Abundance and Population Decline Factors of Chrysopid Juveniles in Olive Groves and Adjacent Trees



Abstract
:1. Introduction
2. Materials and Methods
2.1. Area of Study
2.2. Collection of Samples
2.3. Statistical Analysis
3. Results
3.1. Analysis of Collected Cocoons
3.2. Abundance and Identification of Chrysopids
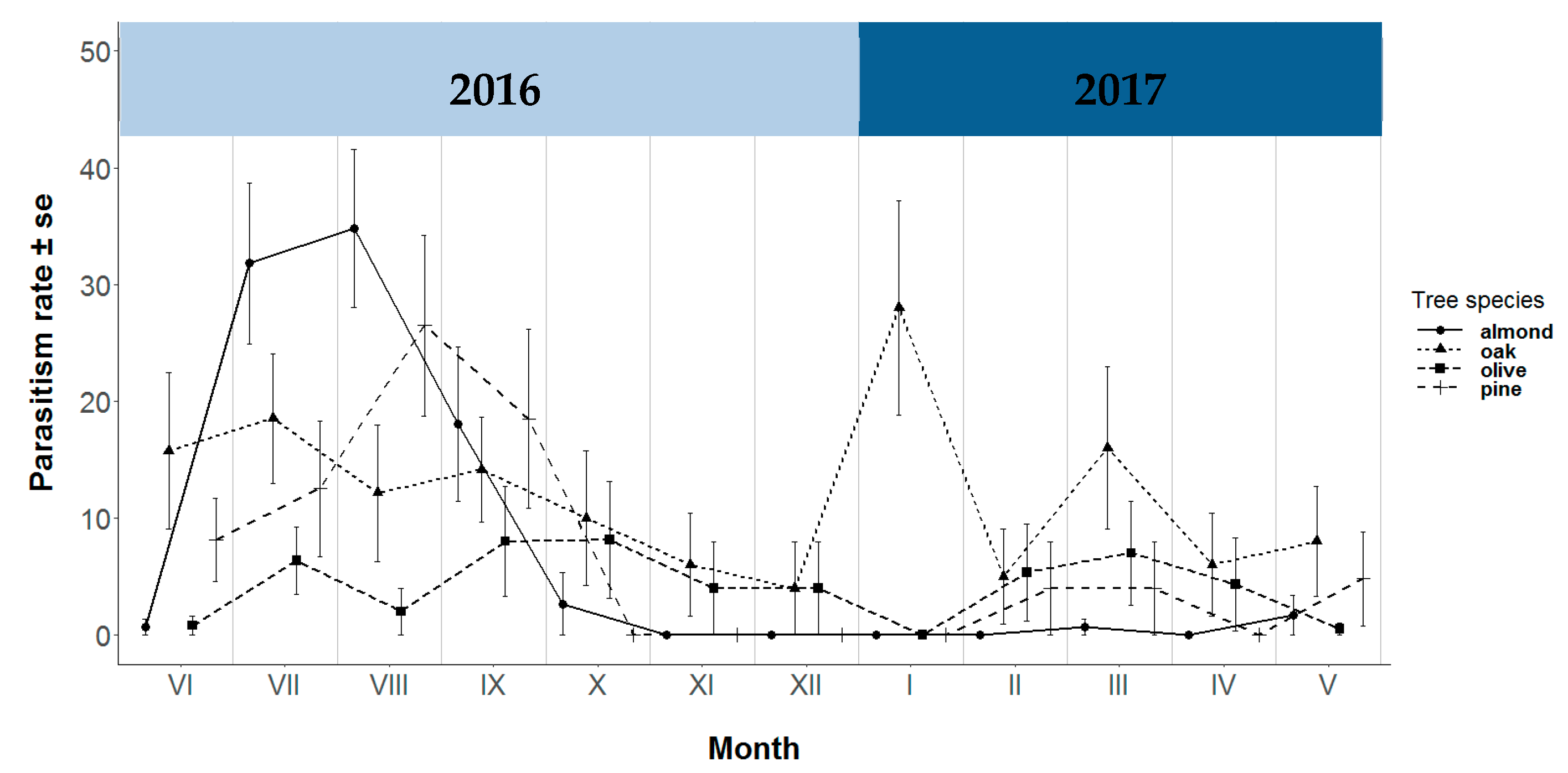
3.3. Parasitism Rate and Juvenile Chrysopid Parasitoid Complex
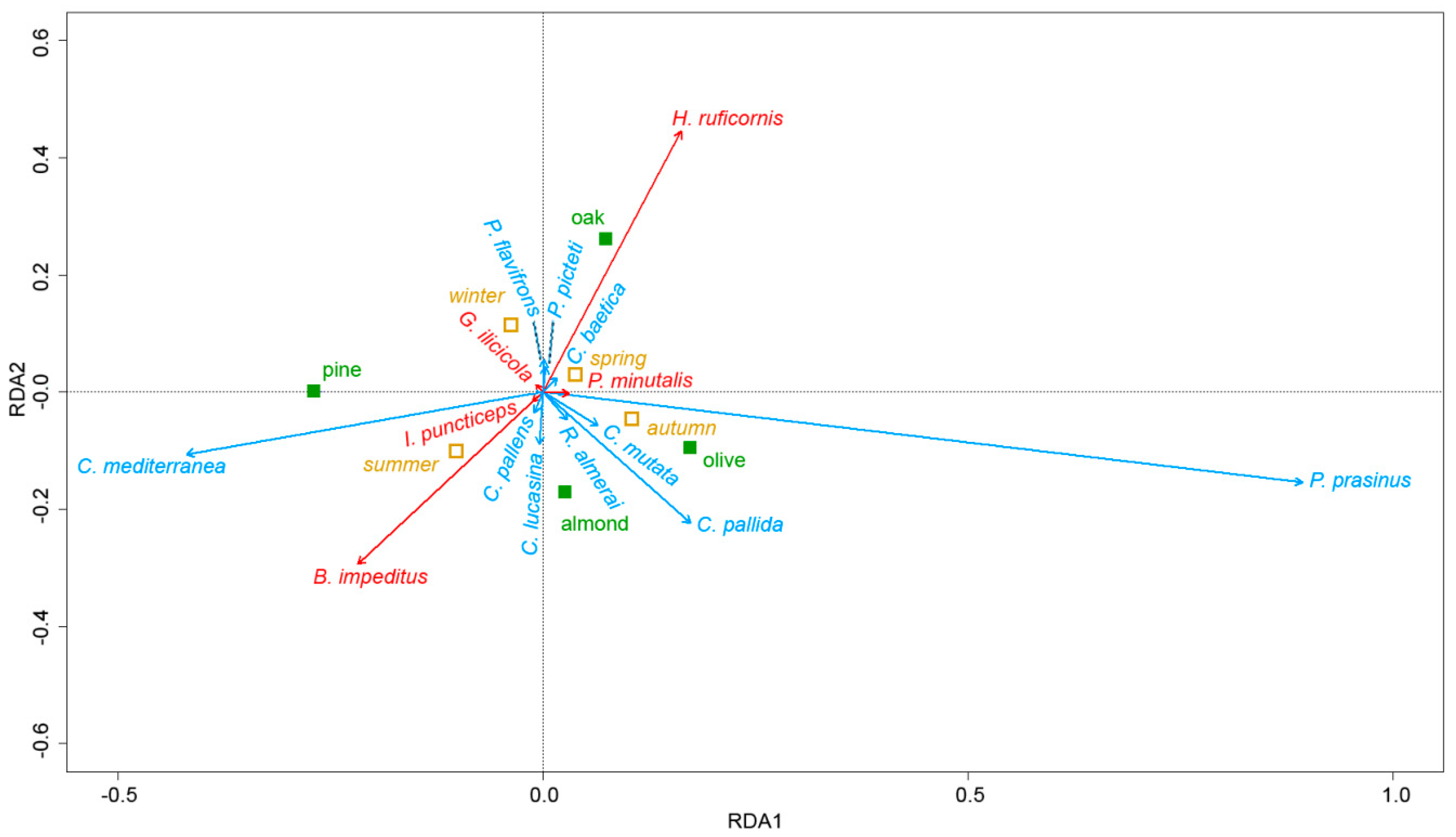
3.4. Multivariate Analysis of the Relationship between Parasitoid and Chrysopid Species, Tree Species and Season
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Monserrat, V.J. Los crisópidos de la Península Ibérica y Baleares (Insecta, Neuropterida, Neuroptera: Chrysopidae). Graellsia 2016, 72, 1–123. [Google Scholar] [CrossRef]
- Duelli, P. Lacewings in field crops. In Lacewings in the Crop Environment; Whittington, A.E., McEwen, P.K., New, T.R., Eds.; Cambridge University Press: Cambridge, UK, 2001; pp. 158–171. [Google Scholar] [CrossRef]
- Szentkirályi, F. Lacewings in vegetables, forests, and other crops. In Lacewings in the Crop Environment; Whittington, A.E., McEwen, P.K., New, T.R., Eds.; Cambridge University Press: Cambridge, UK, 2001; pp. 239–292. [Google Scholar]
- Pappas, M.L.; Broufas, G.D.; Koveos, D.S. Chrysopid predators and their role in biological control. J. Entomol. 2011, 8, 301–326. [Google Scholar] [CrossRef]
- Principi, M.M.; Canard, M. Feeding habits. In Biology of Chrysopidae; Canard, M., Séméria, Y., New, T.R., Eds.; Dr. W. Junk Publishers: The Hague, The Netherlands, 1984; Volume 27, pp. 76–92. [Google Scholar]
- New, T.R. A review of the biology of Neuroptera Planipennia. Neuroptera Int. Suppl. 1986, 1, 1–57. [Google Scholar]
- Oswald, J.D. LDL NSW: Neuropterida Species of the World (Version June 2017). Available online: www.catalogueoflife.org/col (accessed on 18 September 2018).
- Alrouechdi, K. Relations comportementales et trophiques entre Chrysoperla carnea (Stephens) (Neuroptera; Chrysopidae) et trois principaux ravageurs de l’olivier. I. La Teigne de L’olivier Prays oleae Bern. (Lep. Hyponomeutidae). Neuroptera Int. 1981, 1, 122–134. [Google Scholar]
- Tauber, M.J.; Tauber, C.A.; LopezArroyo, J.I. Life-history variation in Chrysoperla carnea: Implications for rearing and storing a Mexican population. Biol. Control 1997, 8, 185–190. [Google Scholar] [CrossRef]
- Gerling, D.; Kravchenko, V.; Lazare, M. Dynamics of common green lacewing (Neuroptera: Chrysopidae) in Israeli cotton fields in relation to whitefly (Homoptera: Aleyrodidae) populations. Environ. Entomol. 1997, 26, 815–827. [Google Scholar] [CrossRef]
- Daane, K.M.; Yokota, G.Y.; Zheng, Y.; Hagen, K.S. Inundative release of common green lacewings (Neuroptera: Chrysopidae) to suppress Erythroneura variabilis and E. elegantula (Homoptera: Cicadellidae) in vineyards. Environ. Entomol. 1996, 25, 1224–1234. [Google Scholar] [CrossRef]
- New, T.R. Lacewings (Neuroptera) as biological control agents. Vic. Entomol. 1975, 5, 102–103. [Google Scholar]
- Henry, C.S. The proliferation of cryptic species in Chrysoperla green lacewings through song divergence. Florida Entomol. 1985, 68, 18–38. [Google Scholar] [CrossRef]
- Tauber, C.A.; Tauber, M.J. Ecophysiological responses in life-history evolution: Evidence for their importance in a geographically widespread insect species complex. Can. J. Zool.-Rev. Can. Zool. 1986, 64, 875–884. [Google Scholar] [CrossRef]
- Henry, C.S.; Brooks, S.J.; Johnson, J.B.; Duelli, P. Chrysoperla lucasina (Lacroix): A distinct species of green lacewing, confirmed by acoustical analysis (Neuroptera: Chrysopidae). Syst. Entomol. 1996, 21, 205–218. [Google Scholar] [CrossRef]
- Henry, C.S.; Brooks, S.J.; Duelli, P.; Johnson, J.B. Discovering the true Chrysoperla carnea (Insecta: Neuroptera: Chrysopidae) using song analysis, morphology, and ecology. Ann. Entomol. Soc. Am. 2002, 95, 172–191. [Google Scholar] [CrossRef]
- Canard, M.; Thierry, D. A historical perspective on nomenclature within the genus Chrysoperla Steinmann, 1964 in Europe: The carnea-complex (Neuroptera Chrysopidae). Ann. Museo Civ. Storia Nat. Ferrara 2007, 8, 173–179. [Google Scholar]
- Henry, C.S.; Brooks, S.J.; Duelli, P.; Johnson, J.B.; Wells, M.M.; Mochizuki, A. Obligatory duetting behaviour in the Chrysoperla carnea-group of cryptic species (Neuroptera: Chrysopidae): Its role in shaping evolutionary history. Biol. Rev. 2013, 88, 787–808. [Google Scholar] [CrossRef]
