Biodiversity Inventory and Distribution of Metriorrhynchina Net-Winged Beetles (Coleoptera: Lycidae), with the Identification of Generic Ranges


Simple Summary
Abstract
1. Introduction
2. Methods
3. Results and Discussion
3.1. The Justification of Nomenclatural Acts Proposed in this Study
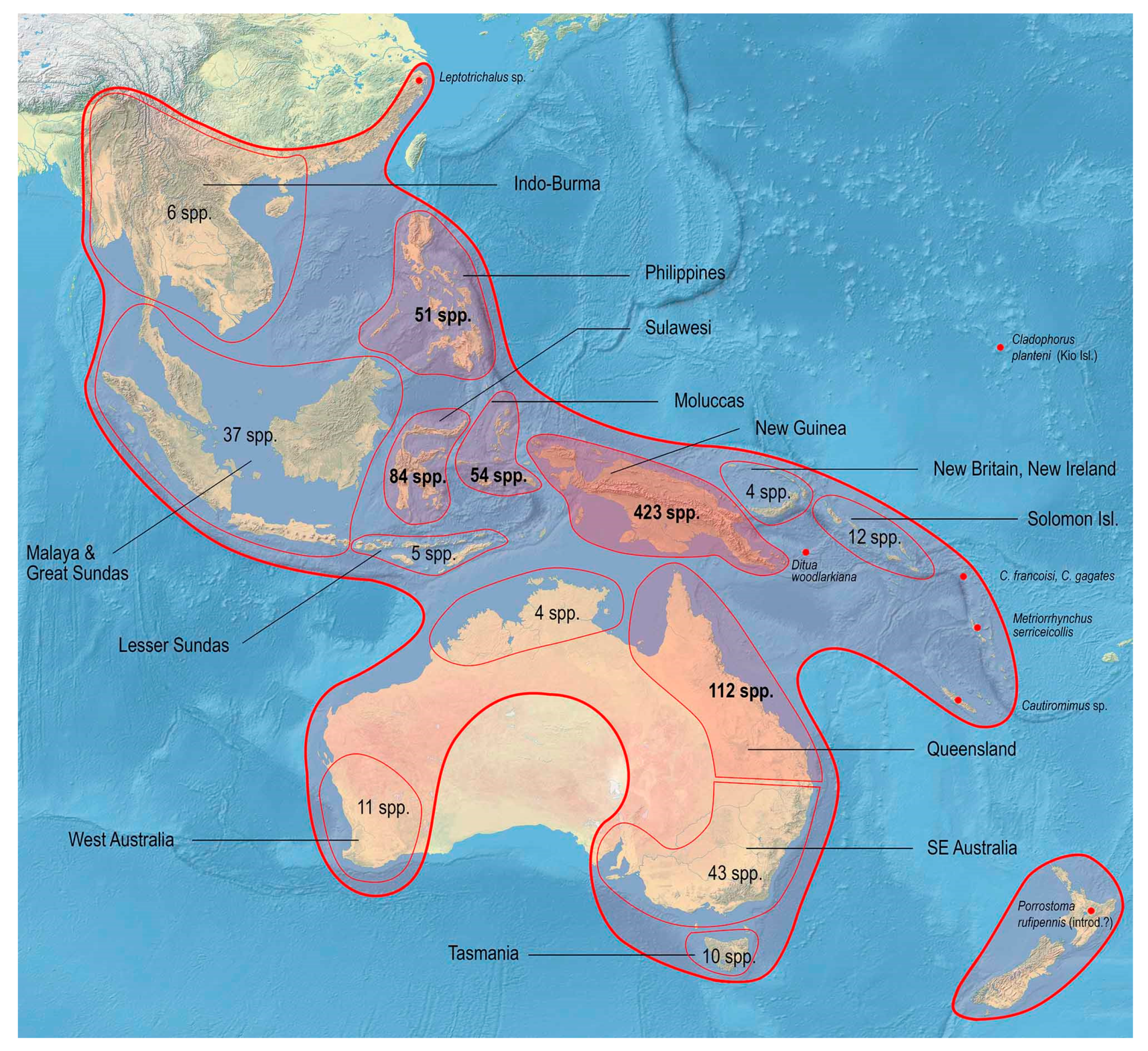
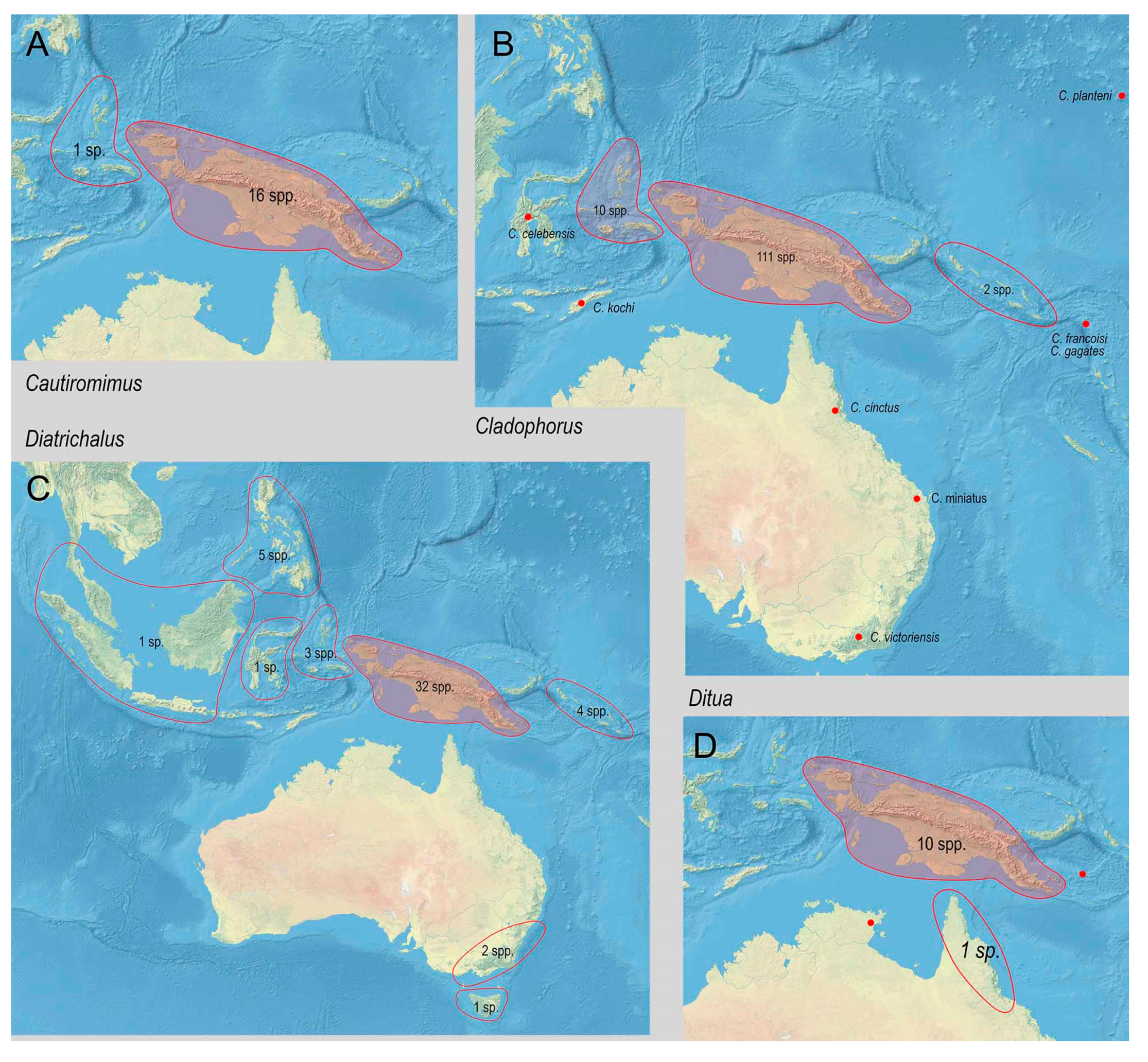
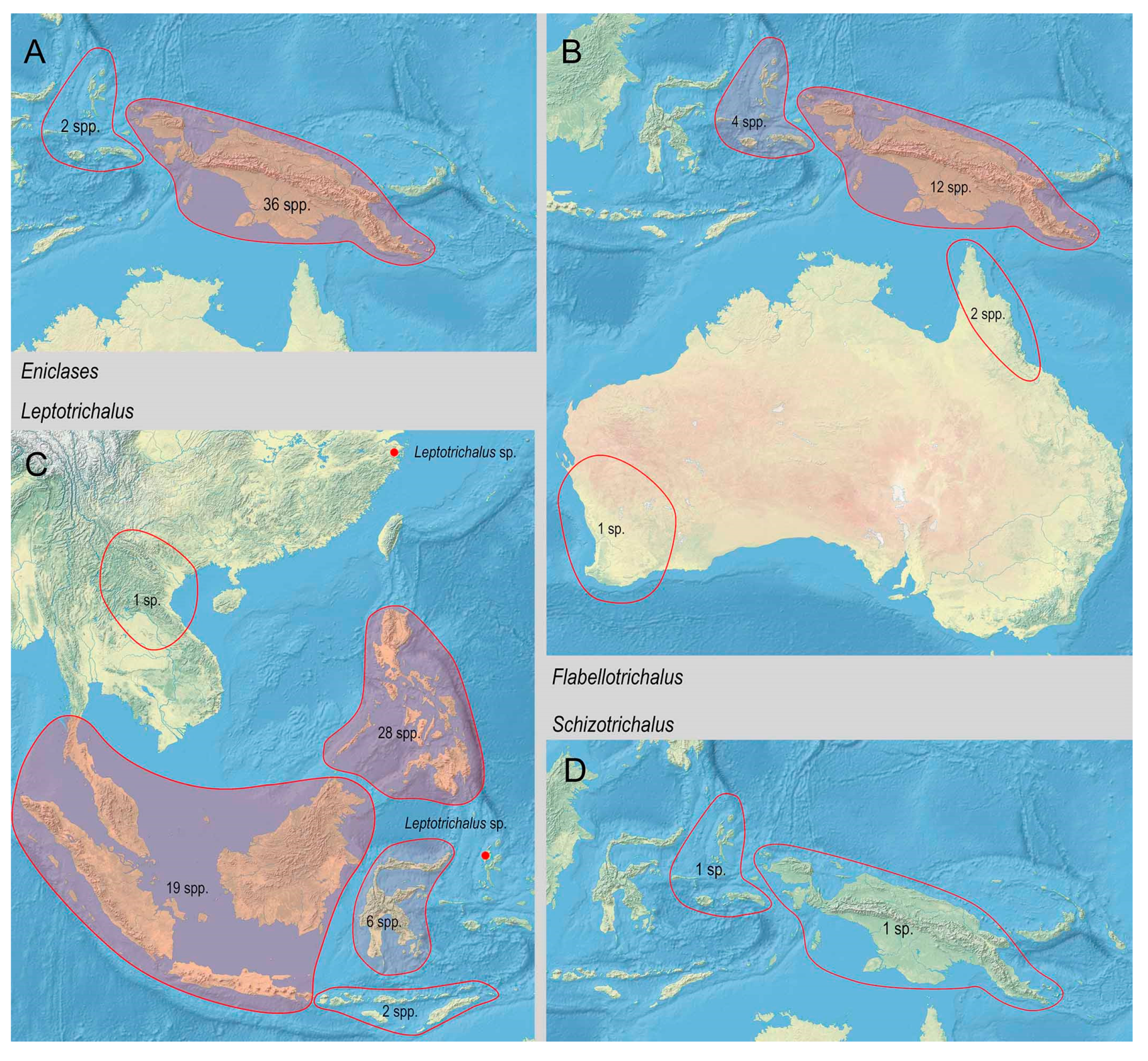
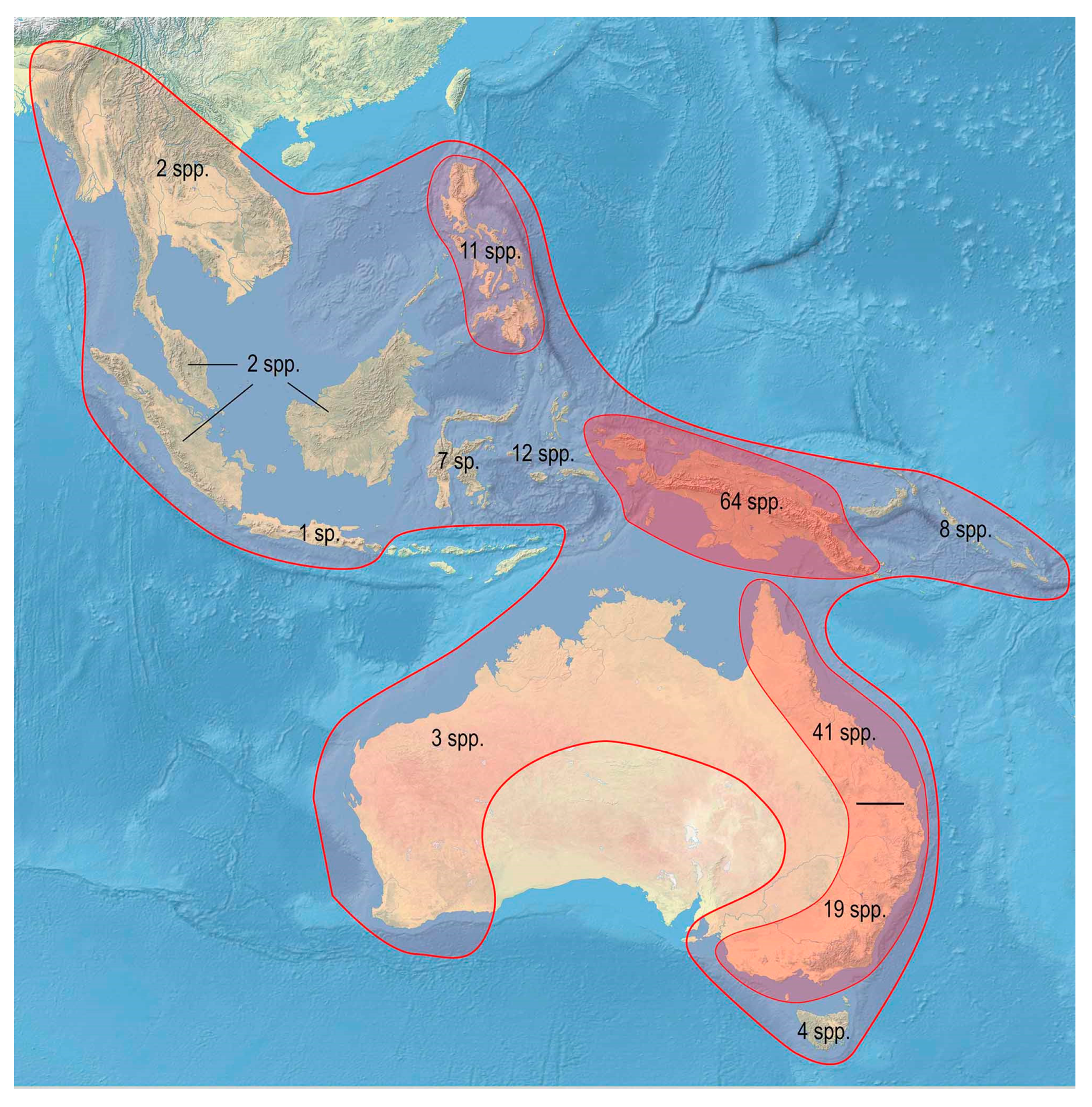
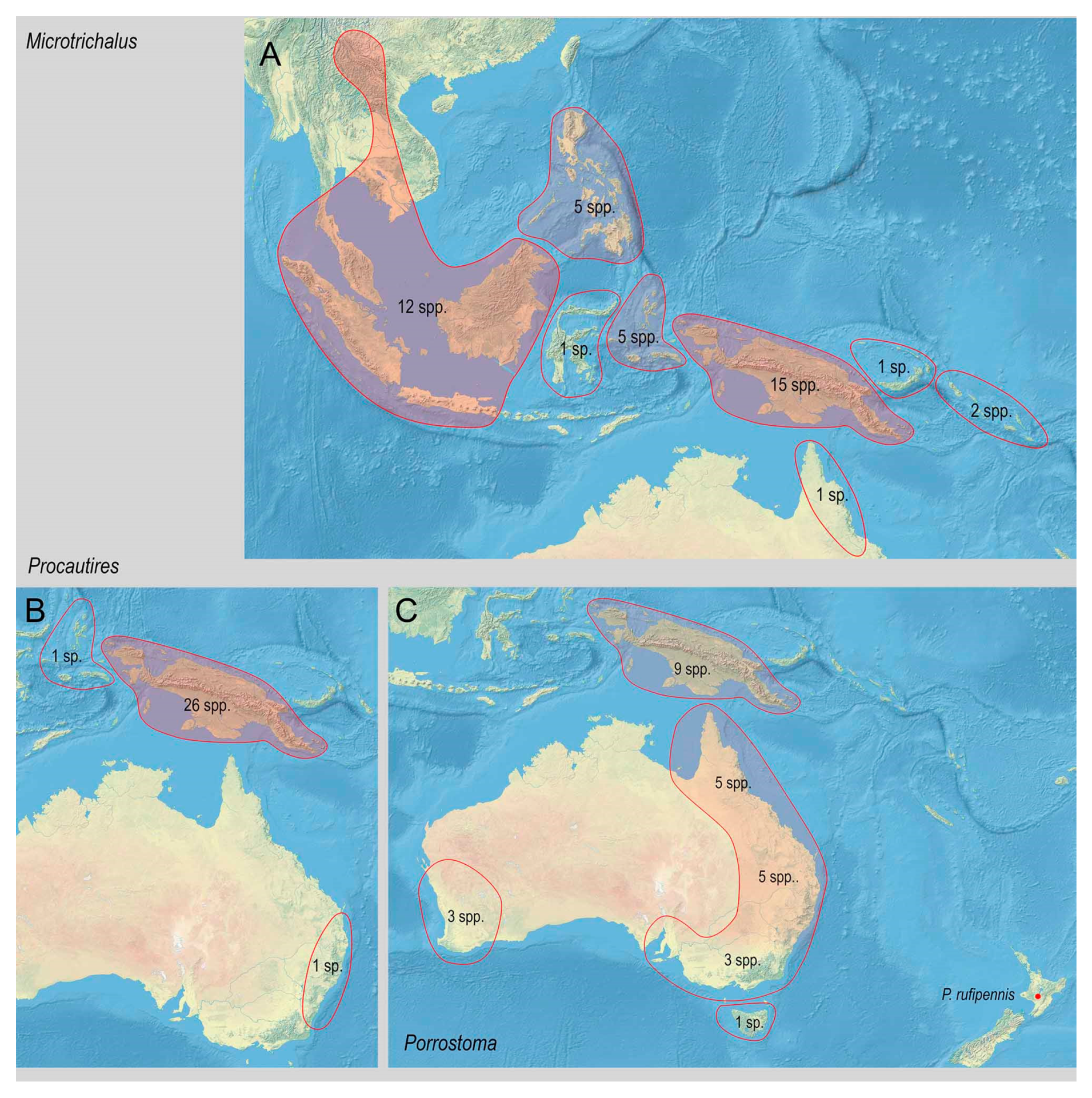
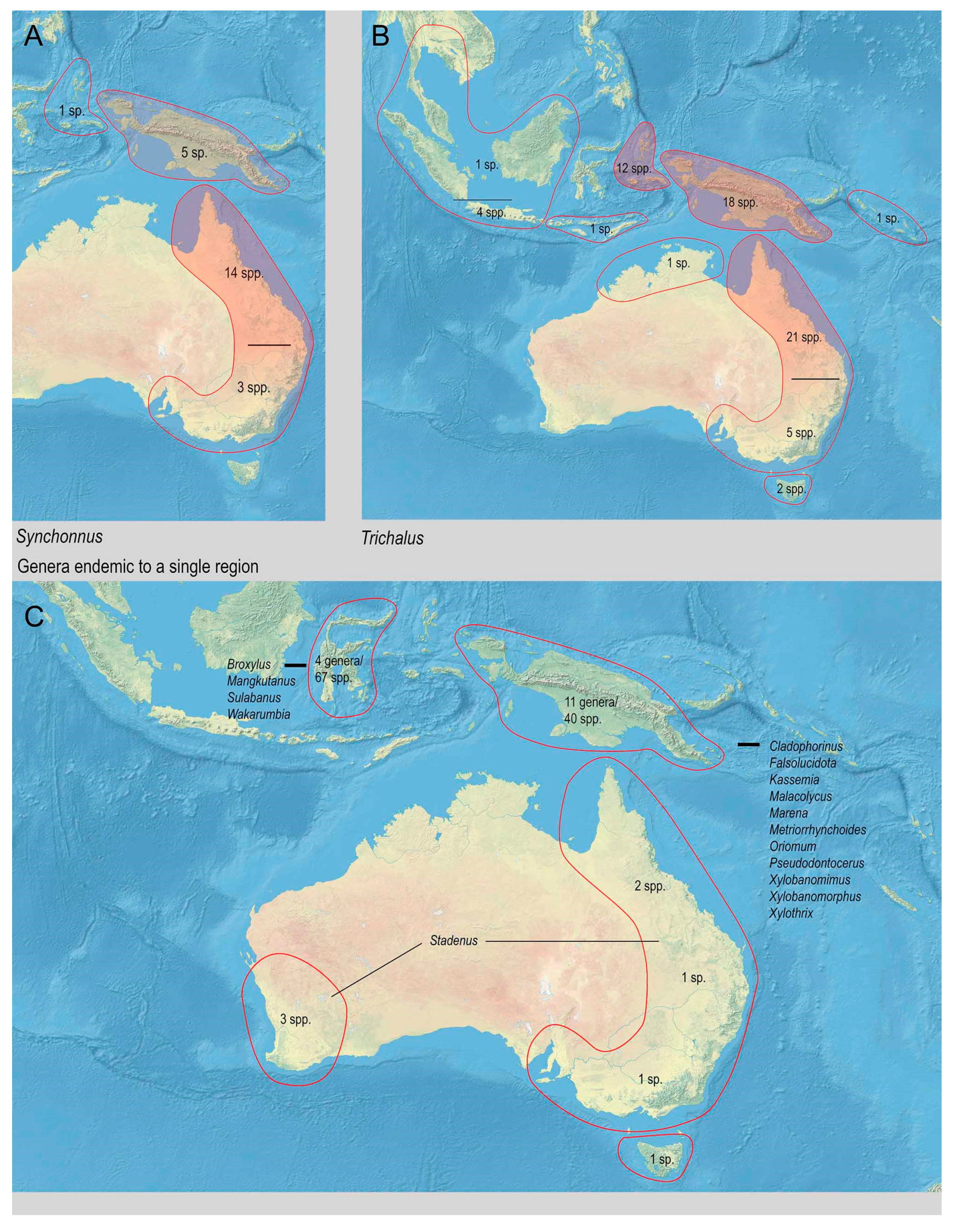
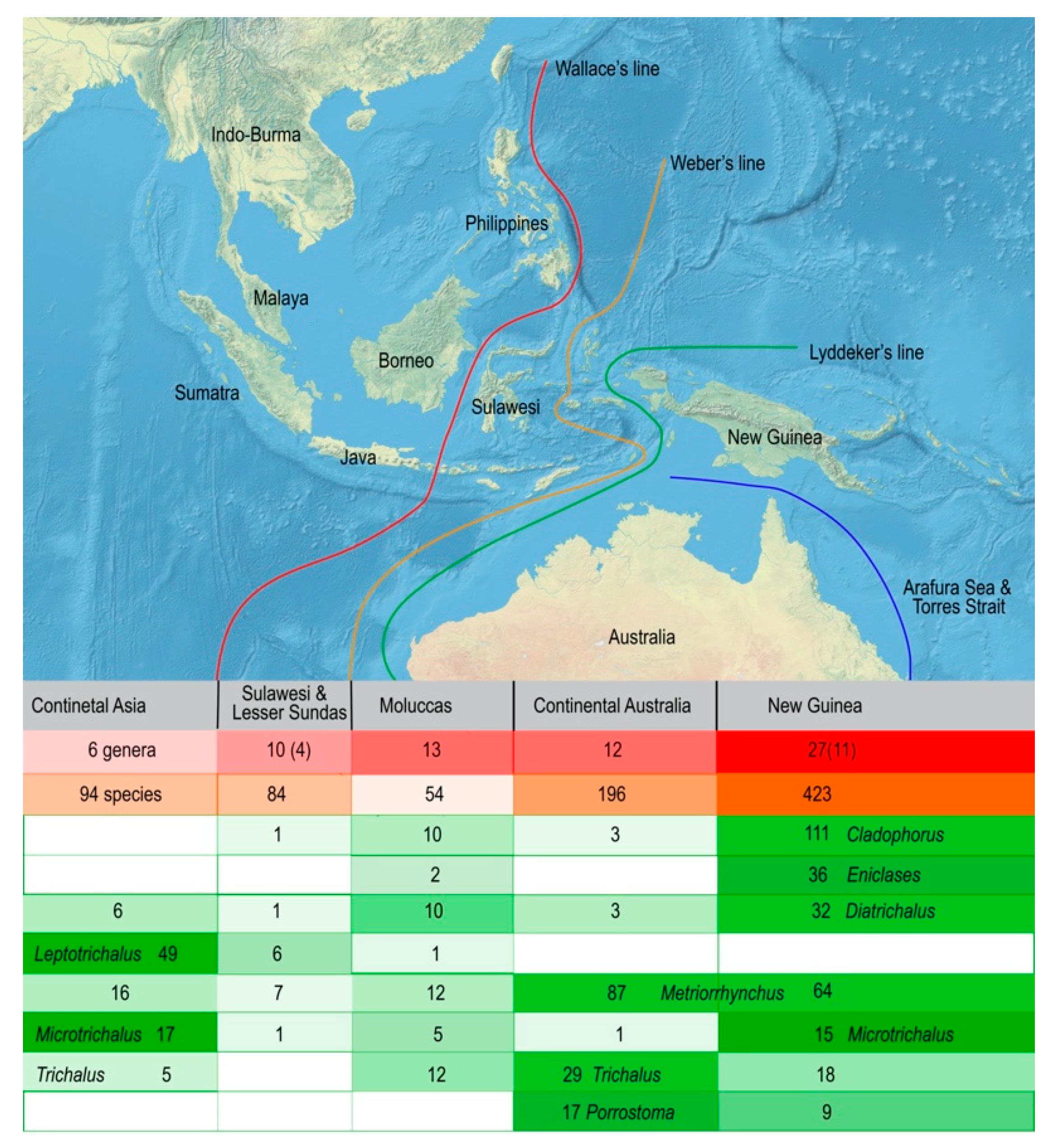
3.2. Biodiversity and Zoogeography of the Metriorrhynchina
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bocak, L.; Barton, C.; Crampton-Platt, A.; Chesters, D.; Ahrens, D.; Vogler, A.P. Building the Coleoptera tree-of-life for >8000 species: Composition of public DNA data and fit with Linnaean classification. Syst. Entomol. 2014, 39, 97–110. [Google Scholar] [CrossRef]
- Masek, M.; Motyka, M.; Kusy, D.; Bocek, M.; Li, Y.; Bocak, L. Molecular Phylogeny, Diversity and Zoogeography of Net-Winged Beetles (Coleoptera: Lycidae). Insects 2018, 9, 154. [Google Scholar] [CrossRef] [PubMed]
- Kleine, R. Lycidae. Part 128. In Coleopterorum Catalogue Auspiciis et Auxilio; Schenkling, S., Ed.; W. Junk: Berlin, Germany, 1933; p. 145. [Google Scholar]
- Dvorak, M.; Bocak, L. Sulabanus gen. nov., a new genus of Lycidae (Coleoptera) from Sulawesi. Zootaxa 2007, 1611, 1–24. [Google Scholar] [CrossRef]
- Dvorak, M.; Bocak, L. Ten new species of Wakarumbia Bocak, 1999 from Sulawesi (Coleoptera: Lycidae), with a key to males of the genus. Zootaxa 2009, 2282, 51–61. [Google Scholar] [CrossRef]
- Kazantsev, S.V. New Xylobanus and related taxa of net-winged beetles from New Guinea (Coleoptera: Lycidae). Russ. Entomol. J. 2015, 24, 107–118. [Google Scholar] [CrossRef]
- Kazantsev, S.V. New species of Metriorrhynchus Gemminger et Harold, 1869 and Porrostoma Laporte, 1838 from New Guinea (Coleoptera: Lycidae). Russ. Entomol. J. 2015, 24, 211–233. [Google Scholar] [CrossRef]
- Kazantsev, S.V. New species of Procautires Kleine, 1925 from New Guinea (Coleoptera: Lycidae). Russ. Entomol. J. 2016, 25, 35–47. [Google Scholar] [CrossRef]
- Kazantsev, S.V. New and little known species of Falsolucidota Pic, 1921 from New Guinea (Coleoptera: Lycidae). Russ. Entomol. J. 2016, 25, 355–360. [Google Scholar] [CrossRef]
- Kusy, D.; Sklenarova, K.; Bocak, L. The effectiveness of DNA-based delimitation in Synchonnus net-winged beetles (Coleoptera: Lycidae) assessed, and description of 11 new species. Austral Entomol. 2017, 57, 25–39. [Google Scholar] [CrossRef]
- Bocak, L.; Bocakova, M. Revision of the supergeneric classification of the family Lycidae (Coleoptera). Pol. Pismo Entomol. 1990, 59, 623–676. [Google Scholar]
- Bocak, L.; Bocakova, M. Phylogeny and classification of the family Lycidae (Insecta: Coleoptera). Ann. Zool. 2008, 58, 695–720. [Google Scholar] [CrossRef]
