Estimation of the Potential Infestation Area of Newly-invaded Fall Armyworm Spodoptera Frugiperda in the Yangtze River Valley of China

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
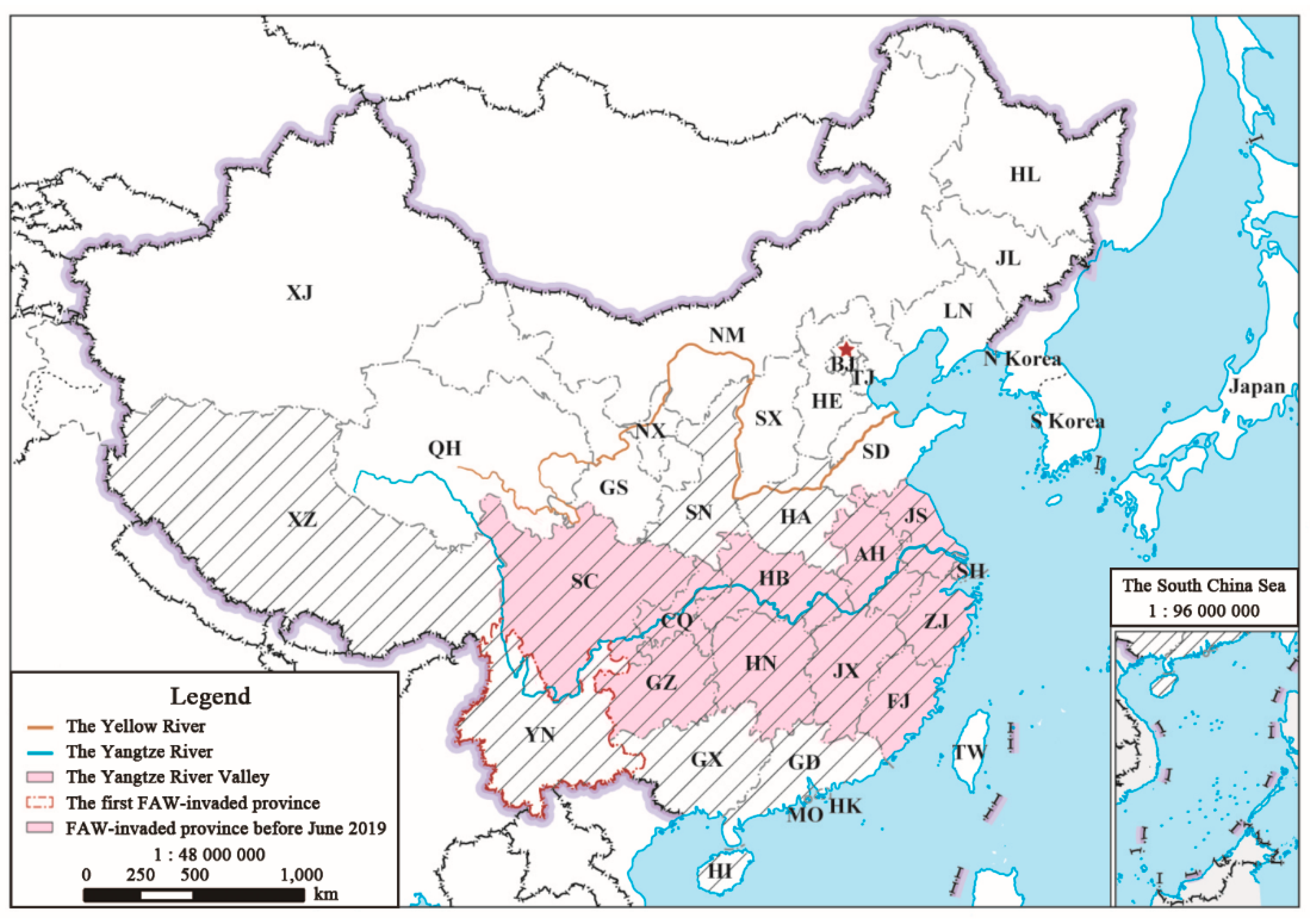
2.1. Study Area and Field Investigations of Invasive FAW before June 2019 in the Yangtze River Valley
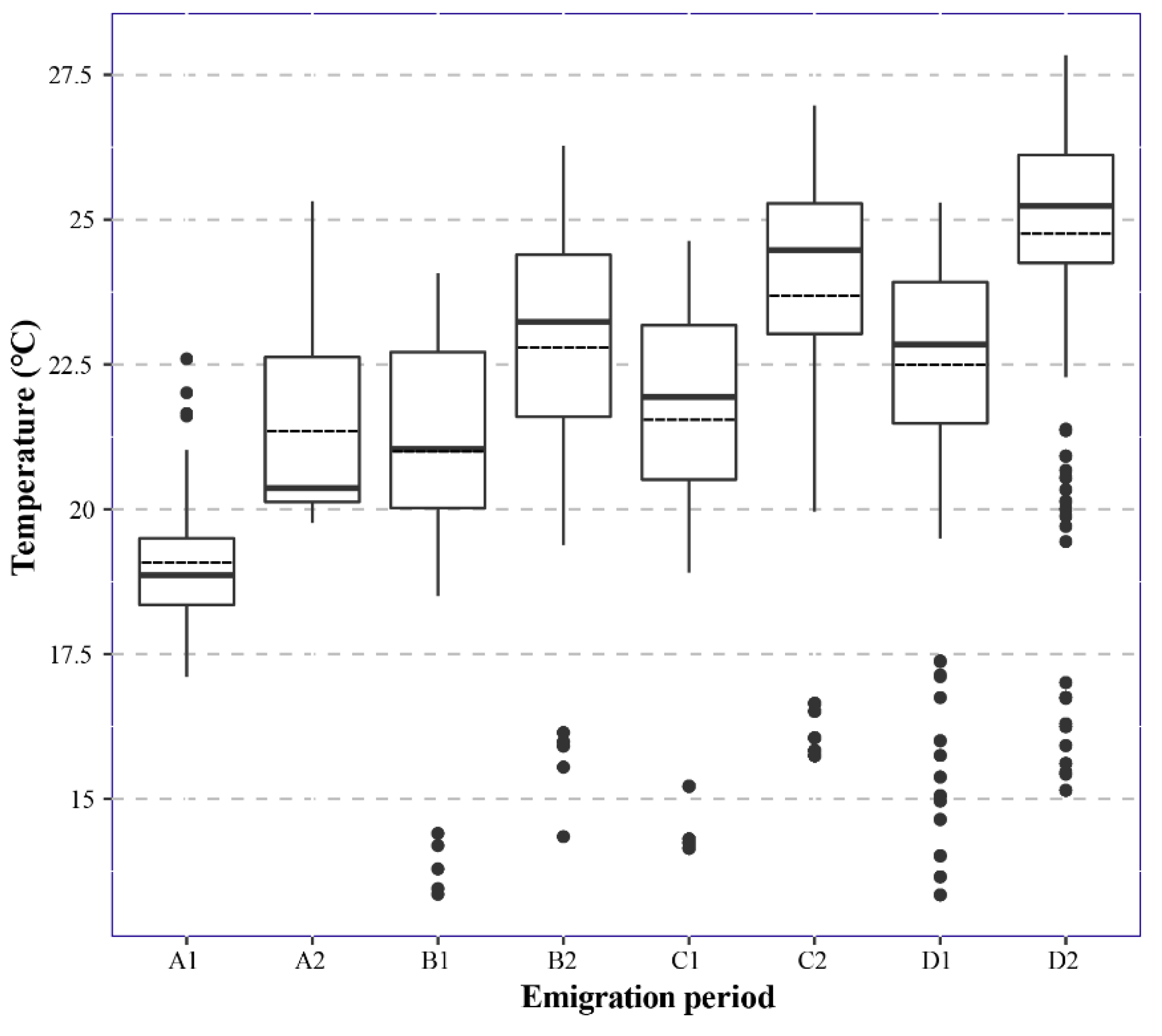