- Price, B.W.; Henry, C.S.; Hall, A.C.; Mochizuki, A.; Duelli, P.; Brooks, S.J. Singing from the Grave: DNA from a 180 Year Old Type Specimen Confirms the Identity of Chrysoperla carnea (Stephens). PLoS ONE 2015, 10, 11. [Google Scholar]
- Campos, M. Observaciones sobre la bioecologia de Chrysoperla carnea (Stephens) (Neuroptera: Chrysopidae) en el sur de España. Neuroptera Int. 1989, 5, 159–164. [Google Scholar]
- Monserrat, V.J.; Marín, F. Plant substrate-specifity of Iberian Chrysopidae (Insecta, Neuroptera). Acta Oecol.-Int. J. Ecol. 1994, 15, 119–131. [Google Scholar]
- Porcel, M. Estudio de la Bioecología de la Familia Chrysopidae (Insecta: Neuroptera) Desde la Perspectiva de su Incremento y Conservación en el Olivar. Ph.D. Thesis, Universidad de Granada, Granada, Spain, 2012. [Google Scholar]
- Szentkirályi, F. Lacewings in fruit and nut crops. In Lacewings in the Crop Environment; Whittington, A.E., McEwen, P.K., New, T.R., Eds.; Cambridge University Press: Cambridge, UK, 2001; pp. 172–238. [Google Scholar]
- Alrouechdi, K. Les Chrysopides en Verger D’oliviers: Bio-écologie de Chrysoperla carnea (Steph.) (Neuroptera, Chrysopidae); Relations Comportementales et Trophiques Avec Certaines Espèces Phytophages. Ph.D. Thesis, L‘Université Pierre et Marie Curie, Paris, France, 1980. [Google Scholar]
- Ramos, P.; Campos, M.; Ramos, J. Estabilización del ataque de Prays oleae Bern. y de la actividad de los depredadores oófagos sobre el fruto del olivo. Bol. Sanidad Vegetal. Plagas 1984, 10, 239–243. [Google Scholar]
- Campos, M. Lacewings in Andalusian olive orchards. In Lacewings in the Crop Environment; Whittington, A.E., McEwen, P.K., New, T.R., Eds.; Cambridge University Press: Cambridge, UK, 2001; pp. 492–497. [Google Scholar]
- McEwen, P.K.; Jervis, M.A.; Kidd, N.A.C. Use of a sprayed L-tryptophan solution to concentrate numbers of the green lacewing Chrysoperla carnea in olive tree canopy. Entomol. Exp. Appl. 1994, 70, 97–99. [Google Scholar] [CrossRef]
- McEwen, P.K.; Ruiz, J. Relationship between non-olive vegetation and green lacewing eggs in a Spanish olive orchard. Antenna 1994, 18, 148–150. [Google Scholar]
- Porcel, M.; Cotes, B.; Castro, J.; Campos, M. The effect of resident vegetation cover on abundance and diversity of green lacewings (Neuroptera: Chrysopidae) on olive trees. J. Pest Sci. 2017, 90, 195–196. [Google Scholar] [CrossRef]
- McEwen, P.K.; New, T.R.; Whittington, A.E. Lacewings in the Crop Environment; Cambridge University Press: Cambridge, UK, 2001; p. 546. [Google Scholar]
- Canard, M.; Séméria, Y.; New, T.R. Biology of Chrysopidae; Dr. W. Junk Publishers: The Hague, The Netherlands, 1984; Volume 27, pp. 1–294. [Google Scholar]
- Canard, M.; Volkovich, T.A. Outlines of lacewing development. In Lacewings in the Crop Environment; Whittington, A.E., McEwen, P.K., New, T.R., Eds.; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Chang, Y.F.; Tauber, M.J.; Tauber, C.A. Storage of the mass-produced predator Chrysoperla carnea (Neuroptera: Chrysopidae): Influence of photoperiod, temperature, and diet. Environ. Entomol. 1995, 24, 1365–1374. [Google Scholar] [CrossRef]
- Nadeem, S.; Hamed, M.; Ishfaq, M.; Nadeem, M.K.; Hasnain, M.; Saeed, N.A. Effect of storage duration and low temperatures on the developmental stages of Chrysoperla carnea (Stephens) (Neuroptera: Chrysopidae). J. Anim. Plant Sci. 2014, 24, 1569–1572. [Google Scholar]
- Pappas, M.L.; Broufas, G.D.; Koveos, D.S. Effect of relative humidity on development, survival and reproduction of the predatory lacewing Dichochrysa prasina (Neuroptera: Chrysopidae). Biol. Control 2008, 46, 234–241. [Google Scholar] [CrossRef]
- Pappas, M.L.; Karagiorgou, E.; Papaioannou, G.; Koveos, D.S.; Broufas, G.D. Developmental temperature responses of Chrysoperla agilis (Neuroptera: Chrysopidae), a member of the European carnea cryptic species group. Biol. Control 2013, 64, 291–298. [Google Scholar] [CrossRef]
- Morris, T.I.; Campos, M.; Jervis, M.A.; McEwen, P.K.; Kidd, N.A.C. Potential effects of various ant species on green lacewing, Chrysoperla carnea (Stephens) (Neuropt., Chrysopidae) egg numbers. J. Appl. Entomol.-Z. Angew. Entomol. 1998, 122, 401–403. [Google Scholar] [CrossRef]
