Effects of Water Immersion and Soil Moisture Content on Larval and Pupal Survival of Bactrocera minax (Diptera: Tephritidae)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Effects of Water Immersion on B. minax Larvae
2.3. Effect of Soil Moisture Content on the B. minax Pupae
2.4. Statistical Analysis
2.4.1. Water Immersion on the Performance of Mature Larvae
2.4.2. Soil Moisture Content on Pupal Survival
2.4.3. Data Analysis
3. Results
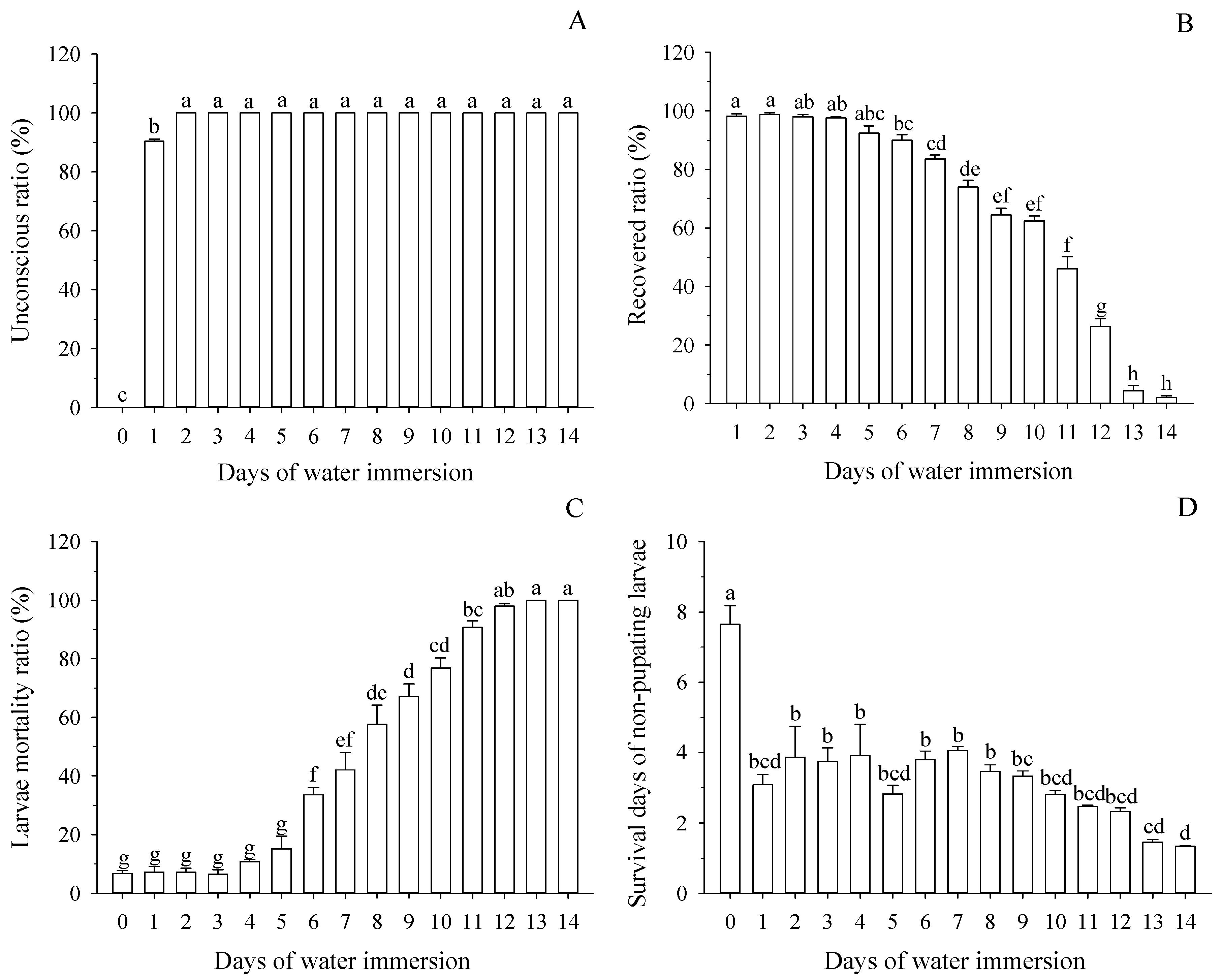
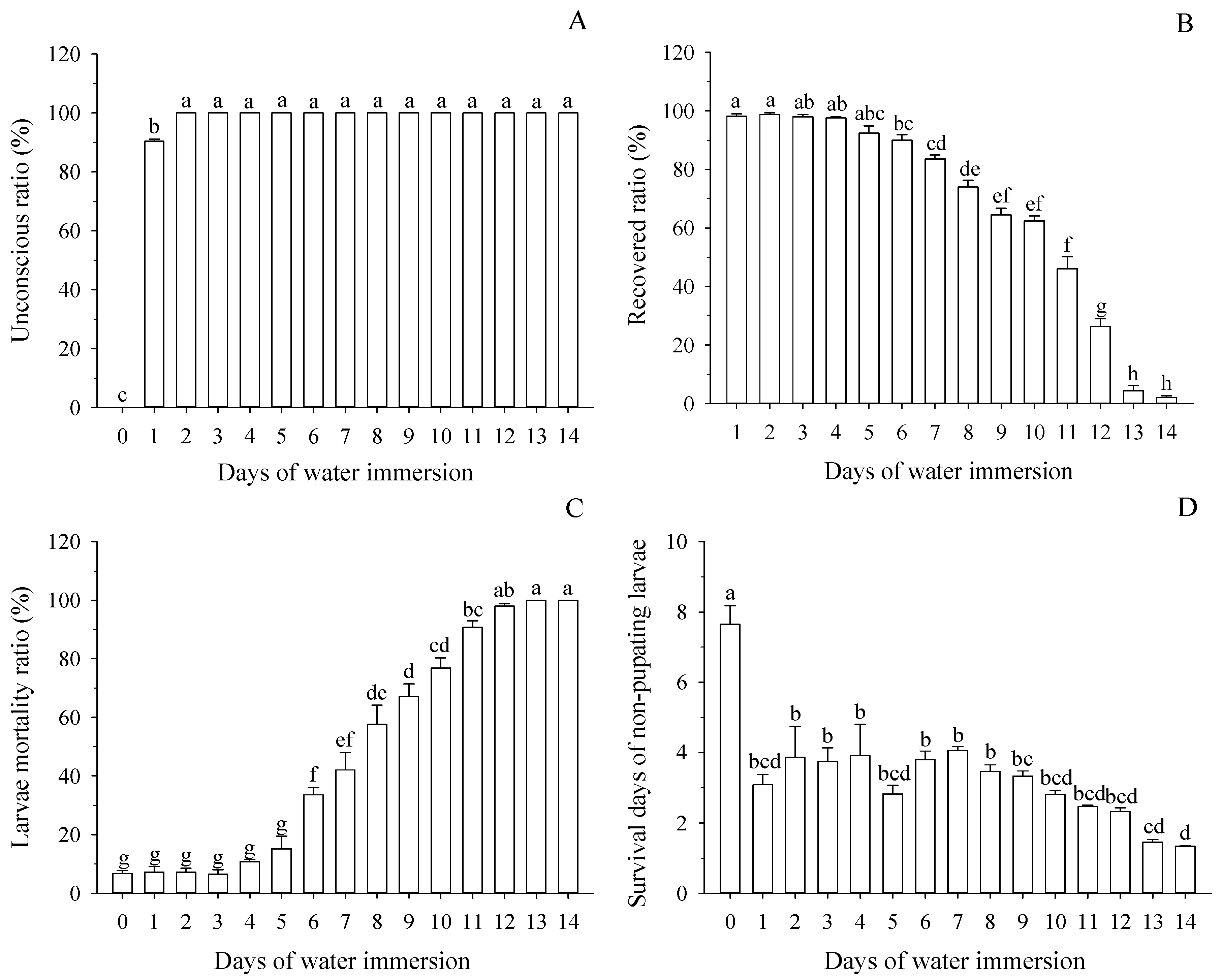
3.1. Effect of Water Immersion on Survival of Mature Larvae
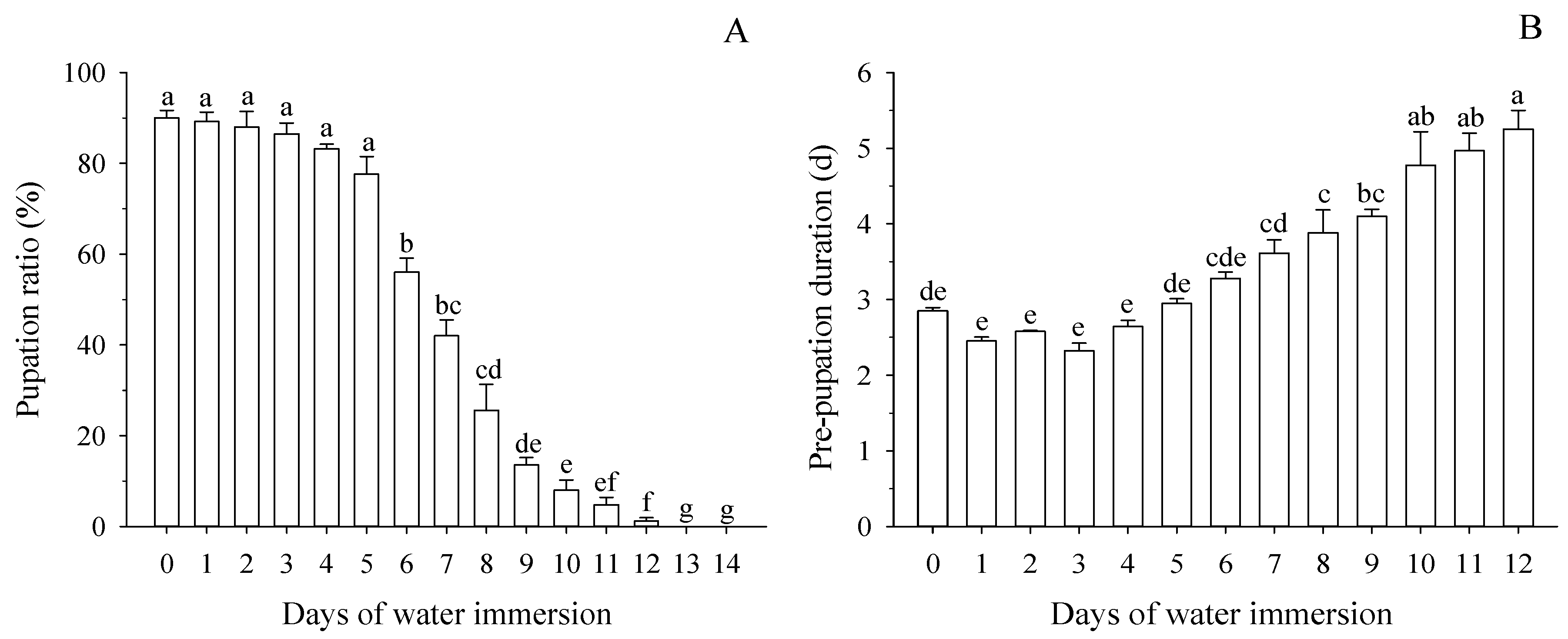
3.2. Effect of Water Immersion on Pupation of Mature Larvae
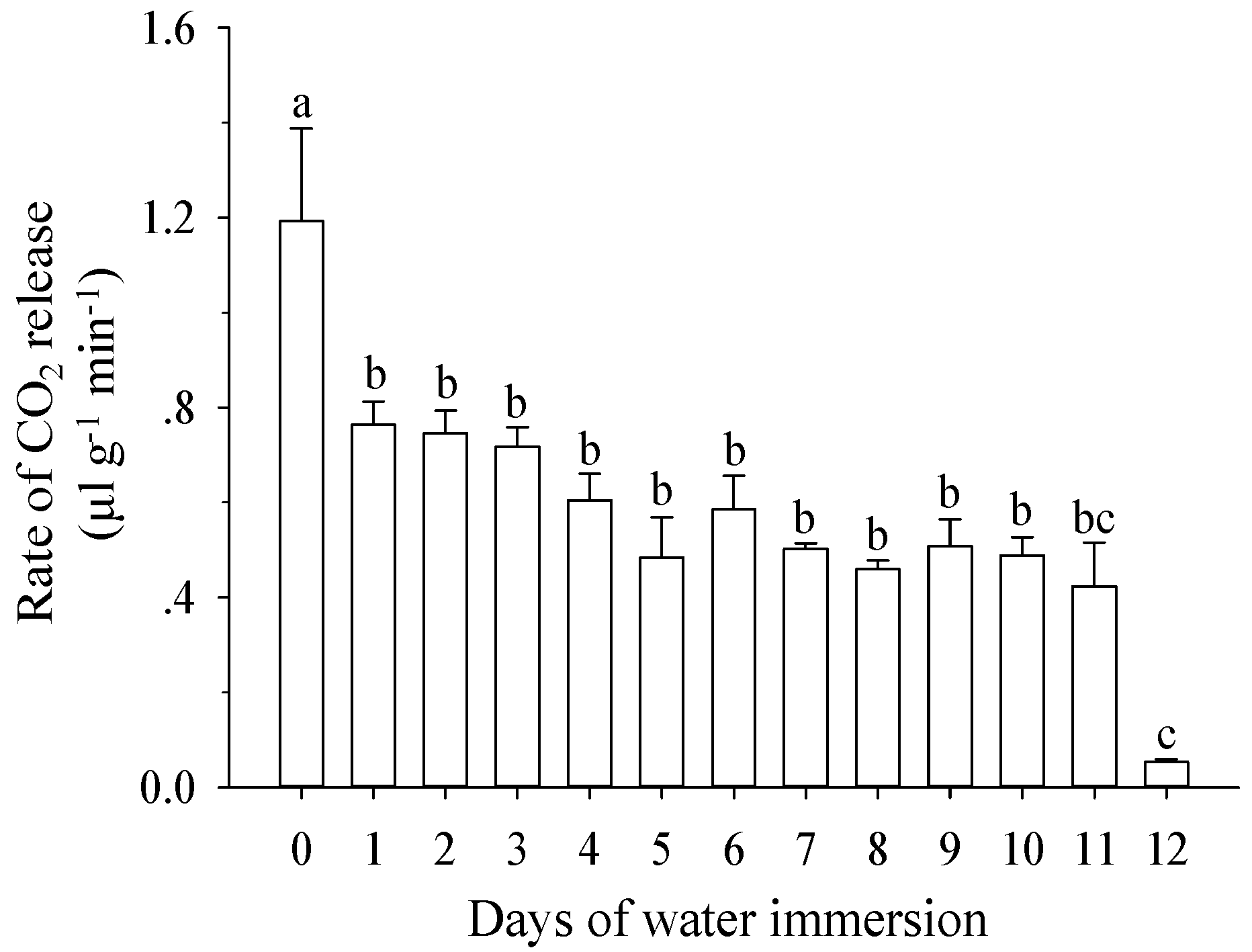
3.3. Effect of Water Immersion on Respiration Rates of Mature Larvae
3.4. Effects of Soil Moisture on Pupal Survival
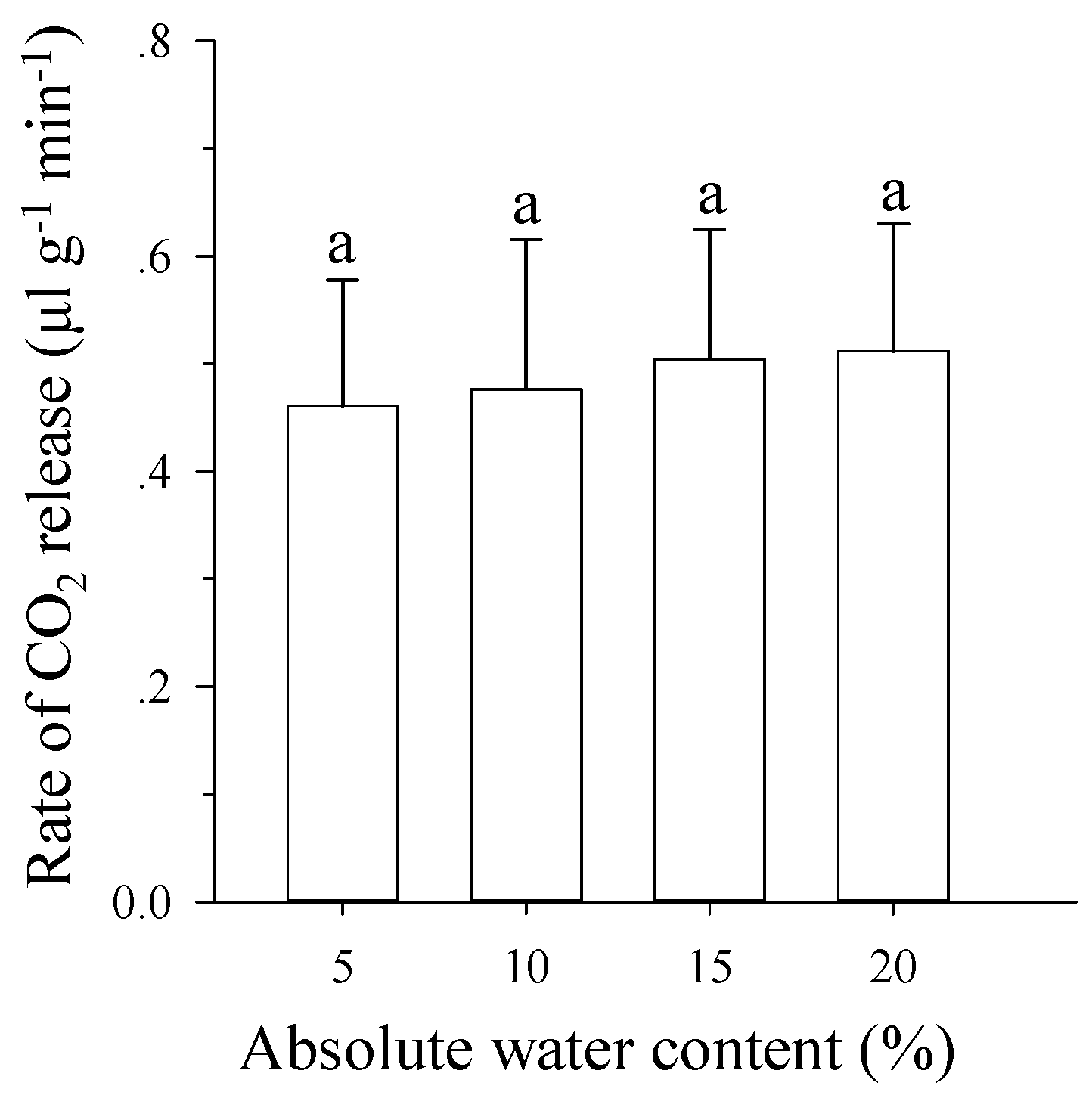
3.5. Effects of Soil Moisture on Respiration Rates of Pupae
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hetz, S.K.; Bradley, T.J. Insects breathe discontinuously to avoid oxygen toxicity. Nature 2005, 433, 516–519. [Google Scholar] [CrossRef]
- Levy, R.I.; Schneiderman, H.A. Discontinuous respiration in insects—II, the direct measurement and significance of changes in tracheal gas composition during the respiratory cycle of silkworm pupae. J. Insect Physiol. 1966, 12, 83–104. [Google Scholar] [CrossRef]
