Comparative Morphology of the Mouthparts in Three Predatory Stink Bugs (Heteroptera: Asopinae) Reveals Feeding Specialization of Stylets and Sensilla


Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Insect Collecting
2.2. Samples for SEM
2.3. Image Processing and Morphometric Measurement
2.4. Terminology
3. Results
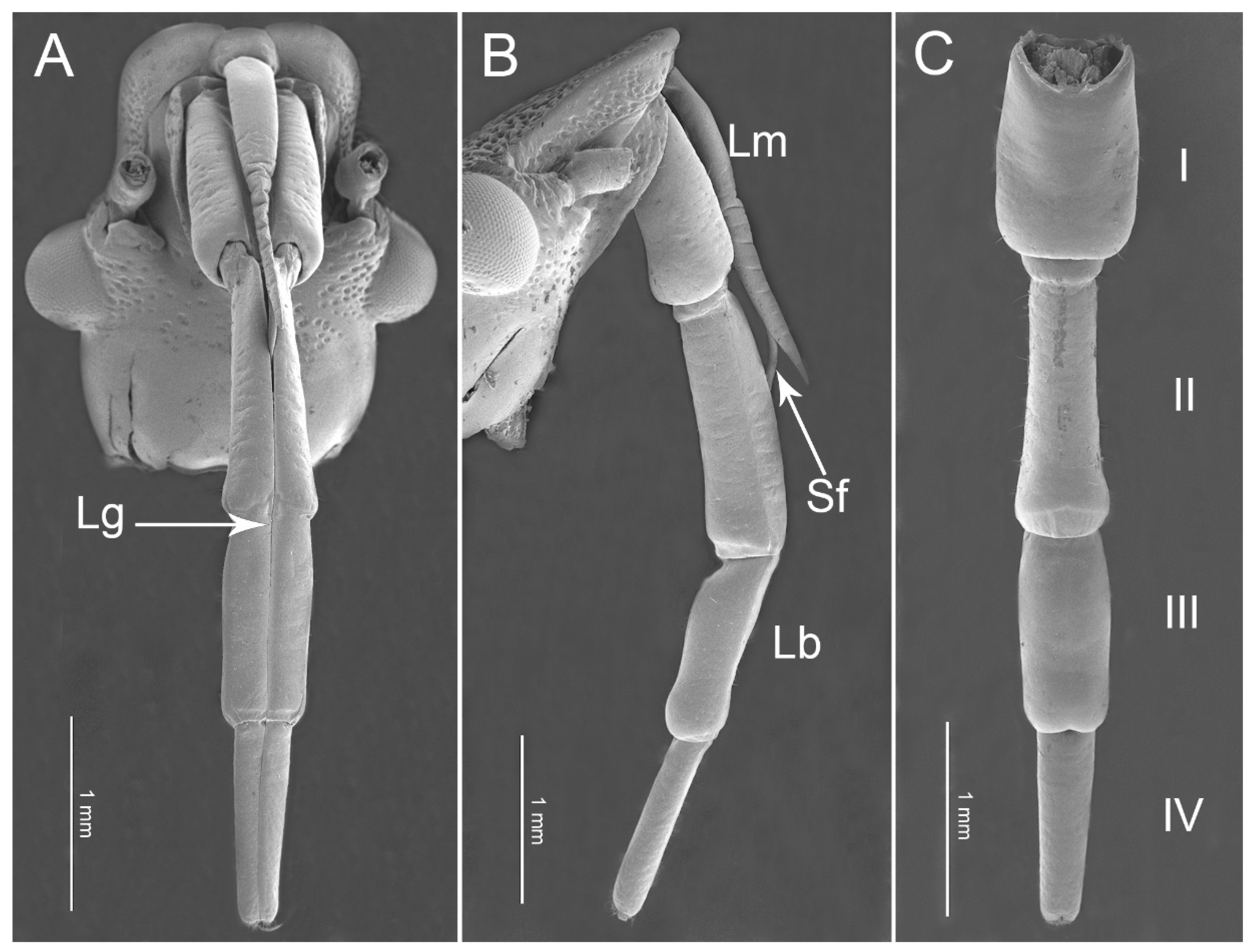
3.1. Overall Morphology of the Mouthparts
3.1.1. Labrum
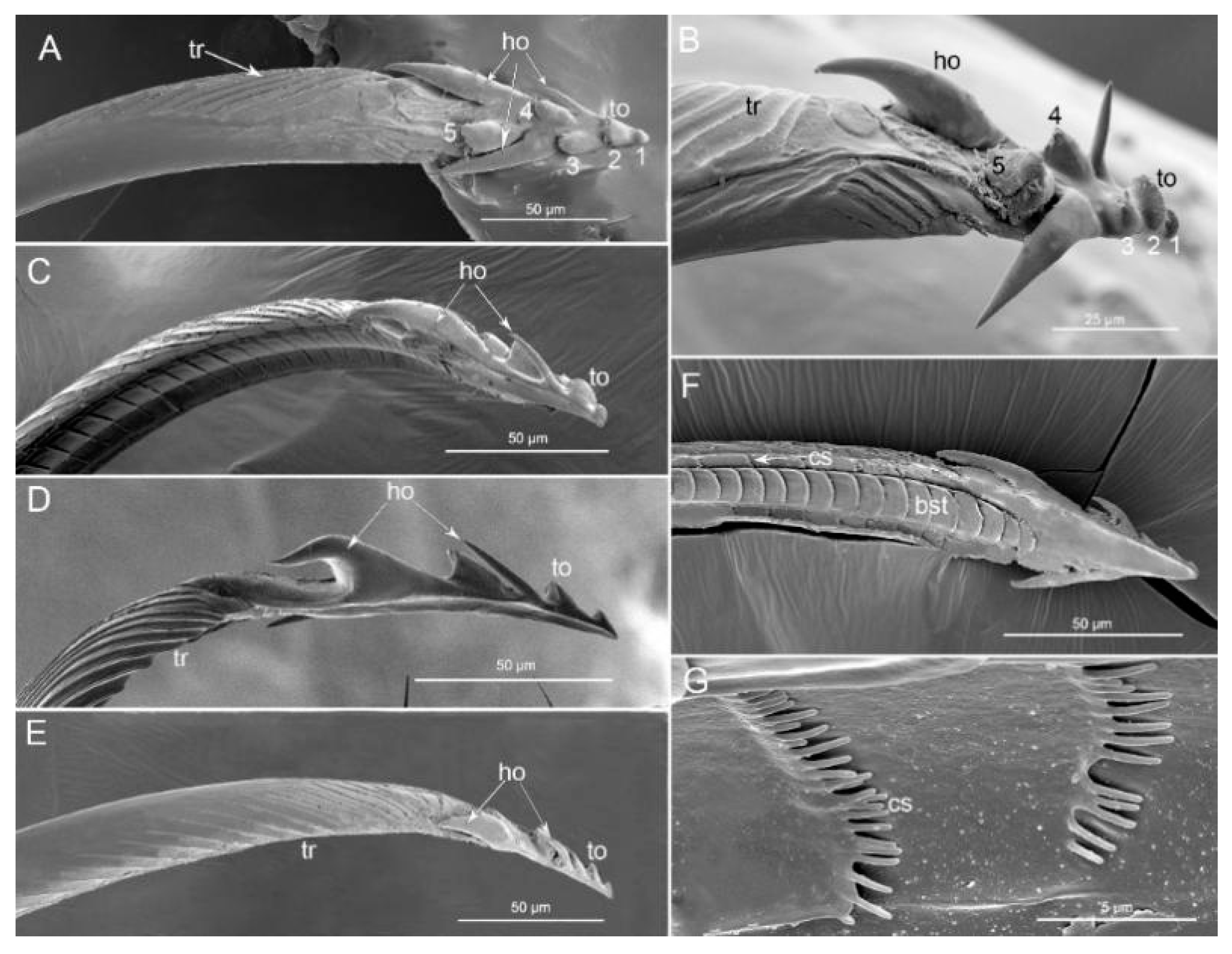
3.1.2. Labium
3.2. Stylet Fascicle
3.2.1. Mandibles
3.2.2. Maxillae
4. Discussion
4.1. The Mouthparts of Predatory Specialist
4.2. Sensilla Types
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cobben, R.H. Evolutionary Trends in Heteroptera. In Part II Mouthpart-Structures and Feeding Strategies; Mededelingen Landbouwhogeschool: Wageningen, The Netherlands, 1978; pp. 1–407. [Google Scholar]
- Faucheux, M.M. Relations entre l’ultrastructure des stylets manibulaires et maxillaires et la prise de nourriture chez les insects Hemipteres. CR Acad. Sci. Paris (Ser. D) 1975, 281, 41–44. [Google Scholar]
- Brożek, J. Phylogenetic signals from Nepomorpha (Insecta: Hemiptera: Heteroptera) mouthparts: Stylets bundle, sense organs, and labial segments. Sci. World J. 2014, 2014, 237854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, C.T. Structure and function in some external receptors. Symp. R. Entomol. Soc. Lond. 1970, 5, 59–76. [Google Scholar]
- Zacharuk, R.Y. Ultrastructure and function of insect chemosensilla. Annu. Rev. Entomol. 1980, 25, 27–47. [Google Scholar] [CrossRef]
- Romani, R.; Salerno, G.; Frati, F.; Conti, E.; Isidoro, N.; Bin, F. Oviposition behaviour in Lygus rugulipennis: A morpho-functional study. Entomol. Exp. Appl. 2005, 115, 17–25. [Google Scholar] [CrossRef]
- Rathore, Y.K. Studies on the mouthparts and feeding mechanism in Dysdercus cingulatus Fabr. (Pyrrhocoridae: Heteroptera). Indian J. Entomol. 1961, 23, 163–185. [Google Scholar]
- Cohen, A.C. Feeding adaptations of some predaceous Hemiptera. Ann. Entomol. Soc. Am. 1990, 83, 1215–1223. [Google Scholar] [CrossRef]
- Boyd, D.W. Digestive enzymes and stylet morphology of Deraeocoris nigritulus (Uhler) (Hemiptera: Miridae) reflect adaptations for predatory habits. Ann. Entomol. Soc. Am. 2003, 96, 667–671. [Google Scholar] [CrossRef] [Green Version]
- Roitberg, B.D.; Gillespie, D.R.; Quiring, D.M.J.; Alma, C.R.; Jenner, W.H.; Perry, J.; Peterson, J.H.; Salomon, M.; Van Laerhoven, S. The cost of being an omnivore: Mandible wear from plant feeding in a true bug. Naturwissenschaften 2005, 92, 431–434. [Google Scholar] [CrossRef]
- Bérenger, J.; Pluot-Sigwalt, D. Notes sur Micrauchenus lineola (Fabricius 1787), espècetermitophile et termitophage (Heteroptera: Reduviidae: Harpactorinae, Apiomerini). Ann. Soc. Entomol. Fr. 2009, 45, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Depieri, R.A.; Panizzi, A.R. Rostrum length, mandible serration, and food and salivary canal areas of selected species of stink bugs (Heteroptera, Pentatomidae). Rev. Bras. Entomol. 2010, 54, 584–587. [Google Scholar] [CrossRef] [Green Version]
