Locust Bacterial Symbionts: An Update




{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
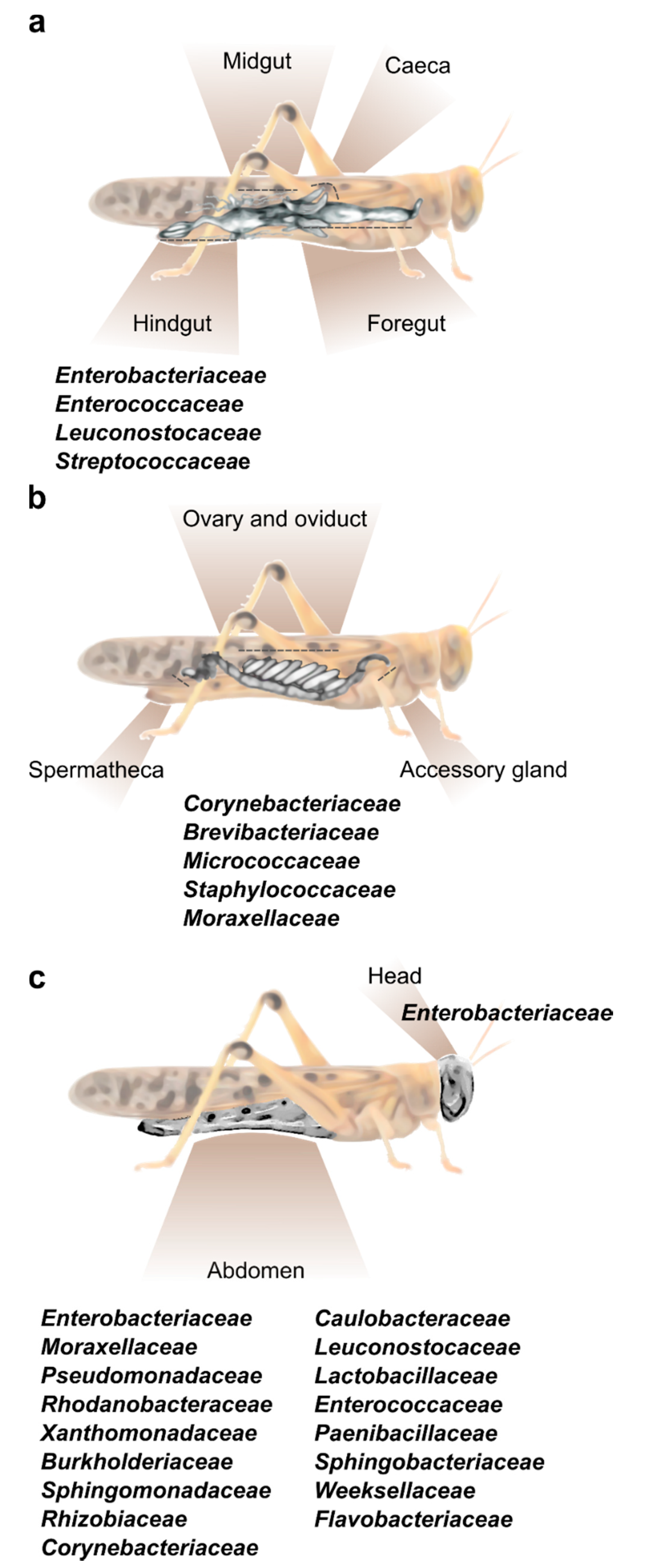
2. Locust Bacterial Symbionts and Their Location
2.1. The Digestive Tract
2.2. The Female Reproductive System
2.3. The Cuticle
3. Locust–Bacteria Mutualism
4. Transgenerational Transmission of Bacterial Symbionts
5. Bacterial Symbionts and Density-Dependent Phase Polyphenism
6. Concluding Remarks and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Pener, M.P.; Simpson, S.J. Locust phase polyphenism: An update. Adv. Insect Phys. 2009, 36, 1–272. [Google Scholar]
- Cullen, D.A.; Cease, A.J.; Latchininsky, A.V.; Ayali, A.; Berry, K.; Buhl, J.; De Keyser, R.; Foquet, B.; Hadrich, J.C.; Matheson, T.; et al. From Molecules to Management: Mechanisms and Consequences of Locust Phase Polyphenism, 1st ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2017; Volume 53. [Google Scholar]
- Ayali, A. The puzzle of locust density-dependent phase polyphenism. Curr. Opin. Insect Sci. 2019, 35, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Bucher, G.; Stephens, J.M. Bacteria of grasshoppers of Western Canada. I. The Enterobacteriaceae. J. Insect Pathol. 1959, 1, 356–373. [Google Scholar]
- Bucher, G.; Stephens, J.M. Bacteria of grasshoppers of Western Canada. II. The Pseudomonadaceae, Achromobacteraceae, Micrococcaceae, Brevibacteriaceae, Lactobacillaceae, and less important families. J. Insect Pathol. 1959, 1, 374–390. [Google Scholar]
- Bucher, G. Bacteria of grasshoppers of Western Canada. III. Frequency of occurrence, pathogenicity. J. Insect Pathol. 1959, 1, 391–405. [Google Scholar]
- Stevenson, J.P. The Normal of Bacterial of the the Alimentary Desert Forskiil Locust. J. Invertebr. Pathol. 1966, 211, 205–211. [Google Scholar] [CrossRef]
- Hunt, J.; Charnley, A.K. Abundance and distribution of the gut flora of the desert locust, Schistocerca gregaria. J. Invertebr. Pathol. 1981, 38, 378–385. [Google Scholar] [CrossRef]
- Mead, L.J.; Khachatourians, G.G.; Jones, G. A Microbial Ecology of the Gut in Laboratory Stocks of the Migratory Grasshopper, Melanoplus sanguinipes (Fab.) (Orthoptera: Acrididae). Appl. Environ. Microbiol. 1988, 54, 1174–1181. [Google Scholar] [CrossRef] [PubMed]
- Dillon, R.; Charnley, A. Mutualism between the desert locust Schistocerca gregaria and its gut microbiota. Res. Microbiol. 2002, 153, 503–509. [Google Scholar] [CrossRef]
- Dillon, R.J.; Vennard, C.T.; Charnley, A.K. A Note: Gut bacteria produce components of a locust cohesion pheromone. J. Appl. Microbiol. 2002, 92, 759–763. [Google Scholar] [CrossRef]