- Lighton, J.R.B. Notes from the underground: Toward ultimate hypotheses of cyclic, discontinuous gas-exchange in tracheate arthropods. Am. Zool. 1998, 38, 483–491. [Google Scholar] [CrossRef]
- Basson, C.H.; Terblanche, J.S. Metabolic responses of Glossina pallidipes (Diptera: Glossinidae) puparia exposed to oxygen and temperature variation: Implications for population dynamics and subterranean life. J. Insect Physiol. 2010, 56, 1789–1797. [Google Scholar] [CrossRef] [PubMed]
- Eskafi, F.M.; Fernandez, A. Larval–pupal mortality of mediterranean fruit fly (Diptera: Tephritidae) from interaction of soil, moisture, and temperature. Environ. Entomol. 1990, 19, 1666–1670. [Google Scholar] [CrossRef]
- Hoback, W.W.; Stanley, D.W.; Higley, L.G.; Barnhart, M.C. Survival of immersion and anoxia by larval tiger beetles, Cicindela togata. Am. Midl. Nat. 1998, 140, 27–33. [Google Scholar] [CrossRef]
- Taschenberg, E.F.; Lopez, F.; Steiner, L.F. Responses of maturing larvae of Anastrepha suspensa to light and to immersion in water. J. Econ. Entomol. 1974, 67, 731–734. [Google Scholar] [CrossRef]
- Yee, W.L. Soil moisture and relative humidity effects during postdiapause on the emergence of western cherry fruit fly (Diptera: Tephritidae). Can. Entomol. 2013, 145, 317–326. [Google Scholar] [CrossRef]
- Quesada-Moraga, E.; Valverde-García, P.; Garrido-Jurado, I. The effect of temperature and soil moisture on the development of the preimaginal mediterranean Fruit Fly (Diptera: Tephritidae). Environ. Entomol. 2012, 41, 966–970. [Google Scholar] [CrossRef]