2.2. Estimations of Emigration Periods of the First and Second FAW Generations of Newly-invaded Populations
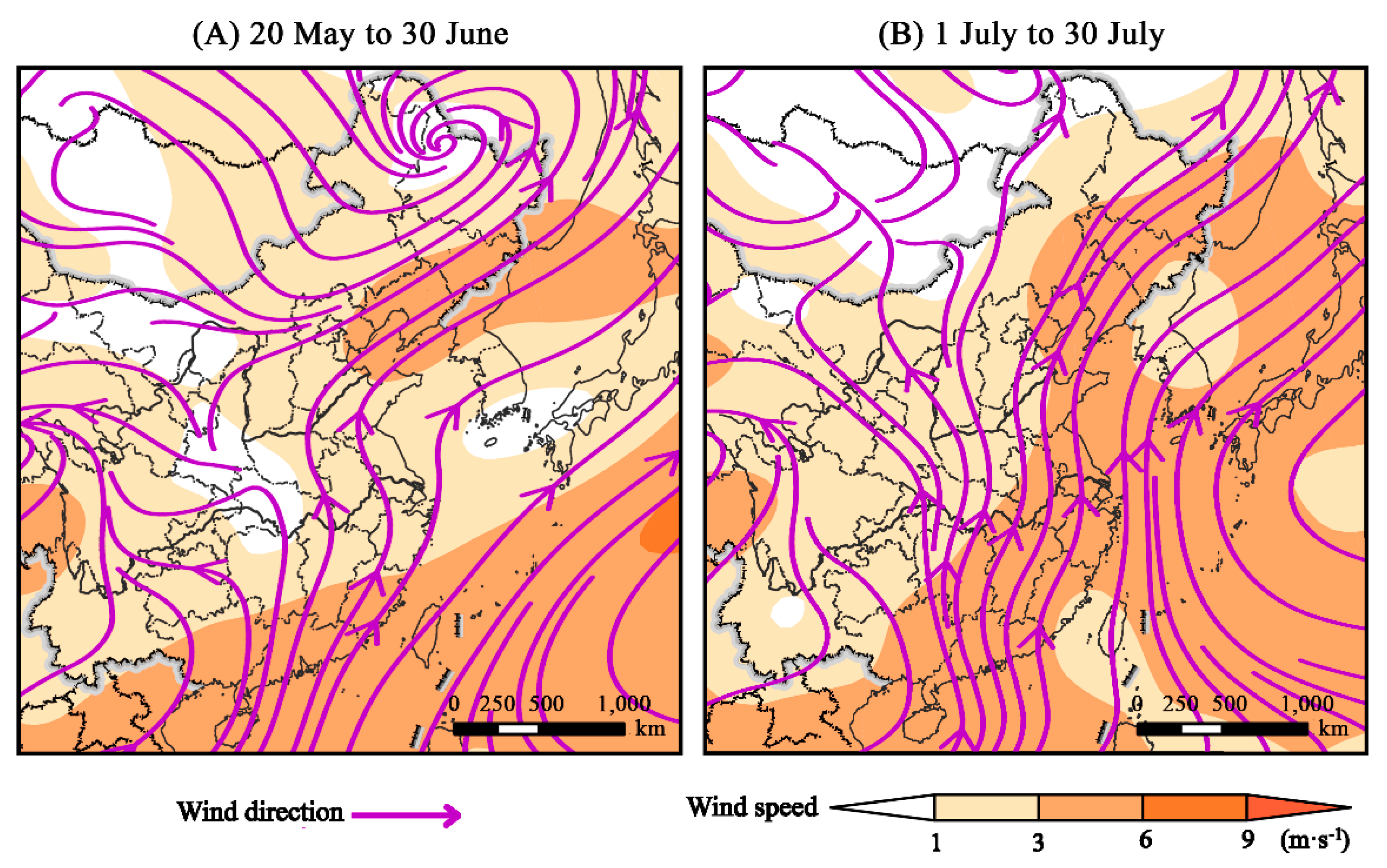
2.3. Analysis of Migratory Flight Relative to Atmospheric Support
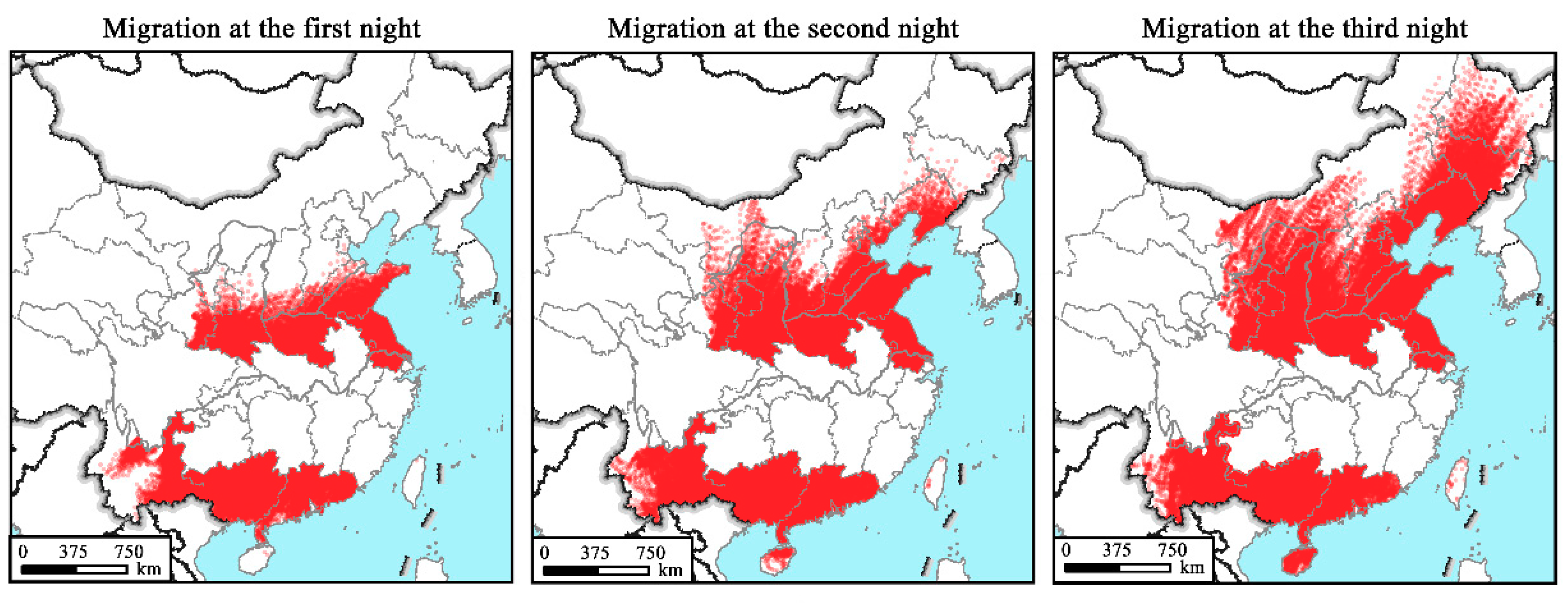
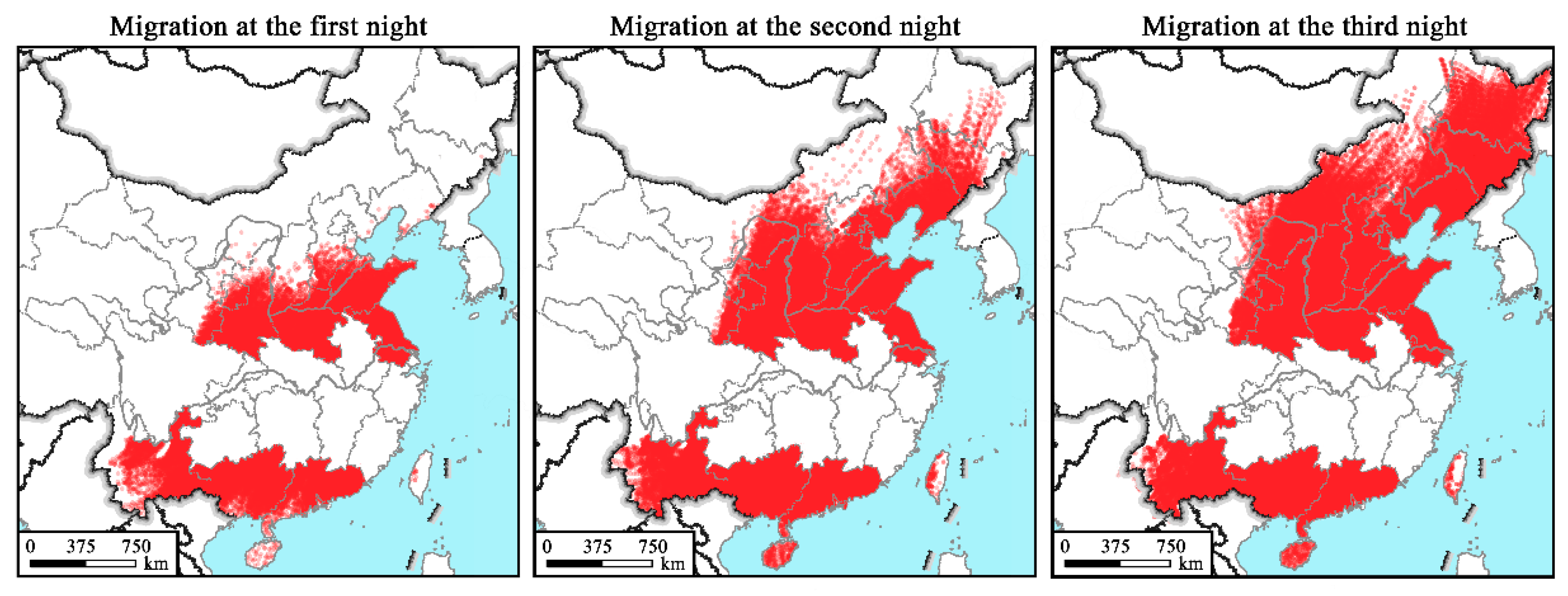
2.4. Numerical Trajectory Modelling Method
3. Results
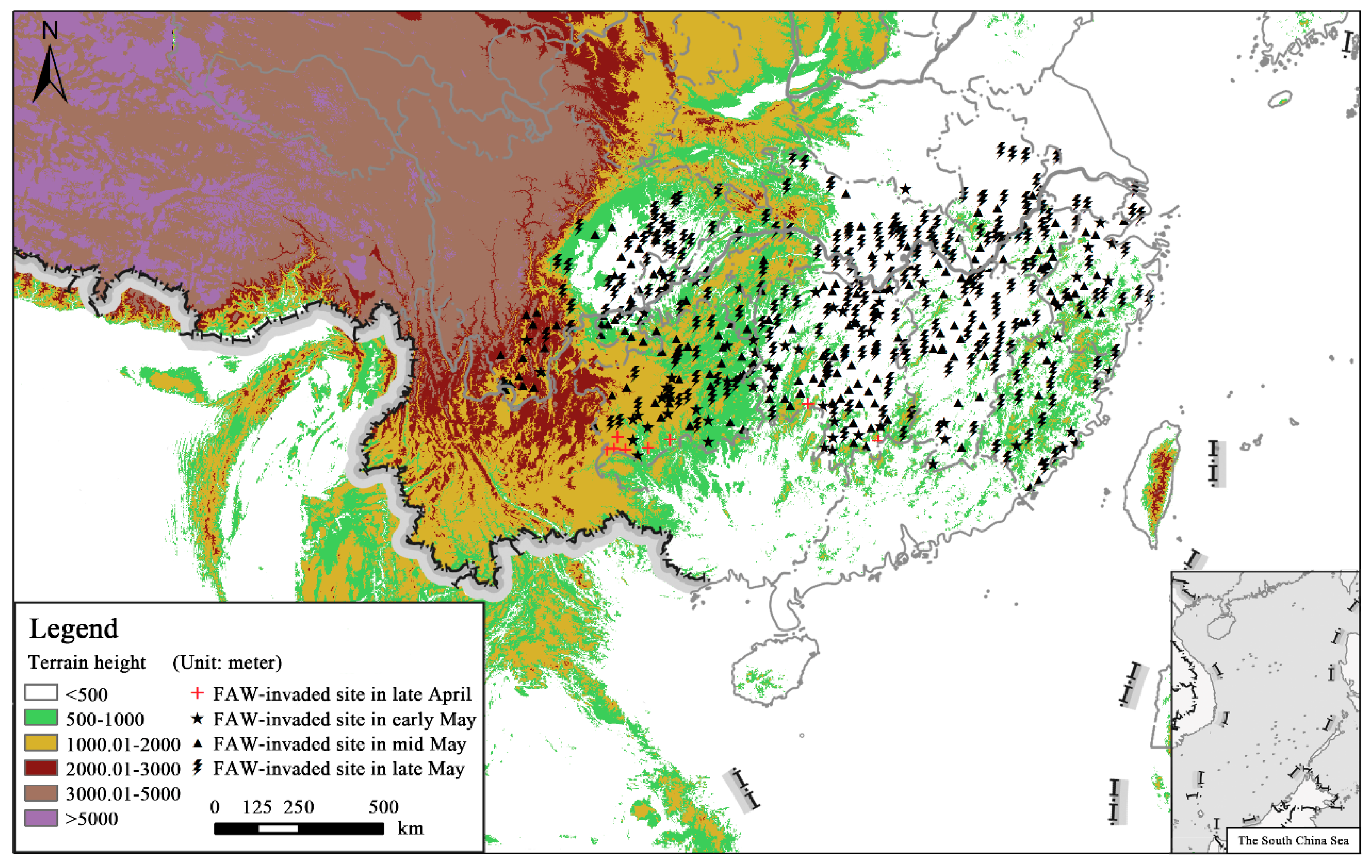
