Telenomus remus, a Candidate Parasitoid for the Biological Control of Spodoptera frugiperda in Africa, is already Present on the Continent

 , ,
, ,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Sampling
2.2. Morphological Analysis
2.3. Molecular Analyses
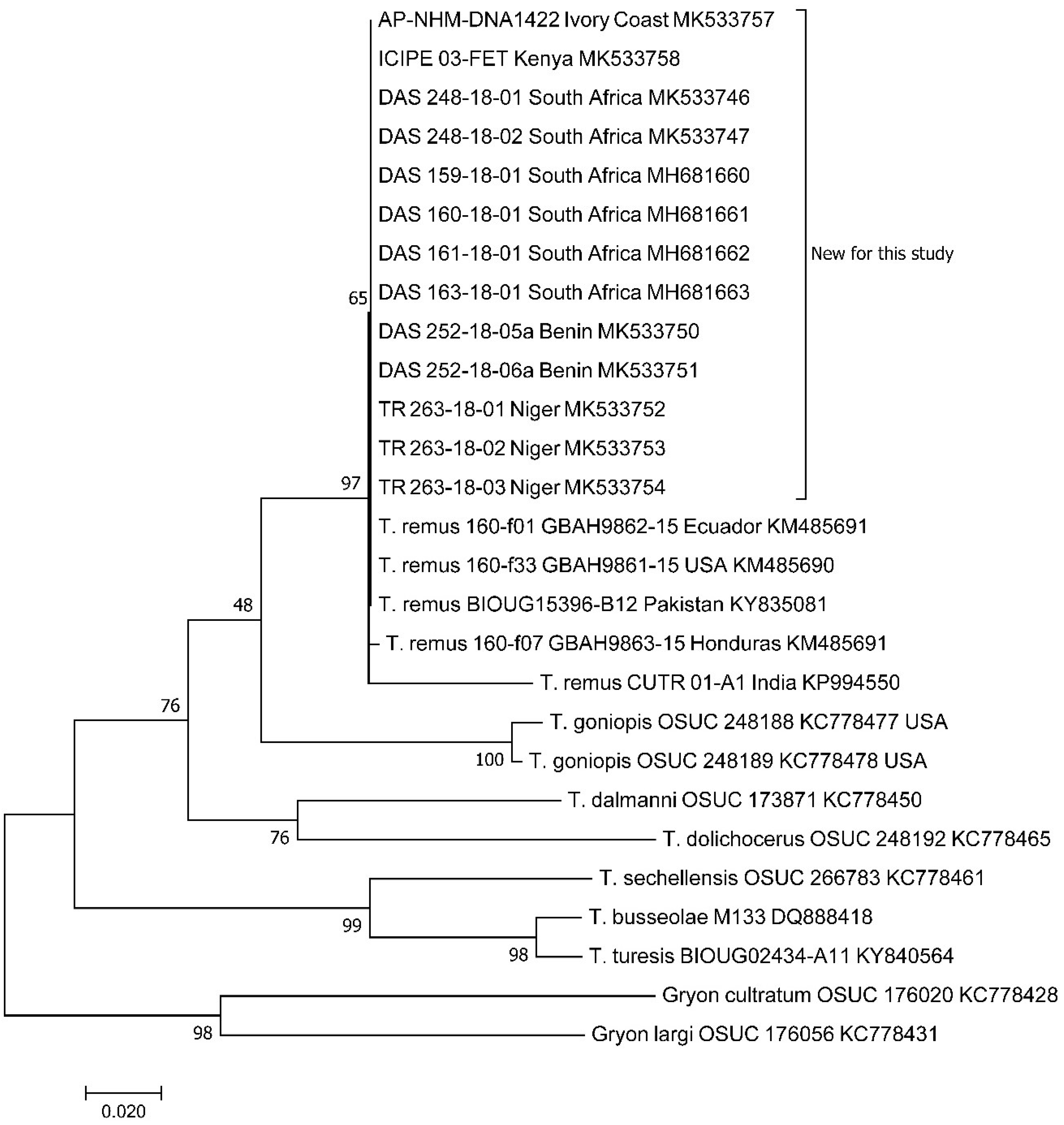
3. Results
4. Discussion
4.1. Presence of T. remus in Africa
4.2. Implications for the Biological Control of S. frugiperda
4.3. Taxonomy of Telenomus remus
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sparks, A.N. A review of the biology of the fall armyworm. Fla. Entomol. 1979, 62, 82–87. [Google Scholar] [CrossRef]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11. [Google Scholar] [CrossRef]
- FAO. Briefing Note on FAO Actions on Fall Armyworm in Africa; FAO: Rome, Italy; Available online: www.fao.org/3/a-bt415e.pdf (accessed on 20 February 2019).