- Hulthen, A.D.; Clarke, A.R. The influence of soil type and moisture on pupal survival of Bactrocera tryoni (Froggatt) (Diptera: Tephritidae). Aust. J. Entomol. 2006, 45, 16–19. [Google Scholar] [CrossRef]
- Ren, L.; Lu, Y.Y.; Zeng, L. Effect of sand water content on the pupal survival of Bactrocera dorsalis (Hendel). J. South China Agric. Univ. 2007, 28, 63–66. [Google Scholar]
- Wei, C.H.; Zeng, L. Effect of different soil water content on pupa of Bactrocera correcta (Bezzi). Guangdong Agric. Sci. 2011, 9, 65–67. [Google Scholar]
- Zhang, Z.; Li, Z.Z. Effects of the soil relative moisture content and the soil-soaking on pupal development of Liriomyza huidobrensis. J. Mt. Agric. Biol. 2001, 220, 422–425. [Google Scholar]
- Liu, Q.; Niu, C.Y.; Wang, X. Influence of drowning time and soil water content on the pupal eclosion rate of Bactrocera dorsalis (Hendel) of Hubei Province. Plant Quar. 2010, 24, 22–24. [Google Scholar]
- Alyokhin, A.V.; Mille, C.; Messing, R.H.; Duan, J.J. Selection of pupation habitats by oriental fruit fly larvae in the laboratory. J. Insect Behav. 2001, 14, 57–66. [Google Scholar] [CrossRef]
- Luo, L.Y.; Chen, C.F. Biological characteristics of pupa of Tetradacus citri (Chen). South China Fruit. 1987, 4, 9–10. [Google Scholar]
- Wang, X.J.; Luo, L.Y. Research progress in the Chinese citrus fly. Entomol. Knowl. 1995, 32, 310–315. [Google Scholar]
