uroobovella vinicolora | deutonymphs | - | 18 | 16 | - | 34 |
| Total | 1956 | 1116 | 1077 | 29 | 4178 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paraschiv, M.; Isaia, G. Disparity of Phoresy in Mesostigmatid Mites upon Their Specific Carrier Ips typographus (Coleoptera: Scolytinae). Insects 2020, 11, 771. https://doi.org/10.3390/insects11110771
Paraschiv M, Isaia G. Disparity of Phoresy in Mesostigmatid Mites upon Their Specific Carrier Ips typographus (Coleoptera: Scolytinae). Insects. 2020; 11(11):771. https://doi.org/10.3390/insects11110771
Chicago/Turabian StyleParaschiv, Marius, and Gabriela Isaia. 2020. "Disparity of Phoresy in Mesostigmatid Mites upon Their Specific Carrier Ips typographus (Coleoptera: Scolytinae)" Insects 11, no. 11: 771. https://doi.org/10.3390/insects11110771
APA StyleParaschiv, M., & Isaia, G. (2020). Disparity of Phoresy in Mesostigmatid Mites upon Their Specific Carrier Ips typographus (Coleoptera: Scolytinae). Insects, 11(11), 771. https://doi.org/10.3390/insects11110771