- Bocak, L.; Bocakova, M.; Hunt, T.; Vogler, A.P. Multiple ancient origins of neoteny in Lycidae (Coleoptera): Consequences for ecology and macroevolution. Proc. R. Soc. B Biol. Sci. 2008, 275, 2015–2023. [Google Scholar] [CrossRef] [PubMed]
- Kazantsev, S.V. Morphology of Lycidae with some considerations on evolution of the Coleoptera. Elytron 2005, 17, 49–226. [Google Scholar]
- Kazantsev, S.V. New and little known taxa of neotenic Lycidae (Coleoptera), with discussion of their phylogeny. Russ. Entomol. J. 2013, 22, 9–31. [Google Scholar]
- Kusy, D.; Motyka, M.; Bocek, M.; Masek, M.; Bocak, L. Phylogenomic analysis resolves the relationships among net-winged beetles (Coleoptera: Lycidae) and reveals the parallel evolution of morphological traits. Syst. Entomol. 2019, 44, 911–925. [Google Scholar] [CrossRef]
- Bocak, L. Revision and phylogenetic analysis of Metriorrhynchinae. Eur. J. Entomol. 2002, 241, 315–351. [Google Scholar] [CrossRef]
- Bocak, L.; Matsuda, K. Review of the immature stages of the family Lycidae (Insecta: Coleoptera). J. Nat. Hist. 2003, 37, 1463–1507. [Google Scholar] [CrossRef]
- Levkanicova, Z.; Bocak, L. Identification of net-winged beetle larvae (Coleoptera: Lycidae) using three mtDNA fragments: A comparison of their utility. Syst. Entomol. 2009, 34, 210–221. [Google Scholar] [CrossRef]
- Sklenarova, K.; Kubecek, V.; Bocak, L. Subtribal classification of Metriorrhynchini (Insecta: Coleoptera: Lycidae): An integrative approach using molecular phylogeny and morphology of adults and larvae. Arthr. Syst. Phylogeny 2014, 72, 37–54. [Google Scholar]
- Motyka, M.; Kampova, L.; Bocak, L. Phylogeny and evolution of Müllerian mimicry in aposematic Dilophotes: Evidence for advergence and size-constraints in evolution of mimetic sexual dimorphism. Sci. Rep. 2018, 8, 3744. [Google Scholar] [CrossRef]
- Motyka, M.; Bocek, M.; Kusy, D.; Bocak, L. Interactions in multi-pattern Müllerian communities support origins of new patterns, false structures, imperfect resemblance and mimetic sexual dimorphism. Sci. Rep. 2020, 10, 11193. [Google Scholar] [CrossRef] [PubMed]
- Bocek, M.; Kusy, D.; Motyka, M.; Bocak, L. Persistence of multiple patterns and intraspecific polymorphism in multi-species Müllerian communities of net-winged beetles. Front. Zool. 2019, 16, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Masek, M.; Palata, V.; Bray, T.C.; Bocak, L. Molecular phylogeny reveals high diversity and geographic structure in Asian neotenic net-winged beetles Platerodrilus (Coleoptera: Lycidae). PLoS ONE 2015, 10, e0123855. [Google Scholar] [CrossRef]
- Kazantsev, S.V. Comparative morphology of mandibular structures in lycid larvae and its phylogenetic implications (Polyphaga, Hexapoda). Acta Zool. 2006, 87, 229–238. [Google Scholar] [CrossRef]
- Beutel, R.G.; Bocak, L.; Bocakova, M. Are Polyphaga (Coleoptera) really a basal neopteran lineage—A reply to Kazantsev. Acta Zool. 2007, 88, 153–158. [Google Scholar] [CrossRef]
- Calder, A.A. Coleoptera: Elateroidea. In Zoological Catalogue of Australia; Wells, A., Ed.; CSIRO Publishing: Melbourne, Australia, 1998; Volume 29.6, p. 248. [Google Scholar]