- Dorji, C.; Clarke, A.R.; Drew, R.A.I.; Fletcher, B.S.; Loday, P.; Mahat, K.; Raghu, S.; Romig, M.C. Seasonal phenology of Bactrocera minax (Diptera: Tephritidae) in western Bhutan. Bull. Entomol. Res. 2006, 96, 531–538. [Google Scholar] [PubMed]
- Drew, R.A.I.; Raghu, S. The fruit fly fauna (Diptera: Tephritidae: Dacinae) of the rainforest habitat of the Western Ghats, India. Raffles B Zool. 2002, 50, 327–352. [Google Scholar]
- Yu, S.C.; Wang, Z.Y.; Yang, G.F.; Feng, Y.X. Adult occurrence period of Tetradacus citri (Chen). Entomol. Knowl. 1996, 33, 210–212. [Google Scholar]
- Nie, J.Y.; Zheng, Z.; Song, F.P.; Tan, S.Q.; Yi, J.P. Observation on occurrence regularity and control experiment of Tetradacus citri (Chen) in Zigui County. Plant Prot. Technol. Ext. 1999, 19, 33–34. [Google Scholar]
- Song, Z.Q. Land-based climate types and distribution patterns of horizontal natural zones. J. Inn. Mong. Norm. Univ. (Nat. Sci. Ed.) 1982, 2, 137–143. [Google Scholar]
- Wu, Z.Q. Primary observation on behavior of Chinese fruit fly (Tetradacus crtri Chen). Entomol. Knowl. 1958, 5, 216–217. [Google Scholar]
- Yuan, Y.M.; Chen, X.G. Autumn rainfall features and main influencing meteorological influence system analysis in upper Yangtze River. Yangtze River 2011, 42, 21–24. [Google Scholar]
- Lu, M.Y.; Pan, J.C. It is important to pay attention on the problem of waterlogging in mountainous orchard. South China Fruit. 1998, 27, 36. [Google Scholar]