- Vinson, S.B.; Scarborough, T.A. Impact of the imported fire ant on laboratory populations of cotton aphid (Aphis gossypii) predators. Fla. Entomol. 1989, 72, 107–111. [Google Scholar] [CrossRef]
- Dinter, A. Intraguild predation between erigonid spiders, lacewing larvae and carabids. J. Appl. Entomol. 1998, 122, 163–167. [Google Scholar] [CrossRef]
- Cisneros, J.; Rosenheim, J.A.Y. Ontogenetic change of prey preference in the generalist predator Zelus renardii and its influence on predator–predator interactions. Ecol. Entomol. 1997, 22, 399–407. [Google Scholar] [CrossRef]
- Lucas, E.; Coderre, D.; Brodeur, J. Instar-specific defense of Coleomegilla maculata lengi (Col.: Coccinellidae): Influence on attack success of the intraguild predator Chrysoperla rufilabris (Neur.: Chrysopidae). Entomophaga 1997, 42, 3–12. [Google Scholar] [CrossRef]
- Canard, M.; Duelli, P. Predatory behavior of larvae and cannibalism. In Biology of Chrysopidae; Canard, M., Séméria, Y., New, T.R., Eds.; Dr. W. Junk Publishers: The Hague, The Netherlands, 1984; Volume 27, pp. 92–100. [Google Scholar]
- Sidor, C. A polyhedral virus disease of Chrysopa perla (L.). Virology 1960, 10, 551–552. [Google Scholar] [CrossRef]
- Ventura, M.A.; Garcia, V.; Canard, M. Antibiosis effect caused by the entomopathogenic fungus Metarhizium anisopliae (Metschnikoff) Sorokin variety anisopliae Tulloch, to a “common green lacewing” Chrysoperla kolthoffi (Navás) (Neuroptera: Chrysopidae). J. Neuropterol. 2000, 3, 33–41. [Google Scholar]
- Clancy, D.W. The insect parasites of the Chrysopidae (Neuroptera). Univ. Calif. Publ. Entomol. 1946, 7, 403–496. [Google Scholar]
- Killington, F.J. The parasites of Neuroptera with special reference to those attacking British species. Trans. Entomol. Soc. South England 1932, 8, 84–91. [Google Scholar]
- Alrouechdi, K.; Panis, A. Les parasites de Chrysoperla carnea Steph. (Neuroptera, Chrysopidae) sur Olivier en Provence. Agronomie 1981, 1, 139–141. [Google Scholar] [CrossRef]
- Alrouechdi, K.; Séméria, Y.; New, T.R. Natural enemies. In Biology of Chrysopidae; Canard, M., Séméria, Y., New, T.R., Eds.; Dr. W. Junk Publishers: The Hague, The Netherlands, 1984; Volume 27, pp. 187–204. [Google Scholar]
- Muma, M.H. Hymenopterous parasites of Chrysopidae on Florida citrus. Fla. Entomol. 1959, 42, 149–153. [Google Scholar] [CrossRef]
- Alrouechdi, K.; Canard, M.; Pralavorio, R.; Arambourg, Y. Influence du complexe parasitaire sur les populations de chrysopides (Neuroptera) dans un verger d’oliviers du Sud-Est de la France. Z. Angew. Entomol.-J. Appl. Entomol. 1981, 91, 411–417. [Google Scholar] [CrossRef]
- Principi, M.M. Contributi allo studio dei Neurotteri Italiani. VII. Osservazioni su alcuni parassiti di crisopidi. Boll. dell’Istituto Entomol. Univ. Studi Bol. 1948, 17, 93–121. [Google Scholar]
- Mehra, B.P. Biology of Chrysopa lacciperda Kimmins. J. Bombay Nat. Hist. Soc. 1966, 63, 215–218. [Google Scholar]
- Ickert, G. Beiträge zur Biologie einheimischer Chrysopiden (Planipennia, Chrysopidae). Entomol. Abhandlungen Staatliches Museum Tierkunde Dresden 1968, 36, 123–192. [Google Scholar]
- New, T.R. A recent host record of Helorus coruscus Hal. (Hym., Heloridae). Entomol. Mon. Mag. 1967, 102, 86. [Google Scholar]
- New, T.R. An Australian species of Helorus Latreille (Hymenoptera: Heloridae). Aust. J. Entomol. 1975, 14, 15–17. [Google Scholar] [CrossRef]
- New, T.R. Hymenopterous parasites of some larvae Chrysopidae (Neuroptera) near Melbourne, Australia. Neuroptera Int. 1982, 2, 33–36. [Google Scholar]
- New, T.R. Trap-banding as a collecting method for Neuroptera and their parasites, and some results obtained. Entomol. Gazette 1967, 18, 37–44. [Google Scholar]
- Judd, W. Emergence of the lacewing, Chrysopa harrisii Fitch (Neuroptera) and three hymenopterous parasites from the cocoon. Ann. Entomol. Soc. Am. 1949, 42, 461–464. [Google Scholar] [CrossRef]
- Daane, K.M. Ecological studies of released lacewings in crops. In Lacewings in the Crop Environment; Whittington, A.E., McEwen, P.K., New, T.R., Eds.; Cambridge University Press: Cambridge, UK, 2001; pp. 338–350. [Google Scholar] [CrossRef]
- Withycombe, C.L. Notes on the biology of some British Neuroptera (Planipennia). Trans. R. Entomol. Soc. Lond. 1923, 70, 501–594. [Google Scholar] [CrossRef]
- Monserrat, V.J. Estrategias de defensa visual en los Neuropterida Ibéricos (Megaloptera, Raphidioptera, Neuroptera). Bol. Soc. Entomol. Aragon. 2015, 57, 459–480. [Google Scholar]