- Dillon, R.J.; Vennard, C.T.; Buckling, A.; Charnley, A.K. Diversity of locust gut bacteria protects against pathogen invasion. Ecol. Lett. 2005, 8, 1291–1298. [Google Scholar] [CrossRef]
- Dillon, R.J.; Webster, G.; Weightman, A.J.; Dillon, V.M.; Blanford, S.; Charnley, A.K. Composition of Acridid gut bacterial communities as revealed by 16S rRNA gene analysis. J. Invertebr. Pathol. 2008, 97, 265–272. [Google Scholar] [CrossRef]
- Shi, W.; Guo, Y.; Xu, C.; Tan, S.; Miao, J.; Feng, Y.; Zhao, H.; St Leger, R.J.; Fang, W. Unveiling the mechanism by which microsporidian parasites prevent locust swarm behavior. Proc. Natl. Acad. Sci. USA 2014, 111, 1343–1348. [Google Scholar] [CrossRef] [PubMed]
- Lavy, O.; Gophna, U.; Gefen, E.; Ayali, A. The effect of density-dependent phase on the locust gut bacterial composition. Front. Microbiol. 2019, 10, 1–8. [Google Scholar] [CrossRef]
- Lavy, O.; Gophna, U.; Gefen, E.; Ayali, A. Dynamics of bacterial composition in the locust reproductive tract are affected by the density-dependent phase. FEMS Microbiol. Ecol. 2020, 96, 1–10. [Google Scholar] [CrossRef]
- Lavy, O.; Gophna, U.; Ayali, A.; Gihaz, S.; Fishman, A.; Gefen, E. The maternal foam plug constitutes a reservoir for the desert locust’s bacterial symbionts. bioRxiv 2020. [Google Scholar] [CrossRef]
- Lavy, O.; Lewin-Epstine, O.; Gophna, U.; Gefen, E.; Hadany, L.; Ayali, A. Microbiome-related aspects of locust density-dependent phase transition. bioRxiv 2020. [Google Scholar] [CrossRef]
- Murdoch, C.C.; Rawls, J.F. Commensal Microbiota Regulate Vertebrate Innate Immunity-Insights from the Zebrafish. Front. Immunol. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Douglas, A.E. Multiorganismal Insects: Diversity and Function of Resident Microorganisms. Annu. Rev. Entomol. 2015, 60, 17–34. [Google Scholar] [CrossRef]
- Dillon, R.J.; Webster, G.; Weightman, A.J.; Keith, C. Diversity of gut microbiota increases with aging and starvation in the desert locust. Antonie Leeuwenhoek 2010, 97, 69–77. [Google Scholar] [CrossRef]
- Stoops, J.; Crauwels, S.; Waud, M.; Claes, J.; Lievens, B.; Van Campenhout, L. Microbial community assessment of mealworm larvae (Tenebrio molitor) and grasshoppers (Locusta migratoria migratorioides) sold for human consumption. Food Microbiol. 2016, 53, 122–127. [Google Scholar] [CrossRef]
- Garofalo, C.; Osimani, A.; Milanović, V.; Taccari, M.; Cardinali, F.; Aquilanti, L.; Riolo, P.; Ruschioni, S.; Isidoro, N.; Clementi, F. The microbiota of marketed processed edible insects as revealed by high-throughput sequencing. Food Microbiol. 2017, 62, 15–22. [Google Scholar] [CrossRef]