- Li, L.; Xiao, R.L.; Huang, Y.; Li, H. Change of moisture and management on water in soil from newly established citrus orchards in the Northwest of Guangxi Zhuang Autonomous Region. Agro-Environ. Prot. 2001, 20, 88–90. [Google Scholar]
- Yang, W.S.; Li, C.R.; Lan, J.; An, H.L. The spread mode and dispersal history of Chinese citrus fly by literature research. J. Yangtze Univ. (Nat. Sci. Ed.) 2013, 10, 8–11. [Google Scholar]
- Xia, Y.L.; Ma, X.L.; Hou, B.H.; Ouyang, G.C. A review of Bactrocera minax (Diptera: Tephritidae) in China for the purpose of safeguarding. Adv. Entomol. 2018, 6, 35–61. [Google Scholar] [CrossRef]
- Cooper, R.L.; McLetchie, D.N. Monitoring carbon dioxide production by Drosophila larvae. Drosoph. Inform. Serv. 2004, 87, 88–91. [Google Scholar]
- Zhao, Y.C.; Guo, S.Z.; Zhang, Z.M. Observation on the erosion period and overwintering mortality of Bactrocera minax. Plant Quar. 2002, 16, 339–340. [Google Scholar]
- Wang, F.L.; Chambi, C.; Li, Z.Y.; Huang, C.; Ma, Y.K.; Li, C.R.; Tian, X.H.; Sangija, F.; Ntambo, M.S.; Kankonda, O.M.; Hafeez, S.; Anwar, T.; Sharif, R. Influence of supplemental protein on the life expectancy and reproduction of the chinese citrus fruit fly, Bactrocera minax (Enderlein) (Tetradacus minax) (Diptera:Tephritidae). J. Insect Sci. 2018, 18, 1–8. [Google Scholar] [CrossRef]
- Sun, R.H.; Wu, H.B.; Zhang, K.P.; Li, A.H.; Zhang, X.P. Influence of the humidity and water immersion on egg hatch and adult emergence of Carposina sasakii. J. Fruit Sci. 2013, 30, 449–451. [Google Scholar]