- Pic, M. Contribution a l’étude des Lycides. L’Echange, Rev. Linn. 1921, 404–406, 1–12. [Google Scholar]
- Pic, M. Malacodermes exotiques. L’Echange, Rev. Linn. 1923, 418–419, 1–84. [Google Scholar]
- Kubecek, V.; Bray, T.C.; Bocak, L. Molecular phylogeny of Metanoeina net-winged beetles identifies Ochinoeus, a new genus from China and Laos (Coleoptera: Lycidae). Zootaxa 2015, 3955, 113–122. [Google Scholar] [CrossRef]
- Dudkova, P.; Bocak, L. A review of the Cautires obsoletus species group from Indo–Burma (Coleoptera: Lycidae). Zootaxa 2010, 2527, 28–48. [Google Scholar] [CrossRef]
- Weiszenstein, M.; Bocak, L. A review of the genus Metanoeus from the Philippines (Coleoptera: Lycidae). Raffl. Bull. Zool. 2011, 59, 163–170. [Google Scholar]
- Bocak, L. Nomenclatural notes on taxa of the family Lycidae described by Guérin-Méneville (Insecta: Coleoptera). Ann. Zool. 1998, 48, 245–251. [Google Scholar]
- Bocak, L. A generic revision and phylogenetic analysis of the subtribe Trichalinina (Coleoptera: Lycidae: Metriorrhynchini). Acta Soc. Zool. Boh. 1998, 62, 167–200. [Google Scholar]
- Bocak, L.; Jašš, R. Revision of the genus Broxylus Waterhouse (Coleoptera, Lycidae). Dtsch. Entomol. Z. 2004, 51, 65–75. [Google Scholar] [CrossRef]
- Kazantsev, S.V. On the status and systematic position of the genera Falsolucidota Pic, 1921 and Macrolycinella Pic, 1922, with notes on Lopheros LeConte, 1881 (Coleoptera: Lycidae). Russ. Entomol. J. 2009, 18, 277–283. [Google Scholar]
- Kazantsev, S.V. A review of Ditua Waterhouse, 1879 with description of seven new species from New Guinea (Coleoptera: Lycidae). Russ. Entomol. J. 2016, 25, 239–248. [Google Scholar] [CrossRef]
- Bocak, L. A revision of the genus Microtrichalus Wat. from the Philippines (Coleoptera: Lycidae). Eur. J. Entomol. 1998, 95, 417–428. [Google Scholar]
- Bocak, L. A revision of the genus Diatrichalus Kleine from New Guinea (Coleoptera: Lycidae). Stuttg. Beitr. Naturk. A 2001, 622, 1–32. [Google Scholar]
- Bocak, L. Revision of the genus Wakarumbia (Coleoptera: Lycidae). Eur. J. Entomol. 2000, 97, 271–278. [Google Scholar] [CrossRef]
- Bocak, L. To the knowledge of the genus Leptotrichalus Kleine from Sumatra, Borneo and Continental Asia (Coleoptera: Lycidae). Ann. Soc. Entomol. Fr. 2000, 36, 71–184. [Google Scholar]
- Bocak, L. A revision of the genus Diatrichalus Kleine from the Philippines (Coleoptera: Lycidae). Raffl. Bull. Zool. 2000, 48, 11–16. [Google Scholar]
- Bocak, L. New species of the genus Wakarumbia from Sulawesi (Coleoptera: Lycidae). Eur. J. Entomol. 2001, 49, 1–9. [Google Scholar]
- Bocak, L. A Revision of Metriorrhynchus (Coleoptera: Lycidae) from the Greater Sunda Islands and Continental. Asia. Eur. J. Entomol. 2007, 55, 253–260. [Google Scholar]
- Bocak, L.; Bocakova, M. Revision of the genus Hemiconderis (Coleoptera, Lycidae). Acta Entomol. Bohemoslov. 1990, 89, 209–220. [Google Scholar]