- Depieri, R.A.; Siqueia, F.; Panizzi, A.R. Aging and food source effects on mandibular stylets teeth wear of Phytophagous Stink Bug (Heteroptera: Pentatomidae). Neotrop. Entomol. 2010, 39, 952–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.; Sahayaraj, K. Gross morphology and histology of head and salivary apparatus of the predatory bug, Rhynocoris marginatus. J. Insect Sci. 2012, 12, 19. [Google Scholar] [CrossRef] [Green Version]
- Brożek, J. A comparison of external and internal maxilla and mandible morphology of water bugs (Hemiptera: Heteroptera: Nepomorpha). Zootaxa 2013, 3635, 340–378. [Google Scholar] [CrossRef]
- Wang, Y.; Dai, W. Fine structure of mouthparts and feeding performance of Pyrrhocoris sibiricus Kuschakevich with remarks on the specialization of sensilla and stylets for seed feeding. PLoS ONE 2017, 12, e0177209. [Google Scholar] [CrossRef]
- Stubbins, F.L.; Mitchell, P.L.; Turnbull, M.W.; Reay-Jones, F.P.F.; Greene, J.K. Mouthpart morphology and feeding behavior of the invasive kudzu bug, Megacopta cribraria (Hemiptera: Plataspidae). Invertebr. Biol. 2017, 136, 309–320. [Google Scholar] [CrossRef]
- Wang, Y.; Li, L.F.; Dai, W. Fine morphology of the mouthparts in Cheilocapsus nigrescens (Hemiptera: Heteroptera: Miridae) reflects adaptation for phytophagous habits. Insects 2019, 10, 143. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Brożek, J.; Dai, W. Sensory armature and stylets of the mouthparts of Stephanitis nashi (Hemiptera: Cimicomorpha: Tingidae), their morphology and function. Micron 2020, 132, 102840. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Brożek, J.; Dai, W. Morphological disparity of the mouthparts in polyphagous species of Largidae (Heteroptera: Pentatomomorpha: Pyrrhocoroidea) reveals feeding specialization. Insects 2020, 11, 145. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, J.R.; Wang, W.S.; Brożek, J.; Dai, W. Unique fine morphology of mouthparts in Haematoloecha nigrorufa (Stål) (Hemiptera: Reduviidae) adapted to millipede feeding. Insects 2020, 11, 386. [Google Scholar] [CrossRef]
- Wang, Y.; Dai, W. How does the intricate mouthpart apparatus coordinate for feeding in the hemimetabolous insect pest Erthesina fullo? Insects 2020, 11, 503. [Google Scholar] [CrossRef]
- De Clercq, P. Predaceous stinkbugs (Pentatomidae: Asopinae). In Heteroptera of Economic Importance; Schaefer, C.W., Panizzi, A.R., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 737–789. [Google Scholar]
- Rani, P.U.; Madhavendra, S.S. Morphology and distribution of antennal sense organs and diversity of mouthpart structures in Odontopus nigricornis (Stål) and Nezara viridula L. (Hemiptera). Int. J. Insect Morphol. Embryol. 1995, 24, 119–132. [Google Scholar] [CrossRef]
- Rani, P.U.; Madhavendra, S.S. External morphology of antennal and rostral sensillae in four hemipteran insects and their possible role in host plant selection. Int. J. Trop. Insect Sci. 2005, 25, 198–207. [Google Scholar] [CrossRef]
- Rani, P.U. Sensillary morphology on the rostral apex and their possible role in prey location behaviour of the carnivorous stinkbug, Eocanthecona furcellata (Wolff) (Heteroptera: Pentatomidae). Acta. Zool. 2009, 90, 246–253. [Google Scholar] [CrossRef]