- Du Plessis, H.; Van den Berg, J.; Kriticos, D.J.; Ota, N. Spodoptera frugiperda. (Fall Armyworm), CLIMEX modelling. CSIRO-InSTePP Pest Geography 2018. ResearchGate. Available online: https://www.researchgate.net/publication/327423712_Spodoptera_frugiperde_-_CLIMEX_modelling_du_Plessis_van_den_Berg_Ota_Kriticos_2018_CSIRO-InSTePP_Pest_Geography_June_2018_Canberra (accessed on 26 March 2019).
- Early, R.; González-Moreno, P.; Murphy, S.T.; Day, R. Forecasting the global extent of invasion of the cereal pest Spodoptera frugiperda, the fall armyworm. NeoBiota 2018, 40, 25–50. [Google Scholar] [CrossRef]
- Ganiger, P.C.; Yeshwant, H.M.; Muralimohan, K.; Vinay, N.; Kumar, A.R.V.; Chandrashekara, K. Occurrence of the new invasive pest, fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae), in the maize fields of Karnataka, India. Current Sci. 2018, 115, 621–623. [Google Scholar]
- Sharanabasappa, D.; Kalleshwaraswamy, C.M.; Asokan, R.; Mahadeva Swamy, H.M.; Marutid, M.S.; Pavithra, H.B.; Hegde, K.; Navi, S.; Prabhu, S.T.; Goergen, G. First report of the fall armyworm, Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae), an alien invasive pest on maize in India. Pest. Manage. Hort. Ecosys. 2018, 4, 23–29. [Google Scholar]
- CABI Fall Armyworm Portal. Available online: https://www.cabi.org/isc/fallarmyworm (accessed on 26 March 2019).
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall armyworm: Impacts and implications for Africa. Outlooks Pest Manage 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Prasanna, B.M.; Huesing, J.E.; Eddy, R.; Peschke, V.M. Fall Armyworm in Africa: A Guide for Integrated Pest Management, 1st ed.; CIMMYT: Mexico City, Mexico, 2018. [Google Scholar]
- Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Day, R.; Early, R.; Godwin, J.; et al. Fall Armyworm: Impacts and Implications for Africa. Evidence Note (2); Report to DFID. Available online: https://www.cabi.org/ISC/FullTextPDF/2018/20187200428.pdf (accessed on 26 March 2019).
- Yu, S.J.; Nguyen, S.N.; Abo-Elghar, G.E. Biochemical characteristics of insecticide resistance in the fall armyworm, Spodoptera frugiperda (J.E. Smith). Pest. Biochem. Phys. 2003, 77, 1–11. [Google Scholar] [CrossRef]
- Meagher, R.L.; Nuessly, G.S.; Nagoshi, R.N.; Hay-Roe, M.N. Parasitoids attacking fall armyworm (Lepidoptera: Noctuidae) in sweet corn habitats. Biol. Control 2016, 95, 66–72. [Google Scholar] [CrossRef]
- Van Driesche, R.G.; Bellows, T.S. Biological Control; Chapman and Hall: New York, NY, USA, 1996. [Google Scholar]
- Kenis, M.; Hurley, B.; Hajek, A.E.; Cock, M. Classical biological control of insect pests of trees—Facts and figures. Biol. Invasions 2017, 19, 3401–3417. [Google Scholar] [CrossRef]
- Molina-Ochoa, J.; Carpenter, J.E.; Heinrichs, E.A.; Foster, J.E. Parasitoids and parasites of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas and Caribbean Basin: An inventory. Fla. Entomol. 2003, 86, 254–289. [Google Scholar] [CrossRef]
- Estrada Vírgen, O.; Cambero Campos, J.; Robles Bermudez, A.; Rios Velasco, C.; Carvajal Cazola, C.; Isiordia Aquino, N.; Ruiz Cancino, E. Parasitoids and entomopathogens of the fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) in Nayarit, Mexico. Southwestern Entomol. 2013, 38, 339–344. [Google Scholar] [CrossRef]