- Cavallaro, M.C.; Barnhart, M.C.; Hoback, W.W. Causes of rapid carrion beetle (coleoptera: Silphidae) death in flooded pitfall traps, response to soil flooding, immersion tolerance, and swimming behavior. Environ. Entomol. 2017, 46, 362–368. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, Z.F.; Wu, W.J. Effect of immersion in water on egg survival in the grasshopper Fruhstorferiola tonkinensis. Chin. Bull. Entomol. 2009, 46, 895–897. [Google Scholar]
- Woodman, J.D. Surviving a flood: Effects of inundation period, temperature and embryonic development stage in locust eggs. B Entomol. Res. 2015, 105, 441–447. [Google Scholar] [CrossRef]
- Terzian, L.A.; Stahler, N. The effects of larval population density on some laboratory characteristics of anopheles quadrimaculatus say. J. Parasitol. 1949, 35, 487–495. [Google Scholar] [CrossRef]
- Montoya, P.; Flores, S.; Toledo, J. Effect of rainfall and soil moisture on survival of adults and immature stages of Anastrepha ludens and A. obliqua (Diptera: Tephritidae) under semi-field conditions. Fla. Entomol. 2008, 91, 643–650. [Google Scholar]
- Wang, J.; Fan, H.; Xiong, K.C.; Liu, Y.H. Transcriptomic and metabolomic profiles of Chinese citrus fly, Bactrocera minax (Diptera: Tephritidae), along with pupal development provide insight into diapause program. PLoS ONE 2017, 12, e0181033. [Google Scholar] [CrossRef]