- Parveen, S.; Ahmad, A.; Brożek, J.; Ramamurthy, V.V. Morphological diversity of the labial sensilla of phytophagous and predatory Pentatomidae (Hemiptera: Heteroptera), with reference to their possible functions. Zootaxa 2015, 4039, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Barsagade, D.D.; Gathalkar, G.B. First predation record of Canthecona furcellata (Wolff.) (Hemiptera: Pentatomidae) on spinning stage silkworm Antheraea mylitta (Drury). Entomol. Res. 2016, 46, 236–245. [Google Scholar] [CrossRef]
- Rider, D.A.; Schwertner, C.F.; Vilímová, J.; Rédei, D.; Kment, P.; Thomas, D.B. Higher systematics of the Pentatomoidea. In Invasive Stink Bugs and Related Species (Pentatomoidea): Biology, Higher Systematics, Semiochemistry, and Management; McPherson, J.E., Ed.; CRC Press: Boca Raton, FL, USA, 2018; pp. 25–201. [Google Scholar]
- Schouteden, H. Genera Insectorum. Heteroptera. Fam. Pentatomidae. Subfam. Asopinae (Amyoteinae). Fasc 1906, 52, 1–82. [Google Scholar]
- Thomas, D.B. Taxonomic synopsis of the Old World asopine genera (Heteroptera: Pentatomidae). Insecta Mundi 1994, 8, 145–212. [Google Scholar]
- Rider, D. Pentatomidae. In Catalogue of the Heteroptera of the Palaearctic region, 5; Aukema, B., Rieger, C., Eds.; Netherlands Entomological Society: Amsterdam, The Netherlands, 2006; pp. 233–402. [Google Scholar]
- Gapud, V.P. A generic revision of the subfamily Asopinae, with consideration of its phylogenetic position in the family Pentatomidae and superfamily Pentatomoidea (Hemiptera-Heteroptera). Parts. I and II. Philipp. Entomol. 1991, 8, 865–961. [Google Scholar]
- Khloptseva, R.I. The use of entomophages in biological pest control in the USSR. Biocontrol News Inf. 1991, 12, 243–246. [Google Scholar]
- Hough-Goldstein, J.; Whalen, J. Inundative release of predatory stink bugs for control of Colorado potato beetle. Biol. Control 1993, 3, 343–347. [Google Scholar] [CrossRef]
- Zanuncio, J.C.; Alves, J.B.; Zanuncio, T.V.; Garcia, J.F. Hemipterous predators of eucalypt defoliator caterpillars. For. Ecol. Manag. 1994, 65, 65–73. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, G.; Bu, W.J. A review of the Chinese species of the genus Picromerus Amyot and Serville, with description of a new species (Hemiptera: Heteroptera: Pentatomidae: Asopinae). Zootaxa 2013, 3613, 146–164. [Google Scholar] [CrossRef] [PubMed]
- Roca-Cusachs, M.; Kim, J.; Park, J.; Jung, S. Taxonomic review of the predatory stink bugs of the Korean Peninsula (Heteroptera: Pentatomidae: Asopinae), with a key to the Korean species and a discussion of their usefulness as biological control agents. J. Asia-Pacific Entomol. 2020, 23, 113–123. [Google Scholar] [CrossRef]
- Gäbler, H. Picromerus bidens L. als Feind der Lophyruslarven. Tharandter Forstl. Jahrb. 1937, 88, 51–58. [Google Scholar]
- Gäbler, H. Die Bedeutung einiger Wanzenarten als Feinde der Nonne. Z. Angew. Entomol. 1938, 25, 277–290. [Google Scholar] [CrossRef]
- Engel, H. Populationsdynamik des Kieferspanners in verschiedenen Biotopen. Verh. VII Intern. Kongr. Entomol. 1939, 3, 1941–1949. [Google Scholar]
- Clausen, C.P. Entomophagous Insects; McGraw-Hill: New York, NY, USA, 1940; p. 688. [Google Scholar]
- Forsslund, K.H. Något om röda tallstekelns (Diprion sertifer Geoffr.) skadegörelse. Medd. Skogsförsöksanst 1946, 34, 365–390. [Google Scholar]