- Smith, R.C. The trash-carrying habit of certain lacewing larvae. Sci. Mon. N. Y. 1926, 23, 265–267. [Google Scholar]
- Duelli, P. A missing link in the evolution of the egg pedicel in lacewings. Experientia 1986, 42, 624. [Google Scholar] [CrossRef]
- Duelli, P. Oviposition. In Biology of Chrysopidae; Canard, M., Séméria, Y., New, T.R., Eds.; Dr. W. Junk Publishers: The Hague, The Netherlands, 1984; Volume 27, pp. 129–133. [Google Scholar]
- Monserrat, V.J. Contribución al conocimiento de los neurópteros de Toledo. Graellsia 1980, 34, 177–193. [Google Scholar]
- Güsten, R.; Dettner, K. The prothoracic gland of the Chrysopidae (Neuropteroidea: Planipennia). In Proceedings of the 4th European Congress of Entomology and the XIII Internationale Symposium für die Entomofaunistik Mitteleuropas, Gödöllö, Hungary, 1–6 September 1991; Zombori, L., Peregovits, L., Eds.; 1991; Volume 1, pp. 60–65. [Google Scholar]
- Blum, M.S.; Wallace, J.D.; Fales, H.M. Skatole and tricedene: Identification and possible role in a chrysopid secretion. Insect Biochem. 1973, 3, 353–357. [Google Scholar] [CrossRef]
- Rothschild, M.; Euw, J.V.; Reichstein, T. Cardiac glycosides in a scale insect (Aspidiotus), a ladybird (Coccinella) and a lacewing (Chrysopa). Physiol. Entomol. 1973, 48, 89–90. [Google Scholar] [CrossRef]
- Kennett, C.E. Defense mechanism exhibited by larvae of Chrysopa californica Coq. (Neuroptera: Chrysopidae). Pan-Pac. Entomol. 1948, 24, 209–211. [Google Scholar]
- Smith, R.C. A study of the biology of the Chrysopidae. Ann. Entomol. Soc. Am. 1921, 14, 27–35. [Google Scholar] [CrossRef]
- New, T.R. Note on the debris-carrying habit in larvae of British Chrysopidae (Neuroptera). Entomol. Gazette 1969, 20, 119–124. [Google Scholar]
- Eisner, T.; Eisner, M. Coiling into a sphere: Defensive behavior of a trash-carrying chrysopid larva Leucochrysa (Nodita) pavida (Neuroptera: Chrysopidae). Entomol. News 2002, 113, 6–10. [Google Scholar]
- Monserrat, V.J.; Diaz-Aranda, L.M. Larval stages of the Iberian green-lacewings (Insecta, Neuroptera, Chrysopidae), new data on larval morphology applicable to the family systematics. Graellsia 2012, 68, 31–158. [Google Scholar] [CrossRef]
- González, R.; Al-Asaad, S.; Bozsik, A. Influencia de las masas forestales en la diversidad y abundancia de los crisópidos (Neur. Chrysopidae) del olivar. Cuaderno Soc. Española Cienc. For. 2008, 26, 33–38. [Google Scholar]
- Alcalá Herrera, R.; Campos, M.; Ruano, F. Late summer oviposition of green lacewings (Neuroptera: Chrysopidae) on olive groves and adjacent trees. Environ. Entomol. 2019. [Google Scholar] [CrossRef]
- Principi, M.M. Contributi allo studio dei Neurotteri Italiani. V. Ricerche su Chrysopa formosa Brauer e su alcuni suoi parassiti. Boll. dell’Inst. Entomol. Univ. Bol. 1947, 16, 134–175. [Google Scholar]
- Principi, M.M. Contributi allo studio dei Neurotteri italiani. XIII. Studio morfologico, etologico e sistematico di un gruppo omogeneo di specie del Gen. Chrysopa Leach (C. flavifrons Brauer, prasina Burm. e clathrata Schn.). Boll. dell’Istituto Entomol. Univ. Studi Bol. 1956, 21, 319–410. [Google Scholar]
- Campos, M. Influencia del complejo parasitario sobre las poblaciones de Chrysoperla carnea (Neuroptera, Chrysopidae) en olivares del sur de España. Neuroptera Int. 1986, 4, 97–105. [Google Scholar]
- Putman, W.L. Biological notes on the Chrysopidae. Can. J. Res. 1937, 15, 29–37. [Google Scholar] [CrossRef]
- Neuenschwander, P.; Michelakis, S. The seasonal and spatial distribution of adult and larval chrysopids on olive-trees in Crete. Acta Oecol. Oecol. Appl. 1980, 1, 93–102. [Google Scholar]
- Junta de Andalucía. Análisis de las Plantaciones de Olivar en Andalucía. Encuesta sobre Superficies y Rendimientos de Cultivos en España (ESYRCE); Secretaria General de Agricultura y Alimentación, Servicios de Estudios y Estadísticas; Consejería de Agricultura, Pesca y Desarrollo Rural: Sevilla, Spain, 2015. [Google Scholar]
- European Union. Reglamento (CE) no. 834/2007 del Consejo, de 28 de junio de 2007, sobre producción y etiquetado de los productos ecológicos y por el que se deroga el Reglamento (CEE) no. 2092/91. Luxemburgo: Diario Oficial de la Unión Europea, de 20 de julio de 2007, no. 189. 2007, pp. 1–23. Available online: https://eur-lex.europa.eu/legal-content/ES/ALL/?uri=celex:32007R0834 (accessed on 1 March 2018).