- Hoback, W.W.; Podrabsky, J.E.; Higley, L.G.; Stanley, D.W.; Hand, S.C. Anoxia tolerance of con-familial tiger beetle larvae is associated with differences in energy flow and anaerobiosis. J. Comp. Physiol. 2000, 170, 307–314. [Google Scholar] [CrossRef]
- Hou, B.; Xie, Q.; Zhang, R. Depth of pupation and survival of the Oriental fruit fly, Bactrocera dorsalis (Diptera: Tephritidae) pupae at selected soil moistures. Appl. Entomol. Zool. 2006, 41, 515–520. [Google Scholar] [CrossRef]
- Ni, W.L.; Chen, H.J.; Li, Z.H.; Wu, J.J.; Qu, W.W. Influence of soil moisture and water immersion on the emergence rate of Bactrocera correcta(Bezzi). Plant Quar. 2011, 275, 192–195. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Regression Equation | R | R0.05 | R0.01 |
|---|---|---|---|---|
| Recovery ratio | YR = 96.669/(1 + Exp(−6.814 + 0.656X)) | 0.988 | 0.532 | 0.661 |
| Larvae mortality ratio | YM = 106.543/(1 + Exp(4.028 − 0.514X)) | 0.997 | 0.514 | 0.641 |
| Pupation ratio | YP = 90.735/(1 + Exp(−5.448 + 0.799X)) | 0.999 | 0.532 | 0.661 |
| Parameter | Estimated | 95% CI | R2 |
|---|---|---|---|
| Recovery ratio | |||
| A | 96.669 | (90.616, 102.722) | 0.977 |
| b | −6.814 | (−8.860, −4.768) | |
| k | −0.656 | (−0.842, −0.470) | |
| Larvae mortality ratio | |||
| A | 106.543 | (99.429, 113.657) | 0.994 |
| b | 4.028 | (3.503, 4.552) | |
| k | 0.514 | (0.432, 0.595) | |
| Pupation ratio | |||
| A | 90.735 | (88.184, 93.286) | 0.998 |
| b | −5.448 | (−6.115, −4.780) | |
| k | −0.799 | (−0.888, −0.710) |
| Parameter | Time Estimates | |||
|---|---|---|---|---|