3.1. Large-scale Invasions of Immigratory FAW in the Yangtze River Valley
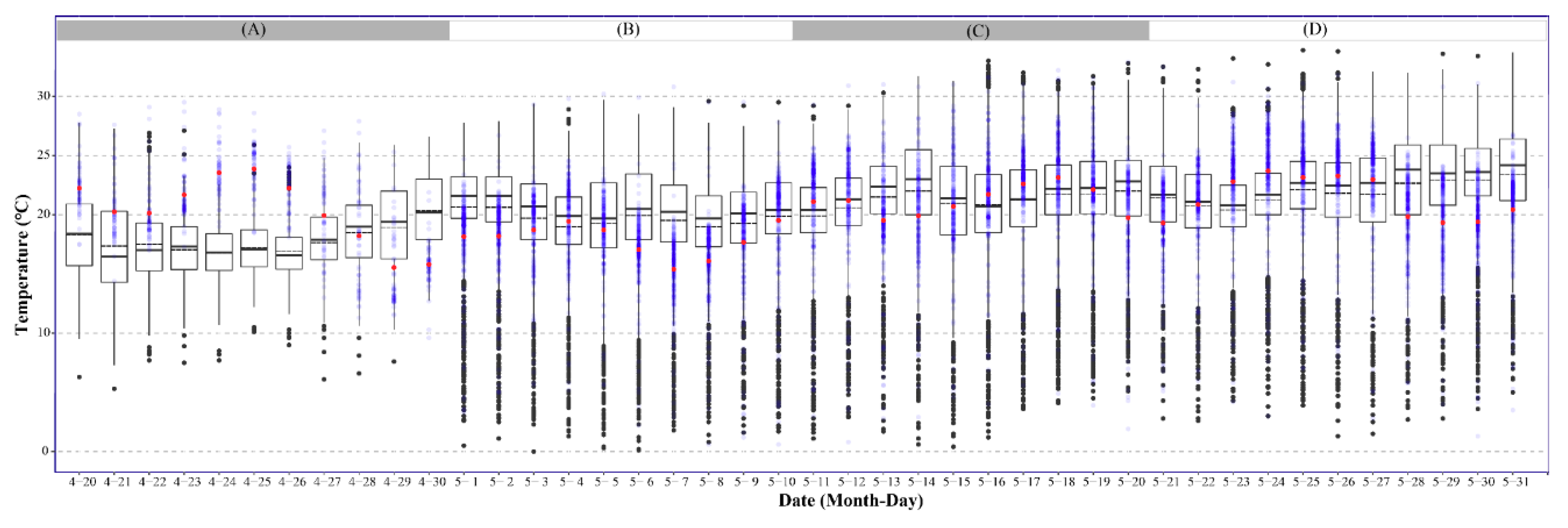
3.2. Temperature Dynamics in the Yangtze River Valley During the FAW Invasion Period
3.3. Migration Arena
3.4. Simulated Flight Trajectories, Landing Sites and Distribution Regions
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Luginbill, P. The fall army worm. USDA Technol. Bull. 1928, 34, 91. [Google Scholar]
- Sparks, A.N. A review of the biology of the fall armyworm. Fla. Entomol. 1979, 62, 82–86. [Google Scholar] [CrossRef]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef] [PubMed]
- Sharanabasappa; Kalleshwaraswamy, C.M.; Asokan, R.; Swamy, H.M.M.; Maruthi, M.S.; Pavithra, H.B.; Hegde, K.; Navi, S.; Prabhu, S.T.; Goergen, G. First report of the fall armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae), an alien invasive pest on maize in India. Pest Manag. Hortic. Ecosyst. 2018, 24, 23–29. [Google Scholar]
- FAO. Five Things to Know about African Farmers’ Latest Foe-The Fall Armyworm. Available online: http://www.fao.org/fao-stories/article/en/c/1104446/ (accessed on 22 March 2018).