- Albrecht, F.O. The Anatomy of the Migratory Locust; University of London: London, UK; Athlone Press: London, UK; Western Printing Services Ltd.: London, UK, 1953. [Google Scholar]
- Dillon, R.J.; Charnley, A.K. The Fate of Fungal Spores in the Insect Gut. Fungal Spore Dis. Initiat. Plants Anim. 1991, 129–156. [Google Scholar] [CrossRef]
- Galal, F.H.; Seufi, A.M. Molecular characterization of cultivable bacteria associated with adult Schistocerca gregaria, using 16S rRNA. J. Taibah Univ. Sci. 2020, 3655. [Google Scholar] [CrossRef]
- Charnley, A.K.; Hunt, J.; Dillon, R.J. The germ-free culture of desert locusts, Schistocerca gregaria. J. Insect. Physiol 1985, 31, 477–485. [Google Scholar] [CrossRef]
- Furman, O.; Shenhav, L.; Sasson, G.; Kokou, F.; Honig, H.; Jacoby, S.; Hertz, T.; Cordero, O.X.; Halperin, E.; Mizrahi, I. Stochasticity constrained by deterministic effects of diet and age drive rumen microbiome assembly dynamics. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Symmons, P.M.; Cressman, K. Desert Locust Guidelines; Food and Agriculture Organization (FAO): Rome, Italy, 2001; p. 42. [Google Scholar]
- Moran, N.A.; Baumann, P. Bacterial endosymbionts in animals. Curr. Opin. Microbiol. 2000, 3, 270–275. [Google Scholar] [CrossRef]
- Oliver, K.M.; Degnan, P.H.; Burke, G.R.; Moran, N.A. Facultative symbionts in aphids and the horizontal transfer of ecologically important traits. Annu. Rev. Entomol. 2010, 55, 247–266. [Google Scholar] [CrossRef]
- Bennett, G.M.; Moran, N.A. Small, smaller, smallest: The origins and evolution of ancient dual symbioses in a phloem-feeding insect. Genome Biol. Evol. 2013, 5, 1675–1688. [Google Scholar] [CrossRef]
- Salem, H.; Florez, L.; Gerardo, N.; Kaltenpoth, M. An out-of-body experience: The extracellular dimension for the transmission of mutualistic bacteria in insects. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142957. [Google Scholar] [CrossRef]
- Currie, C.R.; Poulsen, M.; Mendenhall, J.; Boomsma, J.J.; Billen, J. Coevolved crypts and exocrine glands support mutualistic bacteria in fungus-growing ants. Science 2006, 311, 81–83. [Google Scholar] [CrossRef]
- Kaltenpoth, M.; Wolfgang, G.; Gudrun, H.; Erhard, S. Symbiotic Bacteria Protect Wasp Larvaefrom Fungal Infestation. Curr. Biol. 2005, 15, 475–479. [Google Scholar] [CrossRef]
- Dillon, R.J.; Charnley, A.K. Inhibition of Metarhizium anisopliae by the gut bacterial flora of the desert locust, Schistocerca gregaria: Evidence for an antifungal toxin. J. Invertebr. Pathol. 1986, 47, 350–360. [Google Scholar] [CrossRef]