- Mayné, R.; Breny, R. Picromerus bidens L.: Morphologie. Biologie. Détermination de sa valeur d’utilisation dans la lute biologique contre le doryphore de la pomme de terre—La valeur économique antidoryphorique des Asopines indigènes belges. Parasitica 1948, 4, 189–224. [Google Scholar]
- Pschorn-Walcher, H.; Zinnert, K.D. Investigations on the ecology and natural control of the larch sawfly (Pristiphora erichsonii Htg, Hym: Tenthredinidae) in central Europe. Part II: Natural enemies: Their biology and ecology, and their role as mortality factors in P. erichsonii. Tech. Bull. Commonw. Inst. Biol. Control 1971, 14, 1–50. [Google Scholar]
- Mallach, N. Zur Kenntnis der kleinen Kiefern-Buschhornblattwespe, Diprion (Microdiprion) pallipes (Fall.) (Hym.: Diprionidae). Teil 3. Populationsökologie. Z. Angew. Entomol. 1974, 75, 337–380. [Google Scholar] [CrossRef]
- Asanova, R.B. New and rare species of bugs for the fauna of northern Kazakhstan (Heteroptera). Trudy Inst. Zool. Akad. Nauk Kazakhskoi SSR 1980, 39, 49–54. (In Russian) [Google Scholar]
- Javahery, M. Biology and ecology of Picromerus bidens (Hemiptera: Pentatomidae) in southeastern Canada. Entomol. News 1986, 97, 87–98. [Google Scholar]
- Larivière, M.-C.; Larochelle, A. Picromerus bidens (Heteroptera: Pentatomidae) in North America, with a world review of its distribution and bionomics. Entomol. News 1989, 100, 133–146. [Google Scholar]
- Lin, Y.J.; Long, J.; Zhang, S.M.; Lin, Z. A checklist of species of Asopinae (Hemiptera: Pentatomoidea) from China. Jiangxi Plant Protection 2000, 23, 36–39. (In Chinese) [Google Scholar]
- Rider, D.A.; Zheng, L.Y. Checklist and nomenclatural notes on the Chinese Pentatomidae (Heteroptera). III. Asopinae. Entomotaxonomia 2002, 24, 107–115. [Google Scholar]
- Tang, Y.T.; Guo, Y.; He, G.W.; Liu, C.X.; Chen, H.Y.; Zhang, L.S.; Wang, M.Q. Functional Responses of Picromerus lewisi Scott (Hemiptera: Pentatomidae) attacking Mythimna separata (Walker) (Lepidoptera: Noctuidae). Chin. J. Biol. Control 2018, 34, 825–830. (In Chinese) [Google Scholar]
- Tang, Y.T.; Wang, M.Q.; Chen, H.Y.; Wang, Y.; Zhang, H.M.; Chen, F.S.; Zhao, X.Q.; Zhang, L.S. Predation and behavior of Picromerus lewisi Scott to Spodoptera frugiperda (J. E. Smith). Chin. J. Biol. Con. 2019, 35, 698–703. [Google Scholar]
- Altner, H.; Prillinger, L. Ultrastructure of invertebrate chemo-, thermo-, and hygroreceptors and its functional significance. Int. Rev. Cytol. 1980, 67, 69–139. [Google Scholar]
- Frazier, J.L. Nervous system: Sensory system. In Fundamentals of Insect Physiology; Blum, M.S., Ed.; John Wiley & Sons: New York, NY, USA, 1985; pp. 287–356. [Google Scholar]
- Walker, A.A.; Weirauch, C.; Fry, B.G.; King, G.F. Venoms of Heteropteran Insects: A treasure trove of diverse pharmacological toolkits. Toxins 2016, 8, 43. [Google Scholar] [CrossRef] [Green Version]
- Grazia, J.; Panizzi, A.R.; Greve, C.; Schwertner, C.F.; Campos, L.A.; Garbelotto, T.; Fernandes, J.A.M. Stink Bugs (Pentatomidae). In True Bugs (Heteroptera) of the Neotropics; Chapter 22; Panizzi, A.R., Grazia, J., Eds.; Springer Science + Business Media: Dordrecht, The Netherlands, 2015; pp. 681–756. [Google Scholar]
- Gross, G.F. Plant-feeding and Other Bugs (Hemiptera) of South Australia. Heteroptera, Part I.; James, A.B., Ed.; Handbooks Committee of the South Australian Government: Adelaide, Australia, 1975.