- Vickery, R.A. Studies on the Fall Army Worm in the Gulf Coast District of Texas. USDA Technol. Bull. 1929, 138, 1–64. [Google Scholar]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef]
- Pashley, D.P. Host-associated genetic differentiation in fall armyworm (Lepidoptera: Noctuidae): A sibling species complex? Ann. Entomol. Soc. Am. 1986, 79, 898–904. [Google Scholar] [CrossRef]
- Pashley, D.P.; Martin, J.A. Reproductive incompatibility between host strains of the fall armyworm (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 1987, 80, 731–733. [Google Scholar] [CrossRef]
- Johnson, S.J. Migration and the life history strategy of the fall armyworm, Spodoptera frugiperda in the Western Hemisphere. Int. J. Trop. Insect Sci. 1987, 8, 543–549. [Google Scholar] [CrossRef]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall armyworm: Impacts and implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Rose, A.H.; Silversides, R.H.; Lindquist, O.H. Migration flight by an aphid, Rhopalosiphum maidis (Hemiptera: Aphididae), and a noctuid, Spodoptera frugiperda (Lepidoptera: Noctuidae). Can. Entomol. 1975, 107, 567–576. [Google Scholar] [CrossRef]
- Mitchell, E.R.; McNeil, J.N.; Westbrook, J.K.; Silvain, J.F.; Lalanne-Cassou, B.; Chalfant, R.B.; Pair, S.D.; Waddil, D.; Sotomayor-Rios, A.; Proshold, F.I. Seasonal periodicity of fall armyworm, (Lepidoptera: Noctuidae) in the Caribbean basin and northward to Canada. J. Entomol. Sci. 1991, 26, 39–50. [Google Scholar] [CrossRef]
- Pair, S.D.; Raulston, J.R.; Rummel, D.R.; Westbrook, J.K.; Wolf, W.W.; Sparks, A.N.; Schuster, M.F. Development and production of corn earworm and fall armyworm in the Texas high plains: Evidence for reverse fall migration. Southwest. Entomol. 1987, 12, 89–99. [Google Scholar]
- Mitchell, E.R. Monitoring adult populations of the fall armyworm. Fla. Entomol. 1979, 62, 91–98. [Google Scholar] [CrossRef]
- Pair, S.D.; Raulston, J.R.; Westbrook, J.K.; Wolf, W.W.; Adams, S.D. Fall armyworm (Lepidoptera: Noctuidae) outbreak originating in the lower Rio Grande Valley, 1989. Fla. Entomol. 1991, 74, 200–213. [Google Scholar] [CrossRef]
- Wolf, W.W.; Westbrook, J.K.; Raulston, J.R.; Pair, S.D.; Lingren, P.D. Radar observations of orientation of noctuids migrating from corn fields in the Lower Rio Grande Valley. Southwest. Entomol. 1995, 18, 45–61. [Google Scholar]
- Westbrook, J.K.; Nagoshi, R.N.; Meagher, R.L.; Fleischer, S.J.; Jairam, S. Modeling seasonal migration of fall armyworm moths. Int. J. Biometeorol. 2016, 60, 255–267. [Google Scholar] [CrossRef]
- Cock, M.J.W.; Beseh, P.K.; Buddie, A.G.; Cafá, G.; Crozier, J. Molecular methods to detect Spodoptera frugiperda in Ghana, and implications for monitoring the spread of invasive species in developing countries. Sci. Rep. 2017, 7, 4103. [Google Scholar] [CrossRef]
- Stokstad, E. New crop pest takes Africa at lightning speed. Science 2017, 356, 473–474. [Google Scholar] [CrossRef]
- FAO. FAW Monitoring & Early Warning System (FAMEWS); FAO: Rome, Italy; Available online: https://app.powerbi.com/view?r=eyJrIjoiMmFlOWQxMjctZjIwYy00MTdlLWJmMDgtMGM1ZWQ5YmZmNDQwIiwidCI6IjJmMDYwNjMyLTg4MDgtNGM5ZS05M2NmLTNmY2JkMWM1YTUxYiIsImMiOjh9&refresh=1&pageName=ReportSection0901c9217ada50684ad0 (accessed on 23 July 2018).
- FAO. First Detection of Fall Army Worm on the Border of Thailand. Available online: https://www.ippc.int/en/countries/thailand/pestreports/2018/12/first-detection-of-fall-army-worm-on-the-border-of-thailand/ (accessed on 19 December 2018).
- FAO. FAO Statement on Fall Armyworm in Sri Lanka. Available online: http://www.fao.org/srilanka/news/detail-events/en/c/1177796/ (accessed on 24 January 2019).
- FAO. Briefing Note on FAO Actions on Fall Armyworm. Rome, Italy: FAO, 1-6.. Available online: http://www.fao.org/3/a-bs183e.pdf (accessed on 5 March 2019).
- FAO. First Detection Report of the Fall Armyworm Spodoptera frugiperda (Lepidoptra: Noctuidae) on Maize in Myanmar. Available online: https://www.ippc.int/en/countries/myanmar/pestreports/2019/01/first-detection-report-of-the-fall-armyworm-spodoptera-frugiperda-lepidoptra-noctuidae-on-maize-in-myanma/ (accessed on 14 January 2019).
- FAO. First Detection of Fall Armyworm in China; FAO: Rome, Italy; Available online: https://www.ippc.int/fr/news/first-detection-of-fall-armyworm-in-china/ (accessed on 29 January 2019).
- Wu, Q.L.; Jiang, Y.Y.; Wu, K.M. Analysis of migration routes of fall armyworm (Spodoptera frugiperda (J.E. Smith)) from Myanmar to China. Plant Prot. 2019, 45, 1–9. [Google Scholar]
- Wu, Q.L.; Jiang, Y.Y.; Hu, G.; Wu, K.M. Analysis on spring and summer migration routes of fall armyworm (Spodoptera frugiperda) from tropical and southern subtropical zones of China. Plant Prot. 2019, 45, 1–9. [Google Scholar]
- Ministry of Agriculture and Rural Affairs of the People’s Republic of China. A Notice on Application of Pesticides in Emergency Control of the Fall Armyworm. Available online: http://www.moa.gov.cn/govpublic/ZZYGLS/201906/t20190605_6316201.htm (accessed on 3 June 2019).