- European Union. Reglamento (CE) no. 889/2008 de la Comisión, de 5 de septiembre de 2008, por el que se establecen disposiciones de aplicación del Reglamento (CE) no. 834/2007 del Consejo sobre producción y etiquetado de los productos ecológicos, con respecto a la producción ecológica, su etiquetado y su control. Luxemburgo: Diario Oficial de la Unión Europea, de 18 de septiembre de 2008, no. 250. 2008, pp. 1–84. Available online: https://eur-lex.europa.eu/legal-content/ES/TXT/?uri=celex%3A32008R0889 (accessed on 1 March 2018).
- Goulet, H.; Huber, J.T. Hymenoptera of the World: An Identification Guide to Families; Canada, A., Ed.; Centre for Land and Biological Resources Research: Ottawa, ON, Canada, 1993. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, AT, USA, 2017. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2014, 67, 1–48. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer Science & Business Media: New York, NY, USA, 2002. [Google Scholar]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models, R Package version 0.2.0; 2018; Available online: http://CRAN.Rproject.org/package=DHARMa (accessed on 7 April 2019).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- De Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research, R Package version 1.2-8; 2017; Available online: http://CRAN.Rproject.org/package=agricolae (accessed on 7 April 2019).
- Oksanen, J.; Guillaume Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. vegan: Community Ecology Package, R Package version 2.5-2; 2018; Available online: http://CRAN.Rproject.org/package=vegan (accessed on 7 April 2019).
- Alrouechdi, K. Les chrysopides (Neuroptera) en oliveraie. In Progress in World’s Neuropterology; Proceedings of the 1st International Symposium on Neuropterology, Graz, Austria, 22–26 September 1980; Gepp, J., Ed.; Osterreichischen Akademie der Wissenschaften: Vienna, Austria, 1984. [Google Scholar]
- El-Taeif, H.A.; El-Ghariani, I.M.; Bataw, A.A. Some biological data on the predator Green Lacewing Chrysoperla carnea (Steph.) (Neuropera: Chrysopidae). Egypt. J. Biol. Pest Control 2008, 18, 243–248. [Google Scholar]
- Inclan, D.J.; Cerretti, P.; Marini, L. Landscape composition affects parasitoid spillover. Agric. Ecosyst. Environ. 2015, 208, 48–54. [Google Scholar] [CrossRef]
- Pantaleoni, R.A. Distribuzione spaziale di alcuni Neurotteri Planipenni su piante arboree. Boll. dell’Istituto Entomol. “Guido Grandi” dell’Univ. Bol. 1996, 50, 133–141. [Google Scholar]