- Stone, P.C. A Study of the External Morphology of a Predacious Stink-bug: Apateticus cynicus (Say); Family, Pentatomidae; Order, Hemiptera-Heteroptera. Master’s Thesis, University of Massachusetts Amherst, Amherst, MA, USA, 1936. [Google Scholar]
- Esquivel, J.F. Stink bug rostrum length vs. stylet penetration potential. Èntomol. Exp. Appl. 2019, 167, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Kim, J.G.; Jung, S.H. A New record of Genus Eocanthecona Bergroth (Hemiptera: Heteroptera: Pentatomidae: Asopinae) from the Korean Peninsula. Korean J. Appl. Entomol. 2015, 54, 257–261. [Google Scholar] [CrossRef]
- Schuh, R.T.; Slater, J.A. True Bugs of the World (Hemiptera: Heteroptera—Classification and Natural History; Cornell University Press: Ithaca, NY, USA, 1995. [Google Scholar]
- Esquivel, J.F. Estimating potential stylet penetration of southern green stink bug—A mathematical modeling approach. Èntomol. Exp. Appl. 2011, 140, 163–170. [Google Scholar] [CrossRef]
- Weiraucha, C.; Schuhb, R.T.; Cassisc, G.; Wheeler, W.C. Revisiting habitat and lifestyle transitions in Heteroptera (Insecta: Hemiptera): Insights from a combined morphological and molecular phylogeny. Cladistics 2018, 35, 67–105. [Google Scholar] [CrossRef] [Green Version]
- McGavin, G.C. Food and Feeding. In Bugs of the World; Chapter 6; Blandford: London, UK, 1993; pp. 139–158. [Google Scholar]
- Swart, C.C.; Felgenhauer, B.E. Structure and function of the mouthparts and salivary gland complex of the giant waterbug, Belostoma lutarium (Stål) (Hemiptera: Belostomatidae). Ann. Entomol. Soc. Am. 2003, 96, 870–882. [Google Scholar] [CrossRef]
- Sahayaraj, K.; Kanna, A.V.; Kumar, S.M. Gross morphology of feeding canal, salivary apparatus and digestive enzymes of salivary gland of Catamirus brevipennis (Serville ) (Hemiptera: Reduviidae). J. Entomol. Res. Soc. 2010, 12, 37–50. [Google Scholar]
- Brożek, J.; Zettel, H. A comparison of the external morphology and functions of labial tip sensilla in semiaquatic bugs (Hemiptera: Heteroptera: Gerromorpha). Eur. J. Entomol. 2014, 111, 275–297. [Google Scholar] [CrossRef] [Green Version]
- Taszakowski, A.; Nowińska, A.; Brożek, J. Morphological study of the labial sensilla in Nabidae (Hemiptera: Heteroptera: Cimicomorpha). Zoomorphology 2019, 138, 483–492. [Google Scholar] [CrossRef] [Green Version]
- Ando, T.; Sekine, S.; Inagaki, S.; Misaki, K.; Badel, L.; Moriya, H.; Sami, M.M.; Itakura, Y.; Chihara, T.; Kazama, H.; et al. Nanopore formation in the cuticle of an insect olfactory sensillum. Curr. Biol. 2019, 29, 1512–1520. [Google Scholar] [CrossRef] [PubMed]
- Slifer, E.H. The structure of arthropod chemoreceptors. Annu. Rev. Entomol. 1970, 15, 121–141. [Google Scholar] [CrossRef]
- Steinhrccht, R.A. Chemo-, hygro-, and thermoreceptors. In Biology of the Integument, Invertebrates; Berciter-Hahn, J., Matoltsy, A.G., Richards, K.S., Eds.; Springer: Berlin/Heidelberg, Germany, 1984; Volume 1, pp. 523–553. [Google Scholar]
- Catalá, S. Sensilla associated with the rostrum of eight species of Triatominae. J. Morphol. 1996, 228, 195–201. [Google Scholar] [CrossRef]
- Brożek, J.; Chłond, D. Morphology, arrangement and classification of sensilla on the apical segment of labium in Peiratinae (Hemiptera: Heteroptera: Reduviidae). Zootaxa 2010, 2476, 39–52. [Google Scholar] [CrossRef]