- Fabricius, J.C. Systema Eleutheratorum Secundum Ordines, Genera, Species: Adiectis Synonymis, Locis, Observationibus, Descriptionibus; Bibliopolii Academici: Kiliae, Germania, 1801. [Google Scholar]
- Dalman, J.W. Appendix ad C.J. Schonherr Synonymiam Insectorum Sistens Descriptiones Novarum Specierum; Schonherr, C.J., Ed.; Officina Lewerentziana: Scarfs, Sweden, 1818; Volume 1, Part 3; 266p. [Google Scholar]
- Waterhouse, C.O. A Monograph of the Australian species of the Coleopterous family Lycidae. Trans. Entomol. Soc. Lond. 1877, 73–86. [Google Scholar]
- Waterhouse, C.O. On the different forms occurring in the Coleopterous family Lycidæ, with descriptions of new genera and species. Trans. Entomol. Soc. Lond. 1878, 26, 95–118. [Google Scholar] [CrossRef]
- Waterhouse, C.O. Illustration of the Typical Specimens of Coleoptera in the Collection of the British Museum. Part I.—Lycidae; British Museum: London, UK, 1879; p. 83. [Google Scholar]
- Blackburn, T. Further notes on Australian Coleoptera, with descriptions of new species. Trans. R. Soc. S. Aust. 1888, 10, 177–287. [Google Scholar]
- Blackburn, T. Notes on Australian Coleoptera, with descriptions of new species. Proc. Linn. Soc. N. S. W. 1892, 6, 479–550. [Google Scholar] [CrossRef]
- Blackburn, T. Further notes on Australian Coleoptera, with descriptions of new genera and species. Part XXVI. Trans. R. Soc. S. Aust. 1900, 24, 35–68. [Google Scholar]
- Macleay, W.J. The insects of the Fly River, New Guinea, “Coleoptera”. Proc. Linn. Soc. N. S. W. 1886, 1, 136–157. [Google Scholar]
- Macleay, W.J. The Insects of the Cairns District, Northern Queensland. Proc. Linn. Soc. N. S. W. 1887, 2, 211–238. [Google Scholar] [CrossRef]
- Lea, A.M. Descriptions of new species of Australian Coleoptera. Proc. Linn. Soc. N. S. W. 1895, 9, 589–634. [Google Scholar]
- Lea, A.M. Descriptions of new species of Australian Coleoptera. Part V. Proc. Linn. Soc. N. S. W. 1899, 23, 521–645. [Google Scholar]
- Lea, A.M. The Coleoptera of King Island, Bass Strait. Proc. R. Soc. Vic. 1908, 20, 143–207. [Google Scholar]
- Lea, A.M. Revision of the Australian and Tasmanian Malacodermidae. Trans. Entomol. Soc. Lond. 1909, 45–251. [Google Scholar]
- Lea, A.M. On Coleoptera, mostly from Queensland. Mem. Qld Mus. 1909, 7, 182–240. [Google Scholar]
- Lea, A.M. On Australian Coleoptera of the family Malacodermidae. Trans. R. Soc. S. Aust. 1921, 45, 50–135. [Google Scholar]
- Lea, A.M. Descriptions of new species of Australian Coleoptera. Part XVII. Proc. Linn. Soc. N. S. W. 1921, 46, 351–369. [Google Scholar] [CrossRef]
- Lea, A.M. Results of Dr. E. Mjöberg’s Swedish scientific expeditions to Australia 1910–1913. Cryptophagidae, Cucujidae, Malacodermidae, Melandryidae, Mordellidae, Rhipidophoridae and Oedemeridae. Ark. Zool. 1922, 14, 1–21. [Google Scholar] [CrossRef]
- Lea, A.M. On Coleoptera, mostly from Queensland. Mem. Qld Mus. 1929, 9, 335–363. [Google Scholar]
- Kleine, R. Bestimmungstabelle der Gattung Trichalus. Treubia 1929, 10, 471–493. [Google Scholar]