- Campos, M.; Ramos, P. Crisópidos (Neuroptera) capturados en un olivar del sur de España. Neuroptera Int. 1983, 2, 219–227. [Google Scholar]
- Townes, H. A revision of the Heloridae (Hymenoptera). Contrib. Am. Entomol. Inst. 1977, 15, 1–12. [Google Scholar]
- Van Achterberg, C. European species of the genus Helorus Latreille (Hymenoptera: Heloridae), with description of a new species from Sulawesi (Indonesia). Zool. Meded. 2006, 80, 1. [Google Scholar]
- Buffington, M.L.; Copeland, R.S. Redescription of Helorus ruficornis Förster (Hymenoptera: Heloridae), with a New Synonymy and New Afrotropical Specimen Records. Proc. Entomol. Soc. Wash. 2016, 118, 330–344. [Google Scholar] [CrossRef]
- Algarra, A.; Segade, C.; Ventura, D.; Pujade, J. Dos citas nuevas para la Península Ibérica y Andorra de Helorus Latreille, 1802 (Hymenoptera: Proctotrupoidea: Heloridae). Bol. Asoc. Española Entomol. 1996, 20, 262–263. [Google Scholar]
- Marín, F. Las comunidades de neurópteros de la provincia de Alabcete (Insecta: Neuropteroidea). In Al-Basit Revista de Estudios Albacetenses; Diputación Provincial de Albacete: Albacete, Spain, 1994; pp. 247–304. [Google Scholar]
- Alrouechdi, K.; Lyon, J.P.; Canard, M.; Fournier, D. Les chrysopids (Neuroptera) récoltés dans une oliveraie du sud-est de la France. Acta Oecol.-Oecol. Appl. 1980, 1, 173–180. [Google Scholar]
- Smith, R.C. The Biology of the Chrysopidae; Cornell University Agricultural Experimental Station: Ithaca, NY, USA, 1922; Volume 58. [Google Scholar]
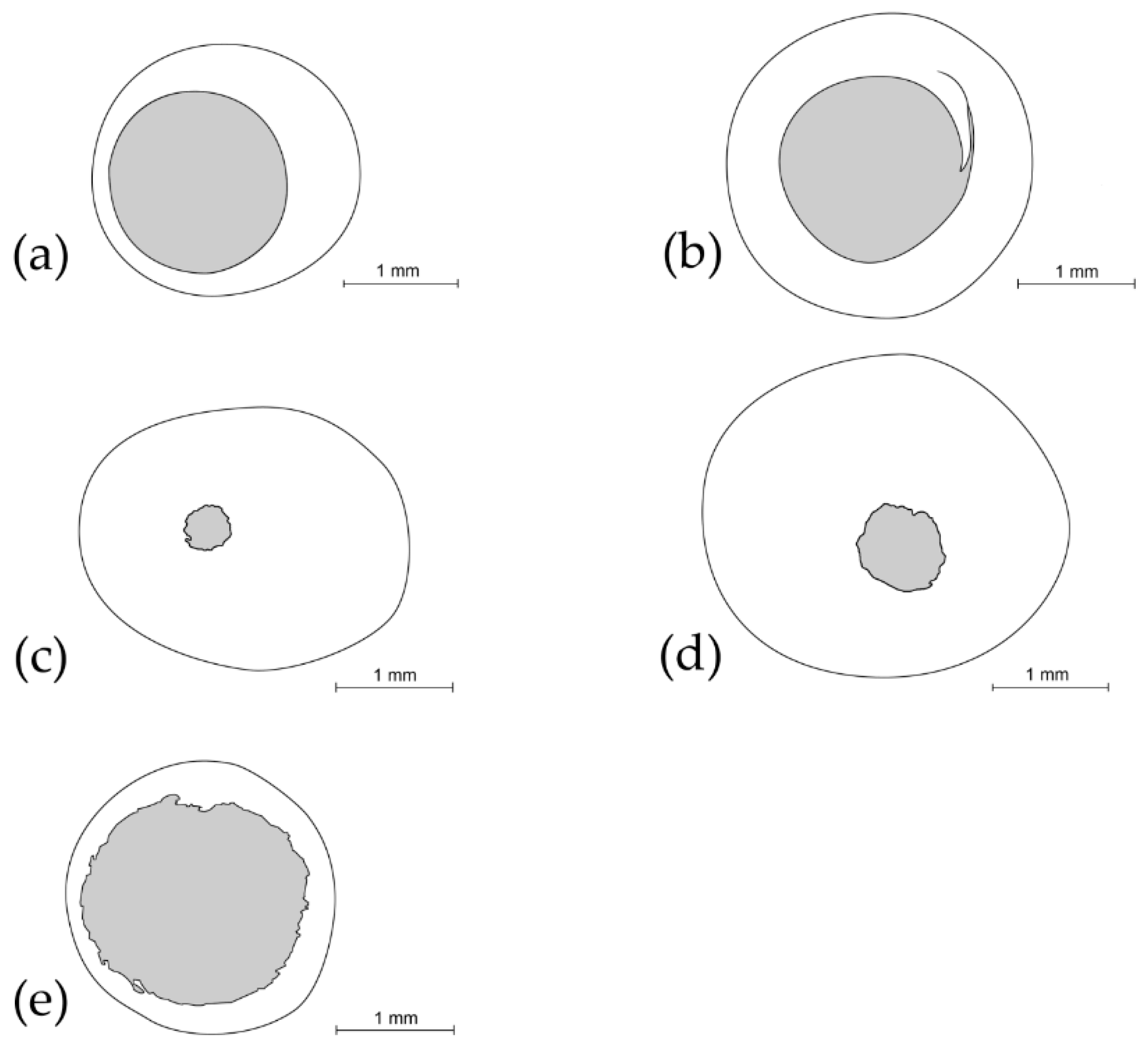
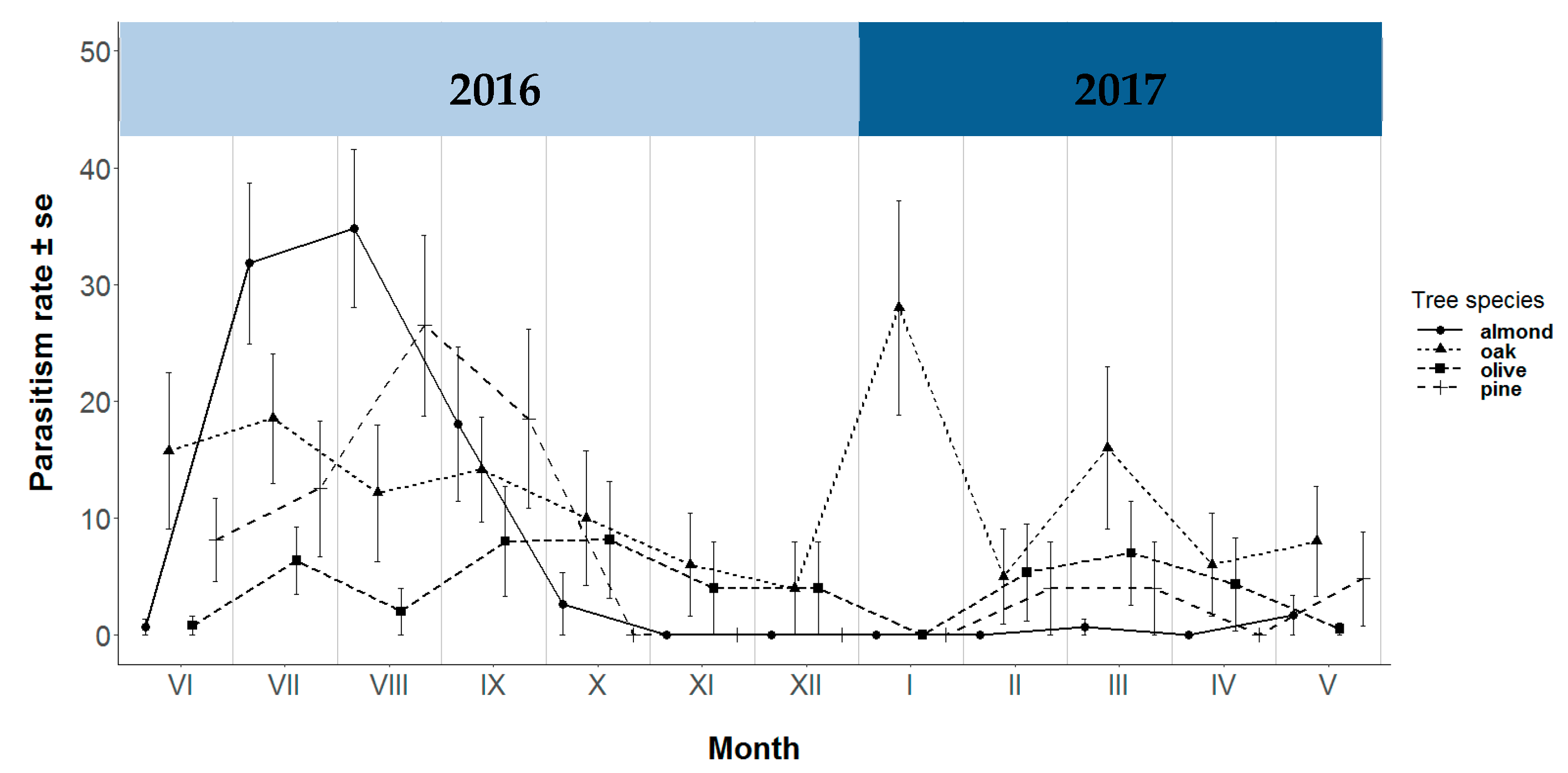
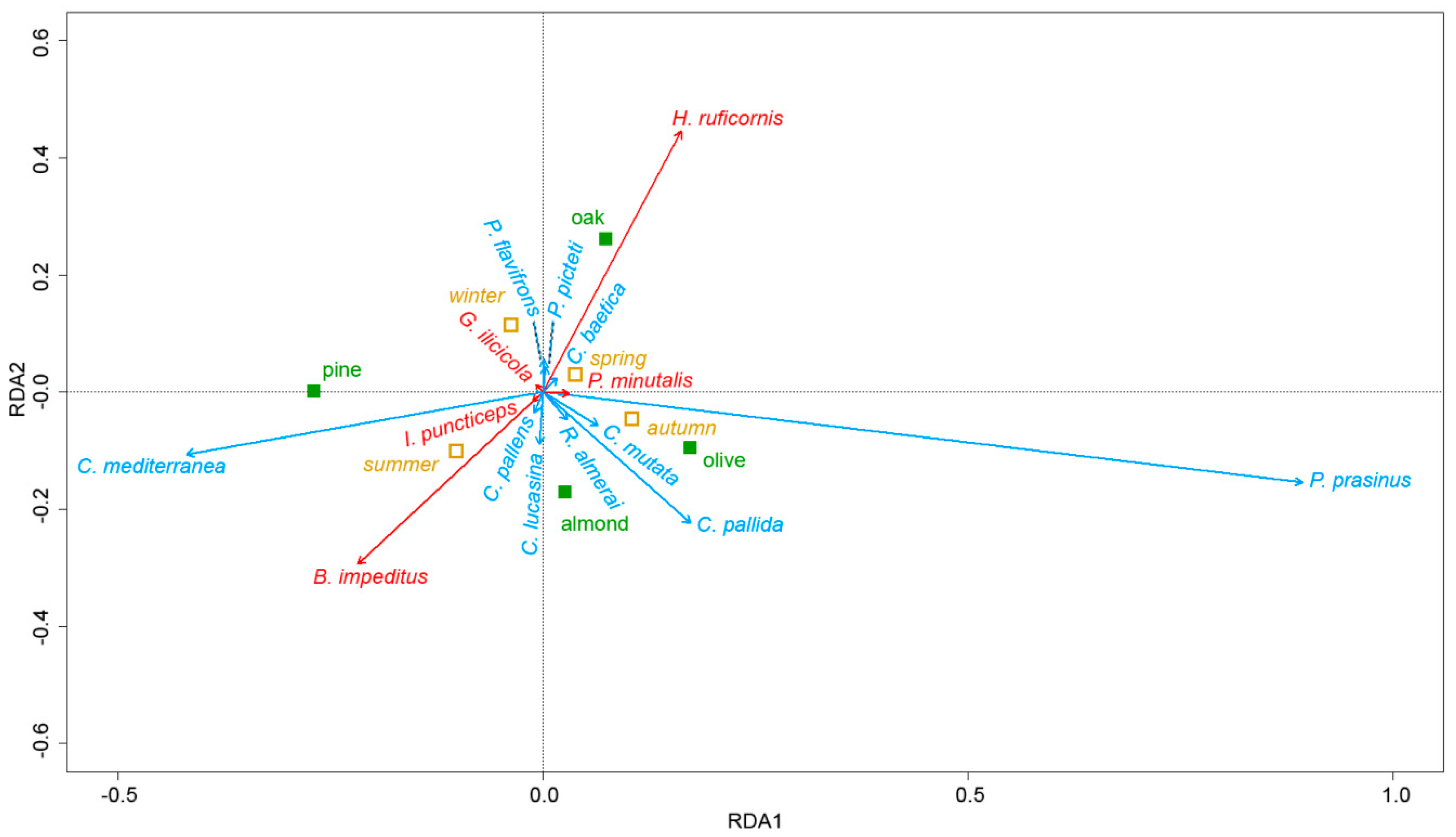

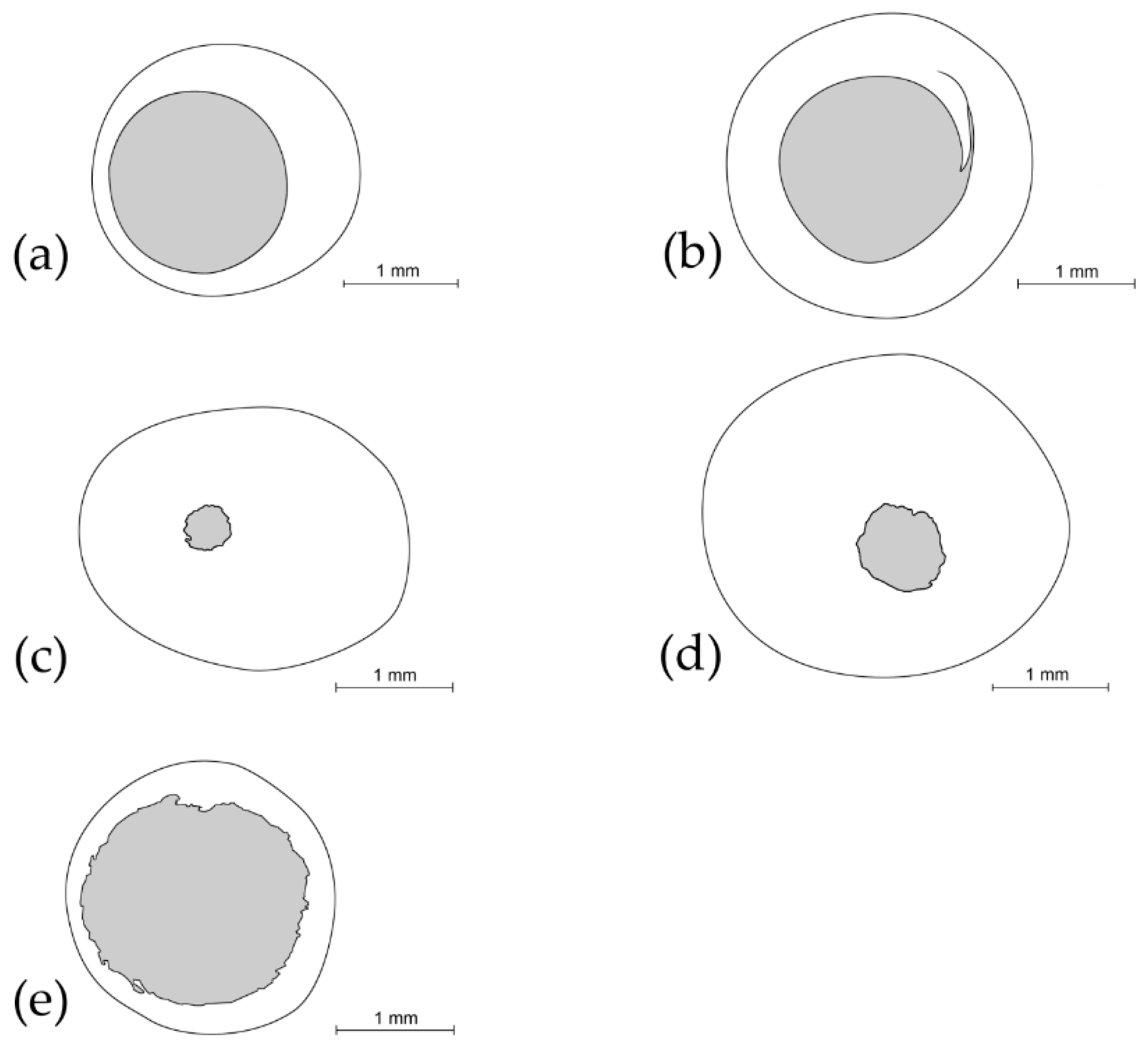{kind=link}
{kind=link}
{kind=link}
| Site | Coordinates | Area (ha) | Number of Trees Sampled | ||||
|---|---|---|---|---|---|---|---|
| Almond | Oak | Olive | Pine | Total | |||
| Norberto | 37°19’5.96” N; 3°34’9.92” W | 4.3 | 9 | 5 | 5 | 9 | 28 |
| La Pedriza | 37°20’17.44” N; 3°33’39.21” W | 0.9 | - | 5 | 5 | 8 | 18 |
| Los Almendros | 37°22’24.76” N; 3°37’46.03” W | 215 | 8 | 5 | 5 | - | 18 |
| Píñar (right) | 37°24’14.29” N; 3°29’14.13” W | 58 | - | 5 | 5 | 8 | 18 |
| Píñar (left) | 37°24’40.93” N; 3°28’52.41” W | 124 | 8 | 5 | 5 | - | 18 |
| Total | 25 | 25 | 25 | 25 | 100 | ||
| Season | Tree Species | Adult Chrysopids | Parasitized Chrysopids | Predated Chrysopids | Unknown Factors | Total |
|---|---|---|---|---|---|---|
| Summer | Almond | 122 (36.7) | 144 (43.4) | 33 (9.9) | 33 (9.9) | 332 |
| Oak | 49 (48) | 34 (33.3) | 9 (8.8) | 10 (9.8) | 102 | |