- McIver, S.B. Structure of cuticular mechanoreceptors of arthropods. Ann. Rev. Entomol. 1975, 20, 381–397. [Google Scholar] [CrossRef]
- Chapman, R.F. Communication. In The Insects: Structure and Function, 4th ed.; Chapman, R.F., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 610–652. [Google Scholar]
- Gullan, P.J.; Cranston, P.S. (Eds.) Sensory Systems and Behavior. In The Insects: An Outline of Entomology, 5th ed.; Wiley Blackwell: Oxford, UK, 2014; pp. 85–112. [Google Scholar]
- Backus, E.A. Anatomical and sensory mechanisms of leafhopper and planthopper feeding behavior. In The Leafhoppers and Planthoppers; Nault, L.R., Rodriguez, J.G., Eds.; John Wiley & Sons: New York, NY, USA, 1985; pp. 163–194. [Google Scholar]
- Chapman, R.F. Chemosensory regulation of feeding. In Regulatory Mechanisms in Insects Feeding; Chapman, R.F., de Boer, G., Eds.; Springer Science + Business Media: Dordrecht, The Netherlands, 1995; pp. 101–136. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
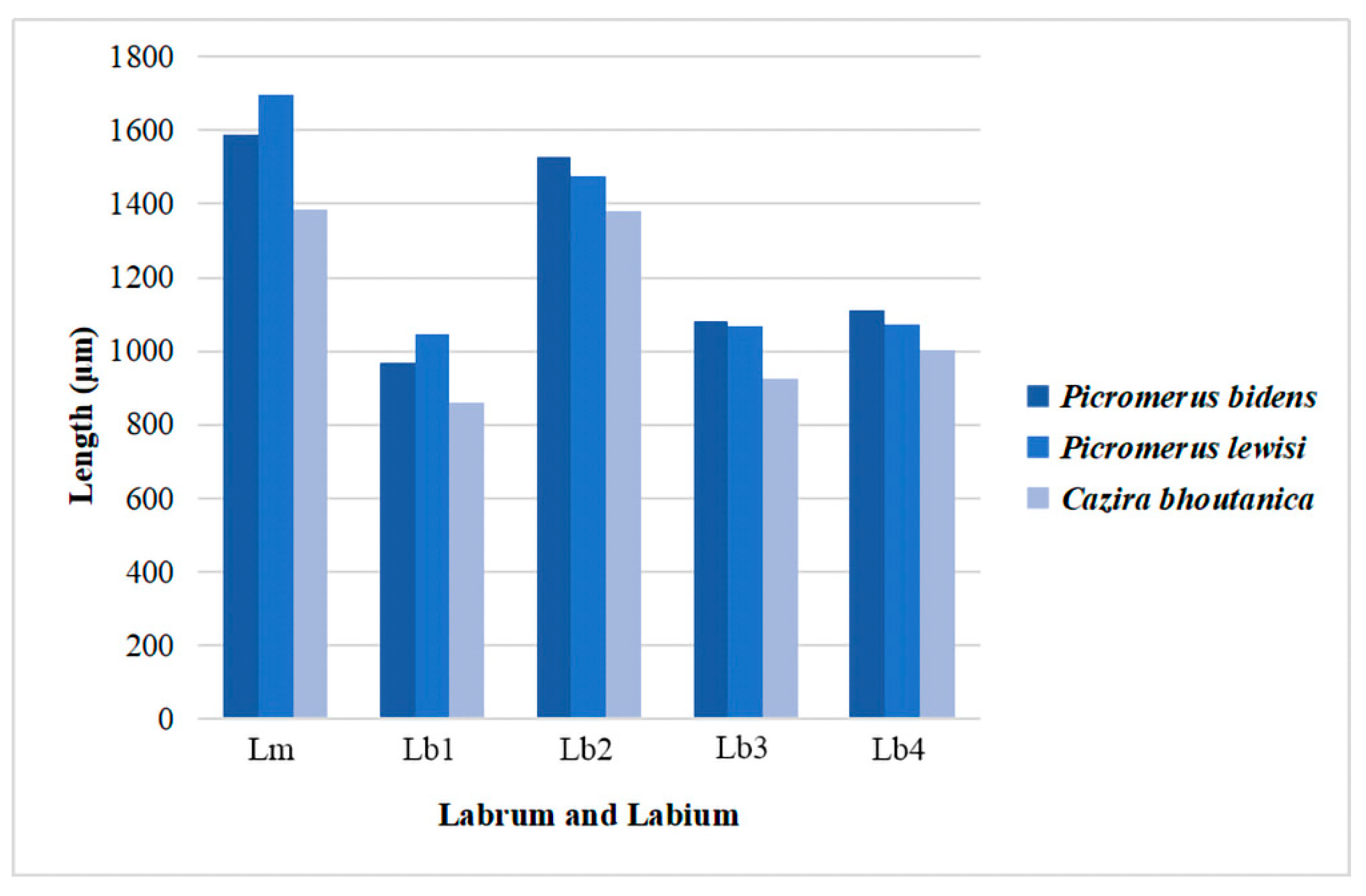
| Species | Labrum | Labial Segments | ||||
|---|---|---|---|---|---|---|
| I (μm) | II (μm) | III (μm) | IV (μm) | Whole | ||
| Picromerus bidens (Linnaeus) | 1586.8 ± 10.6 | 967.3 ± 8.9 | 1527.7 ± 33.2 | 1080.5 ± 34.5 | 1113.4± 24.4 | 4687.8 ± 93.9 |
| Picromerus lewisi Scott | 1694.8 ± 55.9 | 1046.0 ± 12.1 | 1474.1 ± 74.2 | 1068.3 ± 9.2 | 1073.9 ± 17.8 | 4661.5 ± 89.2 |
| Cazira bhoutanica Schouteden | 1385.9 ± 2.9 | 862.1 ± 7.4 | 1379.1 ± 28.6 | 925.2 ± 7.4 | 1001.2 ± 12.5 | 4179.8 ± 43.1 |
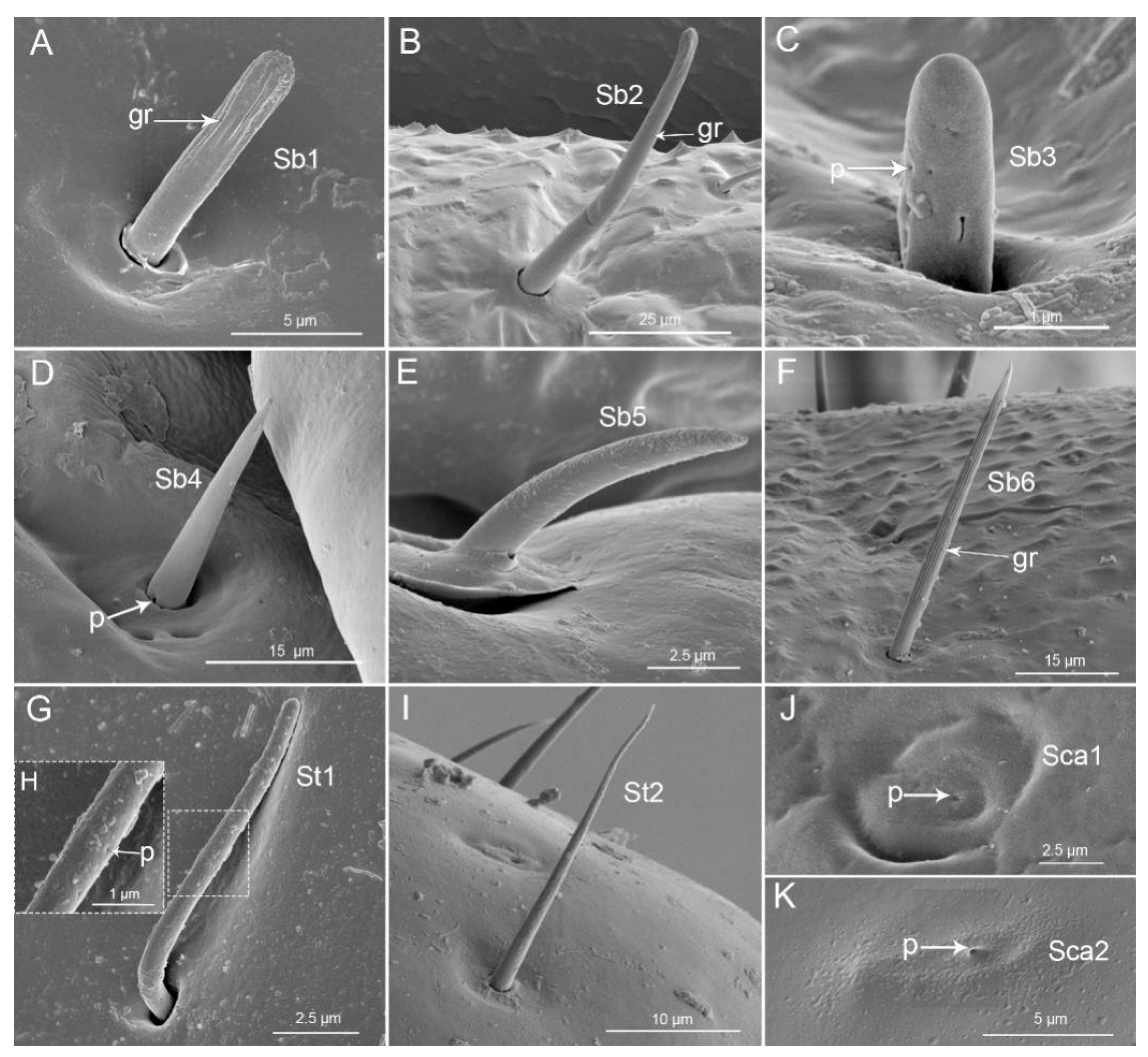
| Sensilla | Picromerus bidens (Linnaeus) | Picromerus lewisi Scott | Cazira bhoutanica Schouteden | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Distri- bution | Length (μm) | Basal Diameter (μm) | N | Distri- bution | Length (μm) | Basal Diameter (μm) | N | Distri- bution | Length (μm) | Basal Diameter (μm) | N | |
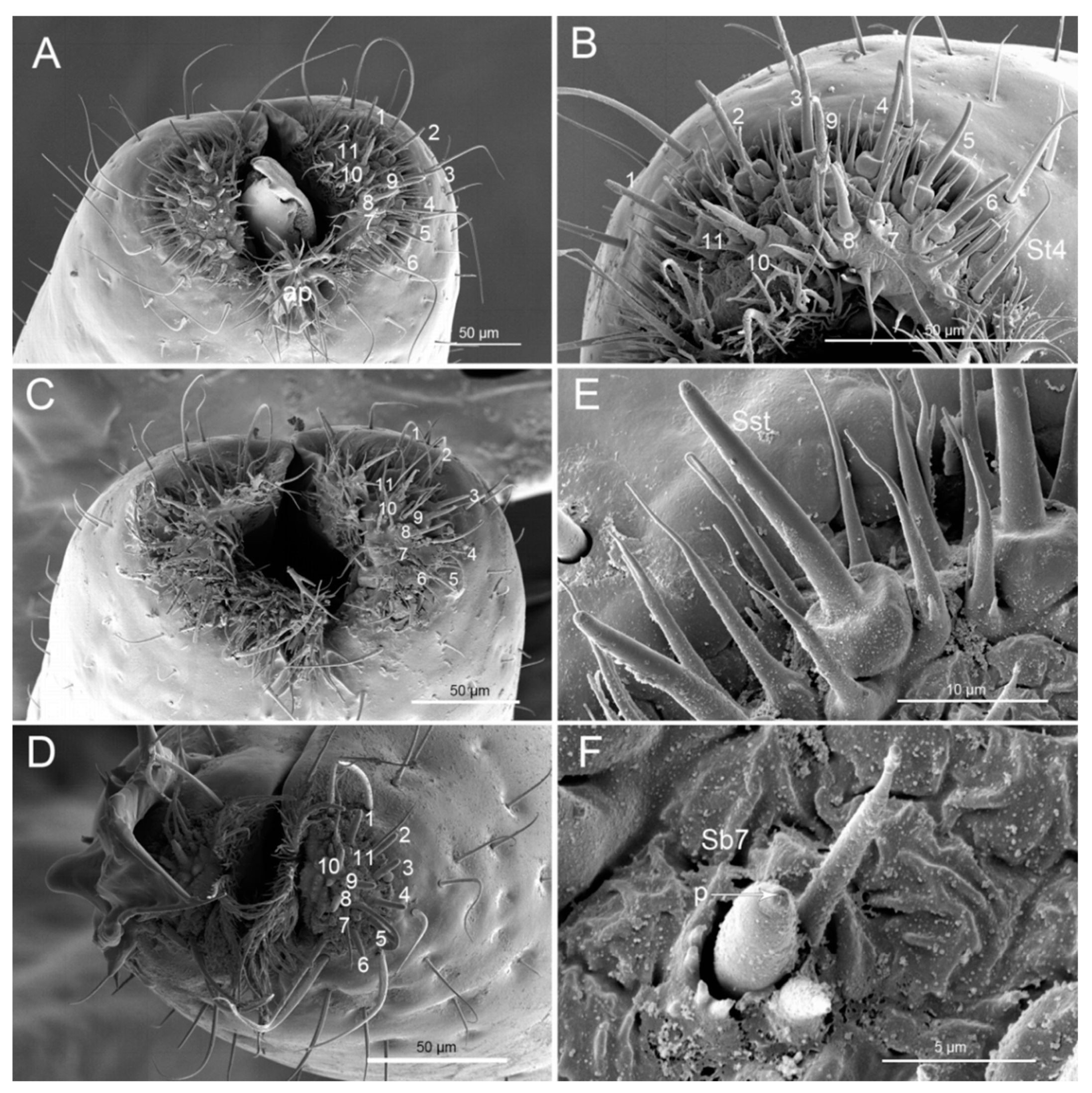
| St1 | Lb1-4 | 22.9 ± 1.2 | 0.83 ± 0 | 10 | Lb1-4 | 16.4 ± 1.5 | 0.8 ± 0 | 10 | ||||
| St2 | Lb4 | 17.3 ± 1.8 | 1.6 ± 0.1 | 8 | Lb4 | 20.7 ± 1.1 | 1.2 ± 0.1 | 10 | Lb4 | 22.5 ± 2.0 | 1.7 ± 0.1 | 7 |
| St3 | Lb4 | 63.5 ± 1.9 | 2.1 ± 0.1 | 5 | Lb4 | 66 ± 4.9 | 2.3 ± 0.1 | 6 | Lb4 | 97.6 ± 9.8 | 2.5 ± 0.1 | 6 |
| St4 | SF | 15.6 ± 0.8 | 1.7 ± 0.1 | 10 | SF | 15.5 ± 1.1 | 1.6 ± 0.1 | 10 | SF | 4.2 ± 0.3 | 1.2 ± 0.1 | 6 |
| Sb1 | Lm, Lb1-4 | 9.4 ± 0.6 | 1.7 ± 0.1 | 10 | Lm, Lb1 | 12 ± 1.8 | 1.9 ± 0.1 | 10 | Lm, Lb1 | 9.2 ± 0.7 | 1.5 ± 0.1 | 9 |
| Sb2 | Lb1 | 67.8 ± 9.5 | 3.8 ± 0.2 | 5 | Lm, Lb2 | 38 ± 5.9 | 3.7 ± 0.6 | 7 | Lb1, 2 | 56.2 ± 4.6 | 4.3 ± 0.2 | 4 |
| Sb3 | Lm | 1.7 ± 0.6 | 0.7 ± 0 | 3 | Lm, Lb1-3 | 1.1 ± 0.1 | 0.8 ± 0.1 | 3 | ||||
| Sb4 | Lb2, Lb4 | 23.3 ± 0.7 | 4.2 ± 0.1 | 5 | Lb2, Lb4 | 18.3 ± 0.6 | 3.6 ± 0.1 | 4 | Lb2, Lb4 | 11.2 ± 1.3 | 3.5 ± 0.2 | 4 |
| Sb5 | Lb1, Lb3 | 14.9 ± 0.4 | 1.8 ± 0.1 | 3 | Lb1,2,4 | 8.7 ± 0.7 | 1.4 ± 0.1 | 4 | Lb2 | 10.9 ± 1.0 | 1.8 ± 0.1 | 5 |
| Sb6 | Lb1-4 | 18 ± 1.4 | 1.5 ± 0.1 | 8 | Lb3,4 | 40.3 ± 0.4 | 3.5 ± 0.4 | 3 | Lb2-4 | 75.3 ± 3.9 | 4.1 ± 0.1 | 3 |
| Sb7 | SF | 4.7 ± 0.4 | 2.6 ± 0.1 | 4 | SF | 4.4 ± 0.8 | 1.4 ± 0.1 | 4 | SF | 4.9 ± 0.2 | 2.5 ± 0.1 | 4 |
| Sst | SF | 22.8 ± 0.9 | 7.3 ± 0.2 | 18 | SF | 22.8 ± 0.5 | 7.5 ± 0.3 | 10 | SF | 11.6 ± 0.8 | 5.1 ± 0.2 | 9 |