- Early, R.; Gonzalez-Moreno, P.; Murphy, S.T.; Day, R. Forecasting the global extent of invasion of the cereal pest Spodoptera frugiperda, the fall armyworm. NeoBiota 2018, 40, 25–50. [Google Scholar] [CrossRef]
- Lin, C.S.; Chang, J.T. Studies on the regularities of the outbreak of the oriental armyworm (Mythimna separata Walker). Acta Phytophy. Sin. 1964, 3, 93–100. [Google Scholar]
- Wu, K.M.; Guo, Y.Y. Geotype differentiation and regional migratory regularity of Helicoverpa armigera in China. Plant Prot. 2007, 33, 6–11. [Google Scholar]
- Zhang, X.X.; Geng, J.G.; Zhou, W.J. Studies on the regularities of the rice leaf folder Cnaphalocrocis medinalis Guenée in China. J. Nanjing Agric. Univ. 1981, 3, 43–54. [Google Scholar]
- Cheng, X.N.; Chen, R.C.; Xi, X.; Yang, L.M.; Zhu, Z.L.; Wu, J.C.; Qian, R.G.; Yang, J.S. Study on the migrations of brown planthopper Nilaparvata lugens stål. Acta Entomol. Sin. 1979, 22, 1–21. [Google Scholar]
- Zhang, L.; Liu, B.; Jiang, Y.; Liu, J.; Wu, K.; Xiao, Y. Molecular characterization analysis of fall armyworm populations in China. Plant Prot. 2019, 1–9, in press. [Google Scholar]
- Wu, Q.L.; Westbrook, J.K.; Hu, G.; Lu, M.H.; Liu, W.C.; Sword, G.A.; Zhai, B.P. Multiscale analyses on a massive immigration process of Sogatella furcifera (Horváth) in south-central China: Influences of synoptic-scale meteorological conditions and topography. Int. J. Biometeorol. 2018, 62, 1389–1406. [Google Scholar] [CrossRef]
- Riley, J.R.; Reynolds, D.R.; Farmery, M.J. Observations of the flight behaviour of the army worm moth, Spodoptera exempta, at an emergence site using radar and infra-red optical techniques. Ecol. Entomol. 1983, 8, 395–418. [Google Scholar] [CrossRef]
- Rose, D.J.W.; Page, W.W.; Dewhurst, C.F.; Riley, J.R.; Reynolds, D.R.; Pedgley, D.E.; Tucker, M.R. Downwind migration of the African army worm moth, Spodoptera exempta, studied by mark-and-capture and by radar. Ecol. Entomol. 1985, 10, 299–313. [Google Scholar] [CrossRef]
- Skamarock, W.C.; Klemp, J.B.; Dudhia, J.; Gill, D.O.; Barker, D.M.; Duda, M.G.; Huang, X.Y.; Wang, W.; Powers, J.G. A description of the advanced research WRF version 3; NCAR Tech. Note. NCAR/TN-475+STR; Mesoscale and Microscale Meteorology Division, NCAR: Boulder, CO, USA, 2008; pp. 1–125. [Google Scholar]
- Hogg, D.B.; Pitre, H.N.; Anderson, R.E. Assessment of early-season phenology of the fall armyworm (Lepidoptera: Noctuidae) in Mississippi. Environ. Entomol. 1982, 11, 705–710. [Google Scholar] [CrossRef]
- Barfield, C.S.; Ashley, T.R. Effects of corn phenology and temperature on the life cycle of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Fla. Entomol. 1987, 70, 110–116. [Google Scholar] [CrossRef]
- He, L.M.; Ge, S.S.; Chen, Y.C.; Wu, Q.L.; Jiang, Y.Y.; Wu, K.M. The developmental threshold temperature, effective accumulated temperature and prediction model of developmental duration of fall armyworm, Spodoptera frugiperda. Plant Prot. 2019. [Google Scholar] [CrossRef]
- National Centers for Environmental Prediction/National Weather Service/NOAA/U.S. Department of Commerce, 2000: NCEP FNL Operational Model Global Tropospheric Analyses, continuing from July 1999. Computational and Information Systems Laboratory, NCAR: Boulder, CO, USA. Available online: https://doi.org/10.5065/D6M043C6 (accessed on 10 June 2019).
- Wu, Q.L.; Hu, G.; Westbrook, J.K.; Sword, G.A.; Zhai, B.P. An advanced numerical trajectory model tracks a corn earworm moth migration event in Texas, USA. Insects 2018, 9, 115. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Lu, F.; Lu, M.; Liu, W.; Xu, W.; Jiang, X.; Zhai, B.P. The influence of typhoon Khanun on the return migration of Nilaparvata lugens (Stål) in eastern China. PLoS ONE 2013, 8, e57277. [Google Scholar] [CrossRef] [PubMed]
- Westbrook, J.K.; Eyster, R.S.; Wolf, W.W.; Lingren, P.L.; Raulston, J.R. Migration pathways of corn earworm (Lepidoptera: Noctuidae) indicated by tetroon trajectories. Agric. For. Meteorol. 1995, 73, 67–87. [Google Scholar] [CrossRef]
- Westbrook, J.K. Noctuid migration in Texas within the nocturnal aeroecological boundary layer. Integr. Comp. Biol. 2008, 48, 99–106. [Google Scholar] [CrossRef]
- Taylor, P.S.; Shields, E.J. Flight thresholds of the armyworm (Lepidoptera: Noctuidae). Environ. Entomol. 1990, 19, 1410–1417. [Google Scholar] [CrossRef]
- Westbrook, J.K.; Esquivel, J.F.; Lopéz, J.D.; Jones, G.D.; Wolf, W.W.; Raulston, J.R. Validation of bollworm migration across southcentral Texas in 1994-1996. Southwest. Entomol. 1998, 23, 209–220. [Google Scholar]