- Guérin-Méneville, F.E. Crustacés, arachnides et insectes. In In Voyage autour du Monde sur la Corvette de Sa Majesté, La Coquille, pendant les annés 1822, 1823, 1824 et 1825. Zoologie; Duperrey, L.I., Ed.; Bertrand: Paris, France, 1830. [Google Scholar]
- Guérin-Méneville, F.E. Crustacés, arachnides et insectes. In Voyage Autour du Monde sur la Corvette de Sa Majesté, La Coquille, Pendant les Annés1822, 1823, 1824 et 1825. Zoologie. Vol. 2, Part 2; Duperrey, L.I., Ed.; Arthus Bertrand: Paris, France, 1838. [Google Scholar]
- Kleine, R. Fauna Buruana. Coleoptera, Fam. Lycidae. (2. Beitrag zur Kenntnis der Lycidae.). Treubia 1925, 7, 31–37. [Google Scholar]
- Kleine, R. Coleoptera Lycidae. Nova Guinea 1926, 13, 91–195. [Google Scholar]
- Kleine, R. Bericht ueber die von Miss Cheesman in Britisch Neu-Guinea gesammelten Brenthiden und Lyciden. Nova Guinea 1935, 17, 303–322. [Google Scholar]
- Bocek, M.; Bocak, L. Where are species limits in polymorphic mimetic beetles from New Guinean mountains: A case of Eniclases net-winged beetles (Lycidae: Metriorrhynchini). ZooKeys 2016, 593, 15–35. [Google Scholar]
- Bocek, M. New species of Diatrichalus (Coleoptera: Lycidae) from New Guinea and the Moluccas. Zootaxa 2017, 4247, 577–584. [Google Scholar] [CrossRef]
- Bocek, M.; Adamkova, K. New species of Moluccan trichaline net-winged beetles, with remarks on the phylogenetic position and distribution of Schizotrichalus (Coleoptera: Lycidae: Metriorrhynchinae). Zootaxa 2019, 4623, 341–350. [Google Scholar] [CrossRef]
- Bocek, M.; Bocak, L. The molecular phylogeny and classification of trichaline net-winged beetles (Lycidae: Metriorrhynchini). PeerJ 2017, 5, e3963. [Google Scholar] [CrossRef] [PubMed]
- Bocek, M.; Motyka, M.; Kusy, D.; Bocak, L. Kusy Genomic and mitochondrial data identify different species boundaries in aposematically polymorphic eniclases net-winged beetles (Coleoptera: Lycidae). Insects 2019, 10, 295. [Google Scholar] [CrossRef]
- Kusy, D. A new species of Synchonnus (Coleoptera: Lycidae) from New Guinea, with an identification key to the Papuan species. Acta Entomol. Musei Natl. Pragae 2017, 57, 153–160. [Google Scholar] [CrossRef]
- Kleine, R. Bestimmungstabelle der Trichalusverwandtschaft. Treubia 1930, 9, 325–340. [Google Scholar]
- Riedel, A.; Narakusumo, R.P. One hundred and three new species of Trigonopterus weevils from Sulawesi. ZooKeys 2019, 828, 1–153. [Google Scholar] [CrossRef]
- Riedel, A.; Sagata, K.; Surbakti, S.; Tänzler, R.; Balke, M. One hundred and one new species of Trigonopterus weevils from New Guinea. ZooKeys 2013, 280, 1–150. [Google Scholar] [CrossRef] [PubMed]
- Riedel, A.; Tänzler, R.; Balke, M.; Rahmadi, C.; Suhardjono, Y.R. Ninety-eight new species of Trigonopterus weevils from Sundaland and the Lesser Sunda Islands. ZooKeys 2014, 467, 1–162. [Google Scholar] [CrossRef]
- Lohman, D.J.; De Bruyn, M.; Page, T.; Von Rintelen, K.; Hall, R.; Ng, P.K.; Shih, H.-T.; Carvalho, G.; Von Rintelen, T. Biogeography of the Indo-Australian Archipelago. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 205–226. [Google Scholar] [CrossRef]