| Sca1 | Lm | 7.1 ± 0.5 | 4 | Lm, Lb2 | 6.7 ± 0.2 | 3 | Lm, Lb2, 4 | 9.1 ± 0.9 | 3 | |||
| Sca2 | Lb4 | 5.5 ± 0.3 | 12 | Lb4 | 5.8 ± 0.7 | 10 | Lb4 | 7.4 ± 0.4 | 4 | |||
| Type | Shape | Socket | Surface | Pore | Category | Function |
|---|---|---|---|---|---|---|
| St1 | Hair in pit | Inflexible | Smooth | Wp (porous) diameter 0.043 (μm) = 43 nm | Chemoreceptive sensilla | Olfactory |
| St2 | Hair | Flexible | Smooth | No | Mechanoreceptive sensilla | Tactile |
| St3 | Hair | Flexible | Smooth | No | Mechanoreceptive sensilla | Tactile |
| St4 | Hair | Inflexible | Smooth | No | Mechanoreceptive sensilla | Tactile |
| Sb1 | Peg | Flexible | Grooved | No | Mechanoreceptive sensilla | Tactile |
| Sb2 | Peg | Flexible | Grooved | No | Mechanoreceptive sensilla | Tactile |
| Sb3 | Peg in pit | Inflexible | Smooth | Diameter 0.043 (μm) = 43 nm | Chemoreceptive sensilla | Olfactory |
| Sb4 | Peg | Flexible | Smooth | Wp (molting pore) | Proprioceptive sensilla | Perceive the degree of flexion of the joint |
| Sb5 | Peg | Flexible | Smooth | No | Mechanoreceptive sensilla | Tactile |
| Sb6 | Peg, hair | Flexible | Grooved | No | Mechanoreceptive sensilla | Tactile |
| Sb7 | Peg in pit | Inflexible | Smooth | TP | Chemoreceptive sensilla | Gustatory |
| Sst | Peg | Cone sitting on a style(high socket) | Smooth | TP | Chemoreceptive sensilla | Gustatory |
| Sca1 | Oval plate | Inflexible | Smooth | Central pore (molting pore) | Mechanoreceptive sensilla | Responding to strains in the exoskeletons |
| Sca2 | Domelike structures | Inflexible | Smooth | Central pore (molting pore) | Mechanoreceptive sensilla | Responding to strains in the exoskeletons |
| Feeding Behavior | Species | Labrum | Labium | Number of Sensilla Types | The Number of Sensilla of Labium Tip | Apical Plate | Internal Side of Maxillary Stylet | Internal Morphology of Mandibular Stylet | Distal Mandibular Stylet; Serration | References |
|---|---|---|---|---|---|---|---|---|---|---|
| Predatory | Canthecona furcellata (Wolff) | triangular sclerite | elongated | 3 types | unreported | unreported | dentations | unreported | serrated | Barsagade & Gathalkar [28] |
| Predatory | Eocanthecona furcellata (Wolff) | unreported | robust and stout | 6 types | 3 types | unreported | unreported | unreported | unreported | Rani [26]; Parveen et al. [27] |
| Predatory | Perillus bioculatus Fabricius | unreported | robust and stout | 6 types | 3 types | membranous lobe microtrichial extensions | unreported | squamous texture | sharp recurved hooks | Cobben [1]; Parveen et al. [27] |
| Predatory | Podisus maculiventris (Say) | unreported | unreported | unreported | unreported | present | unreported | unreported | teeth | Cohen [8] |
| Predatory | Picromerus bidens (Linnaeus) | elongated, triangular | robust and stout | 14 types | 3 types | membranous lobe microtrichial extensions | short barbs | squamous textures and cuticular spines | five teeth and three hooks | This study |
| Predatory | Picromerus lewisi Scott | elongated, triangular | robust and stout | 14 types | 3 types | membranous lobe microtrichial extensions | short barbs | squamous textures and cuticular spines | five teeth and three hooks | This study |