- Cock, M.J.W.; Bennett, F.D.; Hughes, I.W.; Simmonds, F.J.; Yaseen, M. A Review of Biological Control of Pests in the Commonwealth Caribbean and Bermuda up to 1982; Commonwealth Agricultural Bureaux: Wallingford, Oxfordshire, UK, 1985. [Google Scholar]
- Lobo Lima, M.L.; van Harten, A. Luta biologica contra as pragas de cultures em Cabo Verde situaçao actual a programas futuros. Revista Investigaçáo Agraria CEA 1985, A1, 3–11. (In Portuguese) [Google Scholar]
- Cave, R.D. Biology, ecology and use in pest management of Telenomus remus. Biocontrol News Inf. 2000, 21, 21–26. [Google Scholar]
- Gutierrez-Martinez, A.; Tolon-Becerra, A.; Lastra-Bravo, X.B. Biological control of Spodoptera frugiperda eggs using Telenomus remus Nixon in maize-bean-squash polyculture. Am. J. Agr. Biol. Sci. 2012, 7, 285–292. [Google Scholar] [CrossRef]
- Pomari, A.F.; Bueno, A.D.F.; Bueno, R.C.O.D.F.; Junior, M.; de Oliveiras, A.; Fonseca, A.C.P.F. Releasing number of Telenomus remus (Nixon) (Hymenoptera: Platygastridae) against Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) in corn, cotton and soybean. Ciência Rural 2013, 43, 377–382. [Google Scholar] [CrossRef]
- Vieira, N.F.; Pomari-Fernandes, A.; Lemes, A.A.; Vacari, A.M.; De Bortoli, S.A.; de Freitas Bueno, A. Cost of production of Telenomus remus (Hymenoptera: Platygastridae) grown in natural and alternative hosts. J. Econ. Entomol. 2017, 110, 2724–2726. [Google Scholar] [CrossRef]
- Cave, R.D.; Acosta, N.M. Telenomus remus Nixon: Un parasitoide en el control biológico del gusano cogollero, Spodoptera frugiperda (Smith). Ceiba 1999, 40, 215–227. (In Spanish) [Google Scholar]
- Figueiredo, M.L.C.; Della Lucia, T.M.C.; Cruz, I. Effect of Telenomus remus Nixon (Hymenoptera: Scelionidae) density on control of Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) egg masses upon release in maize field. Rev. Bras. Milho Sorgo 2002, 1, 12–19. [Google Scholar] [CrossRef]
- Pomari-Fernandes, A.; De Queiroz, A.P.; de Freitas Bueno, A.; Sanzovo, A.W.; De Bortoli, S.A. The importance of relative humidity for Telenomus remus (Hymenoptera: Platygastridae) parasitism and development on Corcyra cephalonica (Lepidoptera: Pyralidae) and Spodoptera frugiperda (Lepidoptera: Noctuidae) eggs. Ann. Entomol. Soc. Am. 2015, 108, 11–17. [Google Scholar] [CrossRef]
- Polaszek, A.; Kimani, S.W. Telenomus species (Hymenoptera: Scelionidae) attacking eggs of pyralid pests (Lepidoptera) in Africa: A review and guide to identification. Bull. Entomol. Res. 1990, 80, 57–71. [Google Scholar] [CrossRef]
- Polaszek, A.; Ayshford, T.; Yahya, B.E.; Fusu, L. Wallaceaphytis: An unusual new genus of parasitoid wasp (Hymenoptera: Aphelinidae) from Borneo. J. Nat. Hist. 2014, 48, 1111–1123. [Google Scholar] [CrossRef]
- Cock, M.J.W.; Beseh, P.K.; Buddie, A.G.; Cafà, G.; Crozier, J. Molecular methods to detect Spodoptera frugiperda in Ghana, and implications for monitoring spread of invasive species in developing countries. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotech. 1994, 3, 294–299. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evo. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Ratnasingham, S.; Hebert, P.D.N. A DNA-based registry for all animal species: The Barcode Index Number (BIN) System. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2016, 44, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Hymenoptera Online. Available online: https://hol.osu.edu/index.html?id=3064 (accessed on 14 December 2018).
