Field and Laboratory Performance of False Codling Moth, Thaumatotibia Leucotreta (Lepidoptera: Troticidae) on Orange and Selected Vegetables

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
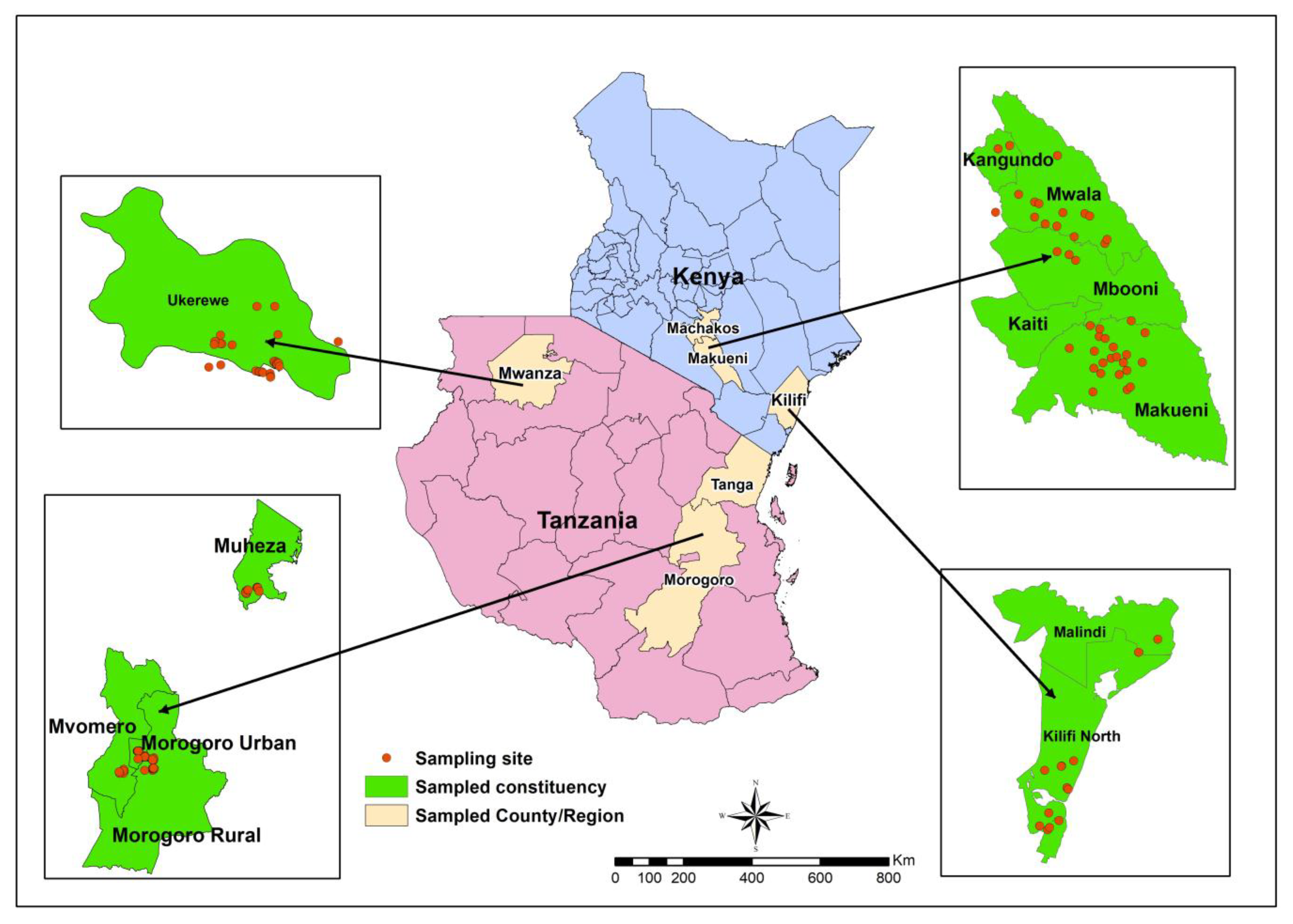
2.1. Study Sites
2.2. Assessment of Thaumatotibia leucotreta Incidence on Orange in Kenya and Tanzania
2.3. Assessment of Thaumatotibia leucotreta Incidence in Vegetables from Morogoro, Tanzania
2.4. Laboratory Performance of Thaumatotibia leucotreta on Orange and Solanaceous Vegetables
2.4.1. Insect Cultures and Host
2.4.2. Oviposition Preference of Thaumatotibia leucotreta for Orange and Solanaceous Vegetables
Choice Test
No-choice Test
2.5. Developmental Duration and Larval Mass of Thaumatotibia leucotreta Reared on Orange and Solanaceous Vegetables
2.5.1. Insect Colonies and Hosts
2.5.2. Egg Stage
2.5.3. Larval Stage
2.5.4. Pre-Pupal Stage
2.5.5. Pupal Stage
2.5.6. Survival of Thaumatotibia leucotreta Reared on Orange and Solanaceous Vegetables