| Olive | 130 (76.9) | 9 (5.3) | 8 (4.7) | 22 (13) | 169 | |
| Pine | 109 (63.7) | 34 (19.9) | 3 (1.8) | 25 (14.6) | 171 | |
| Subtotal | 410 (53) | 221 (28.6) | 53 (6.8) | 90 (11.6) | 774 | |
| Autumn | Almond | 67 (45.3) | 54 (36.5) | 6 (4.1) | 21 (14.2) | 148 |
| Oak | 43 (55.8) | 18 (23.4) | 3 (3.9) | 13 (16.9) | 77 | |
| Olive | 77 (72.6) | 11 (10.4) | 4 (3.8) | 14 (13.2) | 106 | |
| Pine | 18 (42.9) | 19 (45.2) | 2 (4.8) | 3 (7.1) | 42 | |
| Subtotal | 205 (55) | 102 (27.3) | 15 (4) | 51 (13.7) | 373 | |
| Winter | Almond | 13 (68.4) | 0 (0) | 0 (0) | 6 (31.6) | 19 |
| Oak | 3 (18.8) | 11 (68.8) | 0 (0) | 2 (12.5) | 16 | |
| Olive | 24 (72.7) | 1 (3) | 0 (0) | 8 (24.2) | 33 | |
| Pine | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 | |
| Subtotal | 40 (58.8) | 12 (17.6) | 0 (0) | 16 (23.5) | 68 | |
| Spring | Almond | 23 (76.7) | 1 (3.3) | 0 (0) | 6 (20) | 30 |
| Oak | 26 (65) | 11 (27.5) | 0 (0) | 3 (7.5) | 40 | |
| Olive | 29 (60.4) | 7 (14.6) | 1 (2.1) | 11 (22.9) | 48 | |
| Pine | 8 (66.7) | 3 (25) | 0 (0) | 1 (8.3) | 12 | |
| Subtotal | 86 (66.2) | 22 (16.9) | 1 (0.8) | 21 (16.2) | 130 | |
| Total | 741 (55.1) | 357 (26.5) | 69 (5.1) | 178 (13.2) | 1345 |
| Model | Variable | χ² | Degree of Freedom (d.f.) | p Value | |
|---|---|---|---|---|---|
| Chrysopid abundance | Tree species | 29.168 | 3 | <0.001 | *** |
| Site | 48.165 | 4 | <0.001 | *** | |
| Month sampled | 320.795 | 11 | <0.001 | *** | |
| Parasitism rate | Tree species | 34.707 | 3 | <0.001 | *** |
| Site | 11.832 | 4 | 0.0187 | * | |
| Month sampled | 57.895 | 11 | <0.001 | *** | |
| Season | Tree Species | Cunctochrysa baetica | Chrysoperla lucasina | Chrysoperla mediterranea | Chrysoperla mutata | Chrysoperla pallida | Chrysopa pallens | Pseudomallada flavifrons | Pseudomallada picteti | Pseudomallada prasinus | Rexa almerai |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Summer | Almond | * | 0.04 ± 0.02 | 0 | 0.04 ± 0.02 | 0.36 ± 0.09 | 0.04 ± 0.02 | 0 | * | 0.41 ± 0.13 | 0 |
| Oak | * | * | 0 | 0.04 ± 0.02 | 0.07 ± 0.03 | 0 | 0 | 0 | 0.32 ± 0.08 | 0 | |
| Olive | 0 | 0.05 ± 0.03 | * | 0.05 ± 0.03 | 0.28 ± 0.1 | 0 | 0 | 0 | 0.12 ± 0.04 | 0.05 ± 0.03 | |
| Pine | 0 | 0.03 ± 0.02 | 0.52 ± 0.25 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Autumn | Almond | 0 | 0 | * | 0 | * | 0 | 0 | 0 | 0.45 ± 0.14 | 0 |
| Oak | 0.03 ± 0.02 | 0 | 0 | * | 0 | 0 | 0 | * | 0.17 ± 0.05 | 0 | |
| Olive | 0 | 0 | 0 | 0.04 ± 0.02 | 0.12 ± 0.04 | 0 | 0 | * | 0.52 ± 0.1 | 0 | |
| Pine | 0 | 0 | 0 | 0 | 0 | 0 | 0 | * | * | 0 | |