- Dillon, R.J.; Charnley, A.K. Chemical barriers to gut infection in the desert locust—In-vivo production of antimicrobial phenols associated with the bacterium Pantoea agglomerans. J. Invertebr. Pathol. 1995, 66, 72–75. [Google Scholar] [CrossRef]
- Dillon, R.J.; Vennard, C.T.; Charnley, A.K. Exploitation of gut bacteria in the locust. Nature 2000, 403, 851–852. [Google Scholar] [CrossRef] [PubMed]
- Chapman, R.F.; Joern, A. (Eds.) Biology of Grasshoppers; Wiley-IEEE: New York, NY, USA, 1990. [Google Scholar]
- Skaf, R.; Popov, G.B.; Roffey, J. The desert locust: An international challenge. Philos. Trans. R. Soc. Lond. B 1990, 328, 525–538. [Google Scholar] [CrossRef]
- Bar-Shmuel, N.; Behar, A.; Segoli, M. What do we know about biological nitrogen fixation in insects? Evidence and implications for the insect and the ecosystem. Insect Sci. 2019, 1–12. [Google Scholar] [CrossRef]
- Wilson, K.; Thomas, M.B.; Blanford, S.; Doggett, M.; Simpson, S.J.; Moore, S.L. Coping with crowds: Density-dependent disease resistance in desert locusts. Proc. Natl. Acad. Sci. USA 2002, 99, 5471–5475. [Google Scholar] [CrossRef]
- Wada-Katsumata, A.; Zurek, L.; Nalyanya, G.; Roelofs, W.L.; Zhang, A.; Schal, C. Gut bacteria mediate aggregation in the German cockroach. Proc. Natl. Acad. Sci. USA 2015, 112, 1504031112. [Google Scholar] [CrossRef]
- Bourtzis, K.; Thomas, A.M. (Eds.) Insect Symbiosis; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Yuval, B. Symbiosis: Gut Bacteria Manipulate Host Behaviour. Curr. Biol. 2017, 27, R746–R747. [Google Scholar] [CrossRef]
- Onchuru, T.O.; Martinez, A.; Ingham, C.S.; Kaltenpoth, M. Transmission of mutualistic bacteria in social and gregarious insects. Curr. Opin. Insect Sci. 2018, 28, 50–58. [Google Scholar] [CrossRef]
- Zhang, L.; Lecoq, M.; Latchininsky, A.; Hunter, D. Locust and Grasshopper Management. Annu. Rev. Entomol. 2019, 64, 15–34. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lavy, O.; Gophna, U.; Gefen, E.; Ayali, A. Locust Bacterial Symbionts: An Update. Insects 2020, 11, 655. https://doi.org/10.3390/insects11100655
Lavy O, Gophna U, Gefen E, Ayali A. Locust Bacterial Symbionts: An Update. Insects. 2020; 11(10):655. https://doi.org/10.3390/insects11100655
Chicago/Turabian StyleLavy, Omer, Uri Gophna, Eran Gefen, and Amir Ayali. 2020. "Locust Bacterial Symbionts: An Update" Insects 11, no. 10: 655. https://doi.org/10.3390/insects11100655
APA StyleLavy, O., Gophna, U., Gefen, E., & Ayali, A. (2020). Locust Bacterial Symbionts: An Update. Insects, 11(10), 655. https://doi.org/10.3390/insects11100655