- Tänzler, R.; Van Dam, M.H.; Toussaint, E.F.A.; Suhardjono, Y.R.; Balke, M.; Riedel, A. Macroevolution of hyperdiverse flightless beetles reflects the complex geological history of the Sunda Arc. Sci. Rep. 2016, 6, 18793. [Google Scholar] [CrossRef]
- Toussaint, E.F.A.; Tänzler, R.; Rahmadi, C.; Balke, M.; Riedel, A. Biogeography of Australasian flightless weevils (Curculionidae, Celeuthetini) suggests permeability of Lydekker’s and Wallace’s Lines. Zool. Scr. 2015, 44, 632–644. [Google Scholar] [CrossRef]
- Letsch, H.; Balke, M.; Toussaint, E.F.A.; Narakusumo, R.P.; Fiedler, K.; Riedel, A. Transgressing Wallace’s Line brings hyperdiverse weevils down to earth. Ecography 2020, 43, 1329–1340. [Google Scholar] [CrossRef]
- Bocak, L.; Matsuda, K.; Yagi, T. A revision of Metriorrhynchus from the Philippines with molecular evidence of an Australian origin of the Oriental Metriorrhynchus fauna (Coleoptera: Lycidae). Eur. J. Entomol. 2006, 103, 115–126. [Google Scholar] [CrossRef]
- Bocak, L.; Yagi, T. Evolution of mimicry patterns in Metriorrhynchus (Coleoptera: Lycidae): The history of dispersal and speciation in South East Asia. Evolution 2010, 64, 39–52. [Google Scholar] [CrossRef]
- Kubecek, V.; Dvorak, M.; Bocak, L. The phylogenetic structure of Metriorrhynchini fauna of Sulawesi, (Coleoptera: Lycidae) with description of a new genus. Zool. Stud. 2011, 50, 645–656. [Google Scholar]
- Li, Y.; Gunter, N.L.; Pang, H.; Bocak, L. DNA-based species delimitation separates highly divergent populations within morphologically coherent clades of poorly dispersing beetles. Zool. J. Linn. Soc. 2015, 175, 59–72. [Google Scholar] [CrossRef]
- Sklenarova, K.; Chesters, U.; Bocak, L. Phylogeography of poorly dispersing net-winged beetles: A role of drifting India in the origin of afrotropical and oriental fauna. PLoS ONE 2013, 8, e67957. [Google Scholar] [CrossRef] [PubMed]
- Bocek, M.; Bocak, L. The origins and dispersal history of the trichaline net-winged beetles in South East Asia, Wallacea, New Guinea and Australia. Zool. J. Linn. Soc. 2019, 185, 1079–1094. [Google Scholar] [CrossRef]
- Gemminger, M.; Harold, E. Catalogus Coleopterorum Hucusque Descriptorum Synonymicus et Systematicus, Autoribus Dr. Gemminger et B. de Harold; E. H. Gummi: Monachii, Germania, 1868. [Google Scholar]
- Pic, M. Contribution a l’étude des Lycides. L’Echange, Rev. Linn. 1922, 407–410, 13–28. [Google Scholar]
- Kazantsev, S.V. New taxa and a checklist of Afrotropical Metriorrhynchini (Coleoptera: Lycidae), with a note on biogeography of the tribe. Russ. Entomol. J. 2012, 21, 23–33. [Google Scholar] [CrossRef]
- Kazantsev, S.V. New Taxa of Papuan net-winged beetles (Lycidae, Coleoptera). Latv. Entomol. 2010, 48, 92–100. [Google Scholar]
- Motyka, M.; Masek, M.; Bocak, L. Congruence between morphology and molecular phylogeny: The reclassification of Calochromini (Coleoptera: Lycidae) and their dispersal history. Zool. J. Linn. Soc. 2017, 180, 47–65. [Google Scholar]