- Hymenoptera Online. Available online: https://hol.osu.edu/spmInfo.html?id=OSUC%2072859 (accessed on 14 December 2018).
- Hay-Roe, M.M.; Nagoshi, R.N.; Meagher, R.L.; De Lopez, M.A.; Trabanino, R. Isolation and DNA barcode characterization of a permanent Telenomus (Hymenoptera: Platygastridae) population in Florida that targets fall armyworm (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 2015, 108, 729–735. [Google Scholar] [CrossRef]
- Virgilio, M.; Backeljau, T.; Nevado, B.; De Meyer, M. Comparative performances of DNA barcoding across insect orders. BMC bioinformatics 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- Bowers, K.R. Telenomus podisi: One Species, or More? Master’s Thesis, The Ohio State University, Columbus, OH, USA, 2015. [Google Scholar]
- Wojcik, B.; Whitcomb, W.H.; Habeck, D.H. Host range testing of Telenomus remus (Hymenoptera: Scelionidae). Fla. Entomol. 1976, 59, 195–198. [Google Scholar] [CrossRef]
- Sisay, B.; Simiyu, J.; Malusi, P.; Likhayo, P.; Mendesil, E.; Elibariki, N.; Wakgari, M.; Ayalew, G.; Tefera, T. First report of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), natural enemies from Africa. J. Appl. Entomol. 2019. [Google Scholar] [CrossRef]
- Lesieur, V.; Lombaert, E.; Guillemaud, T.; Courtial, B.; Strong, W.; Roques, A.; Auger-Rozenberg, M.A. The rapid spread of Leptoglossus occidentalis in Europe: A bridgehead invasion. J. Pest. Sci. 2018, 92, 198–200. [Google Scholar] [CrossRef]
- Polaszek, A.; Kimani-Njogu, S.W. Scelionidae. In African Cereal Stem Borers: Economic Importance, Taxonomy, Natural Enemies and Control; Polaszek, A., Ed.; CAB International: Wallingford, UK, 1998; pp. 259–264. [Google Scholar]
- Gerling, D. The developmental biology of Telenomus remus Nixon (Hym.: Scelionidae). Bull. Entomol. Res. 1972, 61, 385–488. [Google Scholar] [CrossRef]
- Bueno, R.C.O.D.F.; Carneiro, T.R.; Pratissoli, D.; Bueno, A.D.; Fernandes, O.A. Biology and thermal requirements of Telenomus remus reared on fall armyworm Spodoptera frugiperda eggs. Ciência Rural 2008, 38, 1–6. [Google Scholar] [CrossRef]
- Pomari, A.F.; Bueno, A.D.F.; Bueno, R.C.O.D.F.; Menezes, A.D.O. Biological characteristics and thermal requirements of the biological control agent Telenomus remus (Hymenoptera: Platygastridae) reared on eggs of different species of the genus Spodoptera (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 2012, 105, 73–81. [Google Scholar] [CrossRef]
- Queiroz, A.P.; de Freitas Bueno, A.; Pomari-Fernandes, A.; Grande, M.L.M.; Bortolotto, O.C.; da Silva, D.M. Quality control of Telenomus remus (Hymenoptera: Platygastridae) reared on the factitious host Corcyra cephalonica (Lepidoptera: Pyralidae) for successive generations. Bull. Entomol. Res. 2017, 107, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Nixon, G.E.J. LIV.—Some Asiatic Telenominæ (Hym., Proctotrupoidea). Annals Mag. Natl. Hist. 1937, 20, 444–475. [Google Scholar] [CrossRef]
- Ashmead, W.H. Descriptions of new Hymenoptera from Japan. JSTOR 1904, 12, 65–84. [Google Scholar] [CrossRef]