| Predatory | Cazira bhoutanica Schouteden | elongated, triangular | robust and stout | 14 types | 3 types | membranous lobe microtrichial extensions | short barbs | squamous textures and cuticular spines | five teeth and three hooks | This study |
| Phytophagous | Graphosoma lineatum (Linnaeus) | unreported | unreported | unreported | unreported | unreported | unreported | small, widely spaced notches arranged in longitudinal strips | complex ribbed texture, the apices are knotted with irregular prominences | Cobben [1] |
| Phytophagous | Chrysocoris purpurea (Westwood) | unreported | unreported | unreported | 2 types | present | unreported | unreported | knotted with irregular prominences | Rani & Madhavendra [25] |
| Phytophagous | Cyclopelta siccifolia Westwood | unreported | unreported | unreported | 2 types | present | unreported | unreported | knotted with irregular prominences | Rani & Madhavendra [25] |
| Phytophagous | Dichelops melacanthus (Dallas) | unreported | unreported | unreported | unreported | unreported | unreported | squamous texture | dentition | Depieri & Panizzi [12] |
| Phytophagous | Dolycoris indicus (Stål) | unreported | slender and long | 6 types | 2 types | unreported | unreported | unreported | unreported | Parveen et al. [27] |
| Phytophagous | Euschistus heros Fabricius | unreported | unreported | unreported | unreported | unreported | unreported | squamous texture | dentition | Depieri et al. [13]; Depieri & Panizzi [12] |
| Phytophagous | Nezara viridula (Linnaeus) | unreported | unreported | unreported | 2 types | present | grooves | squamous texture | knotted with irregular prominences | Rani and Madhavendra [24]; Depieri & Panizzi [12] |
| Phytophagous | Piezodorus guildinii (Westwood) | unreported | unreported | unreported | unreported | unreported | unreported | squamous texture | dentition | Depieri & Panizzi [12] |
| Phytophagous | Piezodorus hybneri (Gmelin) | unreported | slender and long | 6 types | 2 types | unreported | unreported | unreported | unreported | Parveen et al. [27] |
| Phytophagous | Plautia crossota (Dallas) | unreported | slender and long | 6 types | 2 types | unreported | unreported | unreported | unreported | Parveen et al. [27] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Brożek, J.; Dai, W. Comparative Morphology of the Mouthparts in Three Predatory Stink Bugs (Heteroptera: Asopinae) Reveals Feeding Specialization of Stylets and Sensilla. Insects 2020, 11, 762. https://doi.org/10.3390/insects11110762
Wang Y, Brożek J, Dai W. Comparative Morphology of the Mouthparts in Three Predatory Stink Bugs (Heteroptera: Asopinae) Reveals Feeding Specialization of Stylets and Sensilla. Insects. 2020; 11(11):762. https://doi.org/10.3390/insects11110762
Chicago/Turabian StyleWang, Yan, Jolanta Brożek, and Wu Dai. 2020. "Comparative Morphology of the Mouthparts in Three Predatory Stink Bugs (Heteroptera: Asopinae) Reveals Feeding Specialization of Stylets and Sensilla" Insects 11, no. 11: 762. https://doi.org/10.3390/insects11110762
APA StyleWang, Y., Brożek, J., & Dai, W. (2020). Comparative Morphology of the Mouthparts in Three Predatory Stink Bugs (Heteroptera: Asopinae) Reveals Feeding Specialization of Stylets and Sensilla. Insects, 11(11), 762. https://doi.org/10.3390/insects11110762