| RD50/MD50/PD50 | RD90/MD90/PD90 | |||
| Days | (95% CI) | Days | (95% CI) | |
| Recovery ratio | 10.3 | (−5.9, 26.5) | 6.4 | (−24.6, 37.4) |
| Larvae mortality ratio | 7.6 | (−1.6, 16.8) | 11.1 | (−6.5, 28.7) |
| Pupation ratio | 6.6 | (−0.6, 13.7) | 0.8 | (−12.7, 14.3) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Chambi, C.; Du, T.; Huang, C.; Wang, F.; Zhang, G.; Li, C.; Juma Kayeke, M. Effects of Water Immersion and Soil Moisture Content on Larval and Pupal Survival of Bactrocera minax (Diptera: Tephritidae). Insects 2019, 10, 138. https://doi.org/10.3390/insects10050138
Li Z, Chambi C, Du T, Huang C, Wang F, Zhang G, Li C, Juma Kayeke M. Effects of Water Immersion and Soil Moisture Content on Larval and Pupal Survival of Bactrocera minax (Diptera: Tephritidae). Insects. 2019; 10(5):138. https://doi.org/10.3390/insects10050138
Chicago/Turabian StyleLi, Zaiyuan, Consolatha Chambi, Tianhua Du, Cong Huang, Fulian Wang, Guifen Zhang, Chuanren Li, and Mohamed Juma Kayeke. 2019. "Effects of Water Immersion and Soil Moisture Content on Larval and Pupal Survival of Bactrocera minax (Diptera: Tephritidae)" Insects 10, no. 5: 138. https://doi.org/10.3390/insects10050138
APA StyleLi, Z., Chambi, C., Du, T., Huang, C., Wang, F., Zhang, G., Li, C., & Juma Kayeke, M. (2019). Effects of Water Immersion and Soil Moisture Content on Larval and Pupal Survival of Bactrocera minax (Diptera: Tephritidae). Insects, 10(5), 138. https://doi.org/10.3390/insects10050138