- Risbec, J.I. La faune entomologique des cultures au Sénégal et au Soudan Français & II Contribution á l’étude des Proctotrupidae; Travaux du Laboratoire d’Entomologie du Secteur Soudanais de Recherches, Gouvernment Général de l’Afrique Occidentale Française: Paris, France, 1950. (In French) [Google Scholar]
- Raveendranath, S. Biology and behaviour of Telenomus spp. (Hymenoptera: Scelionidae) egg parasitoids, attacking Spodoptera spp. (Lepidoptera: Noctuidae). Ph.D. Thesis, Imperial College, University of London, London, UK, 1987. [Google Scholar]
- Arakaki, N.; Noda, H.; Yamagishi, K. Wolbachia-induced parthenogenesis in the egg parasitoid Telenomus nawai. Entomol. Exp. Appl. 2000, 96, 177–184. [Google Scholar] [CrossRef]


{kind=link}
| Country | Locality (Province) | Coordinates | Collection Date | Host Plant | No. Barcoded | Code (as in Figure 1) |
|---|---|---|---|---|---|---|
| South Africa | Mbombela (Mpumalanga) | 25.442149°S 30.992122°E | 20.04.18 | Maize | 4 | DAS 160-18-01 DAS 161-18-01 DAS 163-18-01 DAS-248-18-01 |
| Malelane (Mpumalanga) | 25.595231°S 31.665183°E | 20.04.18 | Maize | 2 | DAS-159-18-01 248-248-18-02 | |
| Côte d’Ivoire | Yamoussoukro (Kami) | 6.875833°N 5.363333°W | 25.05.18 | Maize | 1 | AP-NHM-DNA1422 |
| Niger | Maradi (Djiratawa) | 13.2360°N 7.0760°E | 15.08.17 | Maize | 0 | 1455 (NHM) |
| Tilabéri (Sadoré) | 13.2454°N 2.3047°E | 18.09.18 | Sorghum | 3 | TR 263-18-01 TR 263-18-02 TR 263-18-03 | |
| Benin | Abomey Calavi (Atlantique) | 6.4375°N 2.3283°E | 04.07.18 | Maize | 1 | DAS 252-18-05a |
| Abomey Calavi (Atlantique) | 6.43064°N 2.29544°E | 01.10.18 | Maize | 1 | DAS 252-18-6a | |
| Kenya | Kilifi (Kilifi) | 3.51.06°S 39.9092°E | 01.10.18 | Maize | 1 | 03-FET |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kenis, M.; du Plessis, H.; Van den Berg, J.; Ba, M.N.; Goergen, G.; Kwadjo, K.E.; Baoua, I.; Tefera, T.; Buddie, A.; Cafà, G.; et al. Telenomus remus, a Candidate Parasitoid for the Biological Control of Spodoptera frugiperda in Africa, is already Present on the Continent. Insects 2019, 10, 92. https://doi.org/10.3390/insects10040092
Kenis M, du Plessis H, Van den Berg J, Ba MN, Goergen G, Kwadjo KE, Baoua I, Tefera T, Buddie A, Cafà G, et al. Telenomus remus, a Candidate Parasitoid for the Biological Control of Spodoptera frugiperda in Africa, is already Present on the Continent. Insects. 2019; 10(4):92. https://doi.org/10.3390/insects10040092
Chicago/Turabian StyleKenis, Marc, Hannalene du Plessis, Johnnie Van den Berg, Malick Niango Ba, Georg Goergen, Koffi Eric Kwadjo, Ibrahim Baoua, Tadele Tefera, Alan Buddie, Giovanni Cafà, and et al. 2019. "Telenomus remus, a Candidate Parasitoid for the Biological Control of Spodoptera frugiperda in Africa, is already Present on the Continent" Insects 10, no. 4: 92. https://doi.org/10.3390/insects10040092
APA StyleKenis, M., du Plessis, H., Van den Berg, J., Ba, M. N., Goergen, G., Kwadjo, K. E., Baoua, I., Tefera, T., Buddie, A., Cafà, G., Offord, L., Rwomushana, I., & Polaszek, A. (2019). Telenomus remus, a Candidate Parasitoid for the Biological Control of Spodoptera frugiperda in Africa, is already Present on the Continent. Insects, 10(4), 92. https://doi.org/10.3390/insects10040092