- Liu, J.; Jiang, Y.Y.; Li, H.; Li, Y.H.; Tai, H.K.; Wang, Z.Y. First report of invasive fall armyworm, Spodoptera frugiperda (Smith), on sugarcane from China. Plant Prot. 2019, 39, 35–36, 66. [Google Scholar]
- Chormule, A.; Shejawal, N.; Sharanabasappa, C.M.; Asokan, R.; Swamy, H.M. First report of the fall Armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae) on sugarcane and other crops from Maharashtra, India. J. Entomol. Zool. Stud. 2019, 7, 114–117. [Google Scholar]
- Kalleshwaraswamy, C.M.; Asokan, R.; Mahadevaswamy, H.M.M. First record of invasive fall armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) on rice (Oryza sativa) from India. J. Entomol. Zool. Stud. 2019, 7, 332–337. [Google Scholar]
- Liu, H.; Lan, T.; Fang, D.; Gui, F.; Wang, H.; Guo, W.; Cheng, X.; Chang, Y.; He, S.; Lyu, L.; et al. Chromosome level draft genomes of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), an alien invasive pest in China. bioRxiv 2019. [Google Scholar] [CrossRef]
- Wolf, W.W.; Sparks, A.N.; Pair, S.D.; Westbrook, J.K.; Truesdale, F.M. Radar observations and collections of insects in the Gulf of Mexico. In Insect Flight: Dispersal and Migration; Danthanarayana, W., Ed.; Springer: Berlin/Heidelberg, Germany, 1986; pp. 221–234. [Google Scholar]
- Li, X.J.; Wu, M.F.; Ma, J.; Gao, B.Y.; Wu, Q.L.; Chen, A.D.; Liu, J.; Jiang, Y.Y.; Zhai, B.P.; Early, R.; et al. Prediction of migratory routes of the invasive fall armyworm in eastern China using a trajectory analytical approach. Pest Manag. Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Lu, M.H.; Reynolds, D.R.; Wang, H.; Chen, X.; Liu, W.; Zhu, F.; Wu, X.; Xia, F.; Xie, M.; et al. Long-term seasonal forecasting of a major migrant insect pest: The brown planthopper in the Lower Yangtze River Valley. J. Pest Sci. 2019, 92, 417–428. [Google Scholar] [CrossRef]
- Drake, V.A.; Farrow, R.A. The influence of atmospheric structure and motions on insect migration. Annu. Rev. Entomol. 1988, 33, 183–210. [Google Scholar] [CrossRef]
- FAO. Report of First Detection of Fall Armyworm (FAW) in Republic of Korea. Available online: https://www.ippc.int/en/countries/republic-of-korea/pestreports/2019/06/report-of-first-detection-of-fall-armywormfaw-in-republic-of-korea/ (accessed on 21 June 2019).
- IPPC. Report of First Detection of Spodoptera frugipedera—Fall Armyworm (FAW) in Japan; IPPC Official Pest Report, No. JPN-08/6; IPPC: Rome, Italy, 2019; Available online: https://www.ippc.int (accessed on 3 July 2019).
- Farrow, R.A. Detection of transoceanic migration of insects to a remote island in the Coral Sea, Willis Island. Aust. J. Ecol. 1894, 9, 253–272. [Google Scholar] [CrossRef]
- Fox, K.J. The transoceanic migration of Lepidoptera to New Zealand—A history and a hypothesis on colonisation. N. Z. Entomol. 1978, 6, 368–380. [Google Scholar] [CrossRef]
- Feng, H.Q.; Wu, X.F.; Wu, B.; Wu, K.M. Seasonal migration of Helicoverpa armigera (Lepidoptera: Noctuidae) over the Bohai sea. J. Econ. Entomol. 2009, 102, 95–104. [Google Scholar] [CrossRef]
- FAO. Briefing Note on FAP Actions on Fall Armyworm in Africa 15 December 2017. Available online: http://www.fao.org/fileadmin/templates/fcc/map/map_of_affected_areas/Fall_Armyworm_brief_-__15Dec2017_.pdf (accessed on 15 December 2017).

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Q.-L.; He, L.-M.; Shen, X.-J.; Jiang, Y.-Y.; Liu, J.; Hu, G.; Wu, K.-M. Estimation of the Potential Infestation Area of Newly-invaded Fall Armyworm Spodoptera Frugiperda in the Yangtze River Valley of China. Insects 2019, 10, 298. https://doi.org/10.3390/insects10090298
Wu Q-L, He L-M, Shen X-J, Jiang Y-Y, Liu J, Hu G, Wu K-M. Estimation of the Potential Infestation Area of Newly-invaded Fall Armyworm Spodoptera Frugiperda in the Yangtze River Valley of China. Insects. 2019; 10(9):298. https://doi.org/10.3390/insects10090298
Chicago/Turabian StyleWu, Qiu-Lin, Li-Mei He, Xiu-Jing Shen, Yu-Ying Jiang, Jie Liu, Gao Hu, and Kong-Ming Wu. 2019. "Estimation of the Potential Infestation Area of Newly-invaded Fall Armyworm Spodoptera Frugiperda in the Yangtze River Valley of China" Insects 10, no. 9: 298. https://doi.org/10.3390/insects10090298
APA StyleWu, Q.-L., He, L.-M., Shen, X.-J., Jiang, Y.-Y., Liu, J., Hu, G., & Wu, K.-M. (2019). Estimation of the Potential Infestation Area of Newly-invaded Fall Armyworm Spodoptera Frugiperda in the Yangtze River Valley of China. Insects, 10(9), 298. https://doi.org/10.3390/insects10090298