- Kusy, D.; Motyka, M.; Fusek, L.; Li, Y.; Bocek, M.; Bilkova, R.; Ruskova, M.; Bocak, L. Sexually dimorphic characters and shared aposematic patterns mislead the morphology-based classification of the Lycini (Coleoptera: Lycidae). Zool. J. Linn. Soc. 2020, 26. in press. [Google Scholar]
- Hall, R. Cenozoic geological and plate tectonic evolution of SE Asia and the SW Pacific: Computer-based reconstructions, model and animations. J. Asian Earth Sci. 2002, 20, 353–431. [Google Scholar] [CrossRef]
- Bocak, L. A review of the genus Microtrichalus Pic from Sumatra, with notes on Oriental and Australian species (Coleoptera: Lycidae). Acta Soc. Zool. Bohem. 1999, 64, 3–16. [Google Scholar]
- Yokoyama, Y.; Lambeck, K.; De Deckker, P.; Johnston, P.; Fifield, L.K. Timing of the Last Glacial Maximum from observed sea-level minima. Nat. Cell Biol. 2000, 406, 713–716. [Google Scholar] [CrossRef] [PubMed]
- Bowler, J.; Hope, G.; Jennings, J.; Singh, G.; Walker, D. Late quaternary climates of Australia and New Guinea. Quat. Res. 1976, 6, 359–394. [Google Scholar] [CrossRef]
- Bocak, L.; Bocakova, M. Revision of the genus Eniclases Waterhouse, 1879 (Coleoptera, Lycidae, Metriorrhynchinae). Mitt. Münch. Entomol. Gess. 1991, 81, 203–226. [Google Scholar]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 2001. [Google Scholar]
- Kleine, R. Die Lyciden der Philippinen-Inseln. Philipp. J. Sci. 1926, 31, 33–114. [Google Scholar]
- Tseng, H.-Y.; Huang, W.-S.; Jeng, M.-L.; Villanueva, R.J.T.; Nuñeza, O.M.; Lin, C.-P. Complex inter-island colonization and peripatric founder speciation promote diversification of flightless Pachyrhynchus weevils in the Taiwan-Luzon volcanic belt. J. Biogeogr. 2017, 45, 89–100. [Google Scholar] [CrossRef]
- Bray, T.C.; Bocak, L. Slowly dispersing neotenic beetles can speciate on a penny coin and generate space-limited diversity in the tropical mountains. Sci. Rep. 2016, 6, 33579. [Google Scholar] [CrossRef]
- Jiruskova, A.; Bocak, L. Species delimitation in Cautires (Coleoptera: Lycidae) from Peninsular Malaysia using DNA data and morphology. Ann. Zool. 2015, 65, 239–248. [Google Scholar] [CrossRef]
- Jiruskova, A.; Motyka, M.; Bocak, L. High diversity and endemism in the genus Cautires Waterhouse, 1879 (Coleoptera: Lycidae) from the Malay mountain forests, with the descriptions of fourteen new species. Eur. J. Taxon. 2016, 219, 1–29. [Google Scholar] [CrossRef]
- Jiruskova, A.; Motyka, M.; Bocek, M.; Bocak, L. The Malacca Strait separates distinct faunas of poorly-flying Cautires net-winged beetles. PeerJ 2019, 7, e6511. [Google Scholar] [CrossRef]
- Kalousova, R.; Bocak, L. Species delimitation of colour polymorphic Cladophorus (Coleoptera: Lycidae) from New Guinea. Zootaxa 2017, 4320, 505–522. [Google Scholar] [CrossRef]
- Simpson, G.G. Too many lines; the limits of the Oriental and Australian zoogeographic regions. Proc. Am. Philos. Soc. 1977, 121, 107–120. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total | SEA | Phil | Sul | LSun | Mol | NG | Sol | Aus | |