| Winter | Almond | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.15 ± 0.05 | 0 |
| Oak | 0 | 0 | 0 | 0 | 0 | 0 | * | 0 | 0.09 ± 0.05 | 0 | |
| Olive | 0 | 0 | 0 | * | * | 0 | 0 | 0 | 0.25 ± 0.08 | 0 | |
| Pine | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | * | 0 | |
| Spring | Almond | 0 | 0.08 ± 0.04 | 0 | 0 | * | * | 0 | 0 | 0.25 ± 0.1 | 0 |
| Oak | 0 | 0 | 0 | 0 | 0.07 ± 0.03 | 0 | 0.04 ± 0.02 | 0.03 ± 0.02 | 0.13 ± 0.04 | 0 | |
| Olive | 0 | 0 | 0 | 0 | 0.05 ± 0.04 | 0 | 0 | 0 | 0.31 ± 0.08 | 0.08 ± 0.06 | |
| Pine | 0 | 0 | 0.29 ± 0.12 | 0 | 0 | 0 | * | * | * | 0 |
| Season | Tree Species | Juvenile Chrysopids Parasitized by | ||||
|---|---|---|---|---|---|---|
| Baryscapus impeditus | Gelis ilicicola | Helorus ruficornis | Isodromus puncticeps | Perilampus minutalis | ||
| Summer | Almond | 0.88 ± 0.23 | 0.03 ± 0.02 | 0 | 0.03 ± 0.02 | 0 |
| Oak | 0 | 0.05 ± 0.03 | 0.16 ± 0.05 | * | 0 | |
| Olive | 0 | * | * | * | 0 | |
| Pine | 0.15 ± 0.05 | 0 | 0.11 ± 0.05 | 0 | 0 | |
| Autumn | Almond | * | 0 | * | * | 0 |
| Oak | 0 | 0 | 0.17 ± 0.05 | 0 | 0 | |
| Olive | * | 0 | 0.07 ± 0.04 | 0 | 0.03 ± 0.02 | |
| Pine | 0.05 ± 0.04 | 0 | 0 | 0 | 0 | |
| Winter | Almond | 0 | 0 | 0 | 0 | 0 |
| Oak | 0 | 0 | 0.15 ± 0.05 | 0 | 0 | |
| Olive | 0 | 0 | 0.04 ± 0.02 | 0 | 0 | |
| Pine | 0 | * | 0 | 0 | 0 | |
| Spring | Almond | 0 | 0 | 0 | 0.04 ± 0.04 | * |
| Oak | * | 0 | 0.09 ± 0.04 | 0.04 ± 0.02 | * | |
| Olive | 0 | * | 0.04 ± 0.02 | 0 | * | |
| Pine | 0 | 0 | 0 | * | 0 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alcalá Herrera, R.; Campos, M.; González-Salvadó, M.; Ruano, F. Abundance and Population Decline Factors of Chrysopid Juveniles in Olive Groves and Adjacent Trees. Insects 2019, 10, 134. https://doi.org/10.3390/insects10050134
Alcalá Herrera R, Campos M, González-Salvadó M, Ruano F. Abundance and Population Decline Factors of Chrysopid Juveniles in Olive Groves and Adjacent Trees. Insects. 2019; 10(5):134. https://doi.org/10.3390/insects10050134
Chicago/Turabian StyleAlcalá Herrera, Rafael, Mercedes Campos, Marina González-Salvadó, and Francisca Ruano. 2019. "Abundance and Population Decline Factors of Chrysopid Juveniles in Olive Groves and Adjacent Trees" Insects 10, no. 5: 134. https://doi.org/10.3390/insects10050134
APA StyleAlcalá Herrera, R., Campos, M., González-Salvadó, M., & Ruano, F. (2019). Abundance and Population Decline Factors of Chrysopid Juveniles in Olive Groves and Adjacent Trees. Insects, 10(5), 134. https://doi.org/10.3390/insects10050134