|---|---|---|---|---|---|---|---|---|---|
| Broxylus Waterhouse, 1879 | 11 | 11 * | |||||||
| Cautiromimus Kleine, 1926 | 17 | 1 | 16 # | ||||||
| Cladophorinus Kleine, 1926 | 1 | 1 * | |||||||
| Cladophorus Guérin-Méneville, 1830 | 131 | 1 | 1 | 10 | 111 | 5 | 3 | ||
| Diatrichalus Kleine, 1926 | 49 | 1 | 5 | 1 | 3 | 32 | 4 | 3 | |
| Ditua Waterhouse, 1879 | 12 | 10 | 1 | 1 | |||||
| Eniclases Waterhouse, 1879 | 38 | 2 | 36 # | ||||||
| Falsolucidota Pic, 1921 | 14 | 14 * | |||||||
| Flabellotrichalus Pic, 1921 | 19 | 4 | 12 | 3 | |||||
| Kassemia Bocak, 1998 | 4 | 4 * | |||||||
| Leptotrichalus Kleine, 1925 | 57 | 20 | 28 | 6 | 2 | ||||
| Lobatang Bocak, 1998 | 7 | 1 | 1 | 1 | 4 | ||||
| Malacolycus Kleine, 1943 | 2 | 2 * | |||||||
| Mangkutanus Kubecek et al., 211 | 2 | 2 * | |||||||
| Marena Kazantsev, 2007 | 3 | 3 * | |||||||
| Metriorrhynchoides Kleine, 1926 | 4 | 4 * | |||||||
| Metriorrhynchus Gemm. & Har., 1869 | 194 | 5 | 11 | 7 | 12 | 64 | 8 | 87 | |
| Mimoxylobanus Pic, 1921 | 2 | 1 | 1 | ||||||
| Microtrichalus Pic, 1921 | 43 | 12 | 5 | 1 | 5 | 16 | 3 | 1 | |
| Oriomum Bocak, 1999 | 1 | 1 * | |||||||
| Porrostoma Laporte, 1838 | 26 | 9 | 17 | ||||||
| Procautires Kleine, 1926 | 28 | 1 | 26 | 1 | |||||
| Pseudodontocerus Pic, 1921 | 4 | 4 * | |||||||
| Schizotrichalus Waterhouse, 1879 | 2 | 1 | 1 | ||||||
| Stadenus Waterhouse, 1879 | 8 | 8 * | |||||||
| Sulabanus Dvorak & Bocak, 2007 | 23 | 23 * | |||||||
| Synchonnus Waterhouse, 1879 | 22 | 1 | 5 | 16 | |||||
| Trichalus Waterhouse, 1877 | 65 | 1 | 12 | 18 | 0 | 29 | |||
| Wakarumbia Bocak, 1999 | 31 | 31 * | |||||||
| Xylobanomimus Kleine, 1926 | 1 | 1 * | |||||||
| Xylobanomorphus Kleine, 1935 | 4 | 4 * | |||||||
| Xylothrix Kazantsev, 2015 | 2 | 2 * | |||||||
| ‘Xylobanus’ (species incertae sedis) | 50 | 1 | 22 | 27 | |||||
| Total | 876 | 43 | 51 | 84 | 5 | 54 | 423 | 21 | 196 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bocak, L.; Motyka, M.; Kusy, D.; Bilkova, R. Biodiversity Inventory and Distribution of Metriorrhynchina Net-Winged Beetles (Coleoptera: Lycidae), with the Identification of Generic Ranges. Insects 2020, 11, 710. https://doi.org/10.3390/insects11100710
Bocak L, Motyka M, Kusy D, Bilkova R. Biodiversity Inventory and Distribution of Metriorrhynchina Net-Winged Beetles (Coleoptera: Lycidae), with the Identification of Generic Ranges. Insects. 2020; 11(10):710. https://doi.org/10.3390/insects11100710
Chicago/Turabian StyleBocak, Ladislav, Michal Motyka, Dominik Kusy, and Renata Bilkova. 2020. "Biodiversity Inventory and Distribution of Metriorrhynchina Net-Winged Beetles (Coleoptera: Lycidae), with the Identification of Generic Ranges" Insects 11, no. 10: 710. https://doi.org/10.3390/insects11100710
APA StyleBocak, L., Motyka, M., Kusy, D., & Bilkova, R. (2020). Biodiversity Inventory and Distribution of Metriorrhynchina Net-Winged Beetles (Coleoptera: Lycidae), with the Identification of Generic Ranges. Insects, 11(10), 710. https://doi.org/10.3390/insects11100710