2.5.7. Adult Life History of Thaumatotibia leucotreta Reared on Orange and Solanaceous Vegetables
2.6. Data Analysis
3. Results
3.1. Percentage of Orange Fruit with Thaumatotibia leucotreta Damage Symptoms and Larvae
3.2. Incidence of Thaumatotibia leucotreta Larvae in Vegetables
3.3. Oviposition Preference of Thaumatotibia leucotreta for Orange and Solanaceous Vegetables
3.3.1. Choice Test
3.3.2. No-choice Test
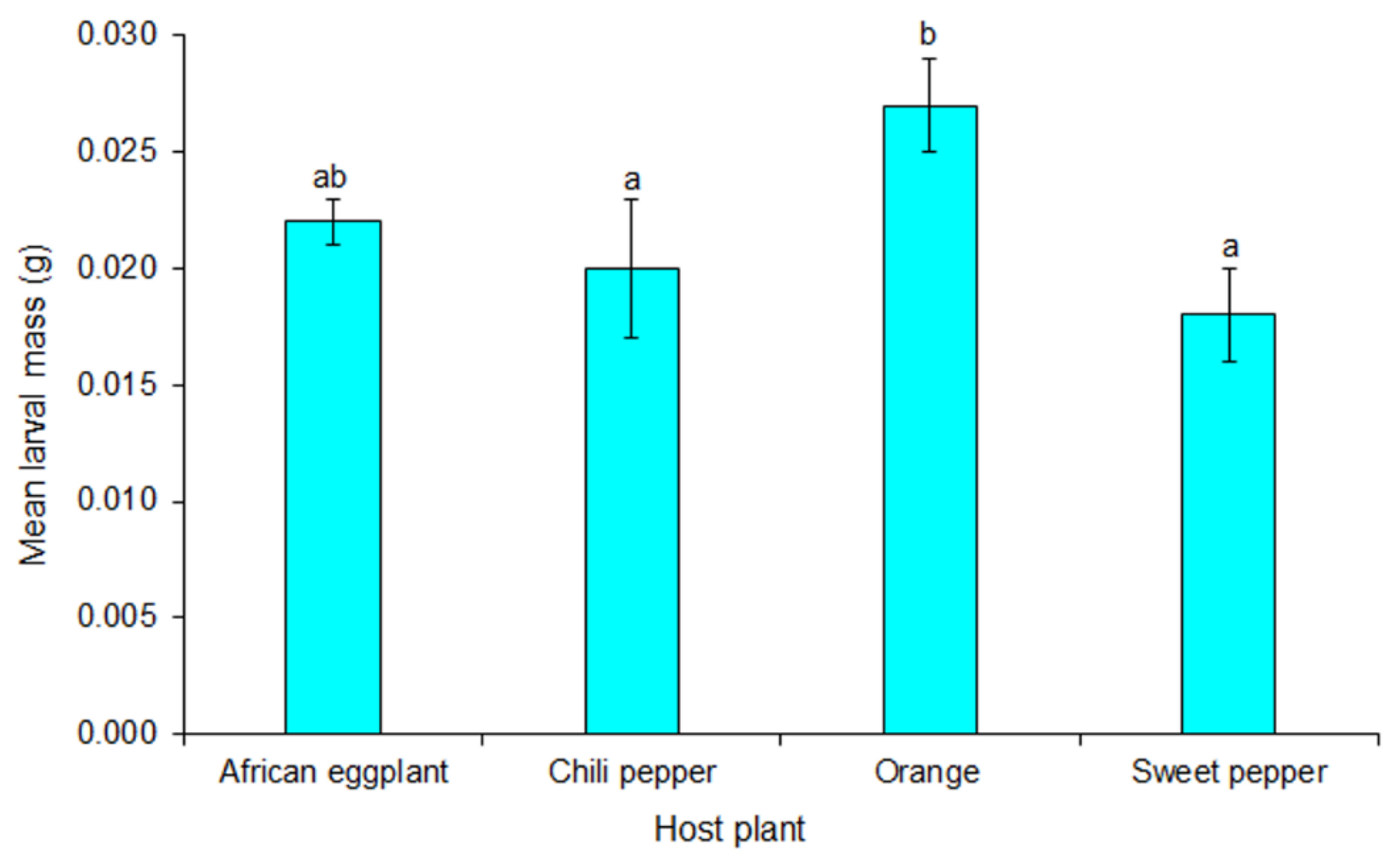
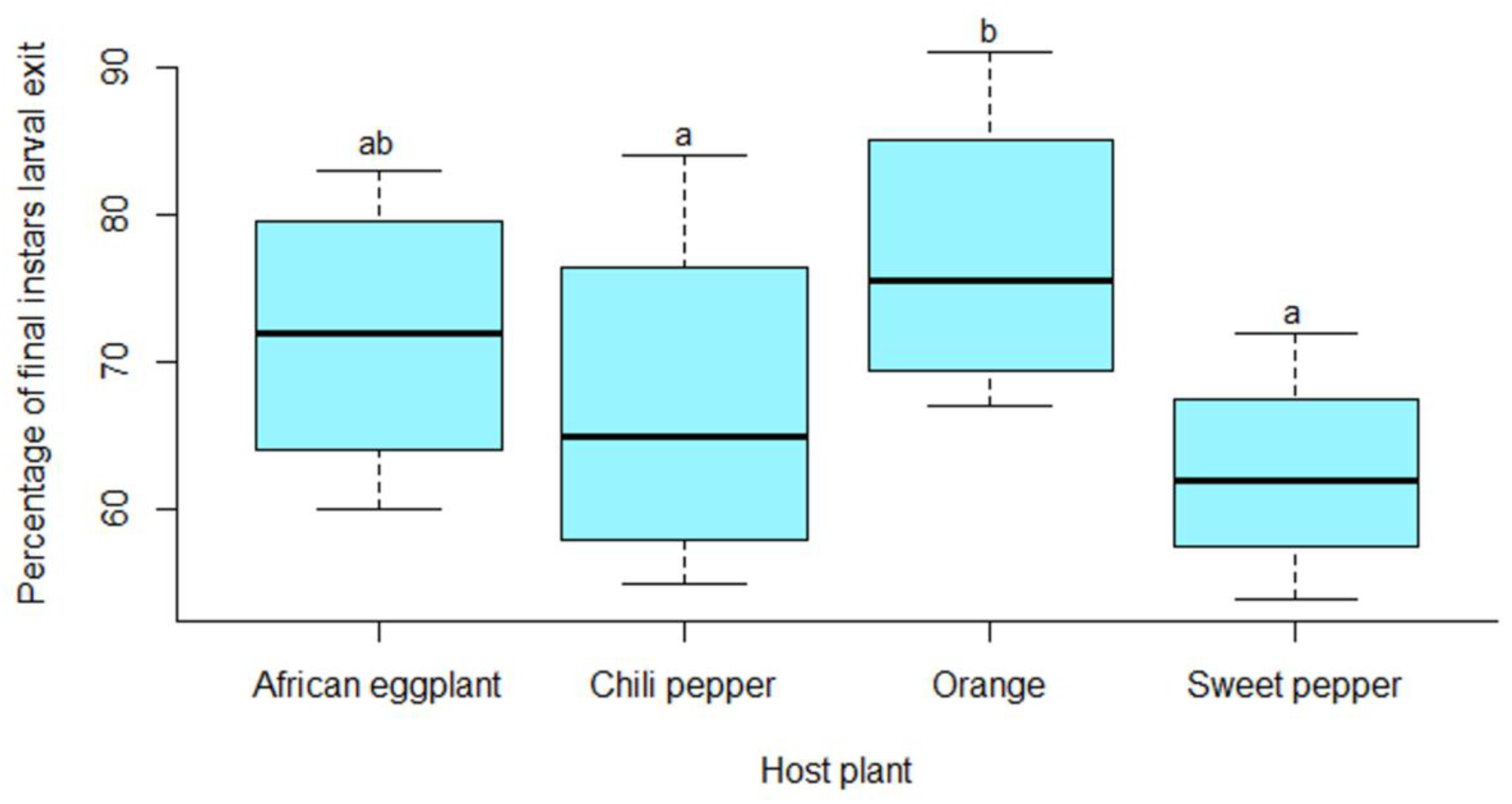
3.4. Development Time of Thaumatotibia leucotreta Immature Stages and Final Larval Instar Mass when Reared on Orange and Solanaceous Vegetables
3.5. Development and Survival of Thaumatotibia leucotreta Immature Stages Reared on Orange and Solanaceous Vegetables
3.6. Reproductive Parameters for Thaumatotibia leucotreta Reared on Orange and Solanaceous Vegetables
3.6.1. Fecundity per Female per Day
3.6.2. Pairwise Comparison on Fecundity Rate of Thaumatotibia leucotreta Reared from Orange and Solanaceous Hosts
3.7. Life History Parameters of Thaumatotibia leucotreta Reared from Orange and Solanaceous Vegetables
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gilligan, T.M.; Epstein, M.E.; Hoffman, K.M. Discovery of false codling moth, Thaumatotibia leucotreta (Meyrick), in California (Lepidoptera: Tortricidae). Proc. Entomol. Soc. Wash. 2011, 113, 426–435. [Google Scholar] [CrossRef]
- Kirkman, W.; Moore, S. A study of alternative hosts for the false codling moth, Thaumatotibia (=Cryptophlebia) leucotreta in the Eastern Cape. SA Fruit J. 2007, 6, 33–38. [Google Scholar]
- Malan, A.P.; Knoetze, R.; Moore, S.D. Isolation and identification of entomopathogenic nematodes from citrus orchards in South Africa and their biocontrol potential against false codling moth. J. Invertebr. Pathol. 2011, 108, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Mazza, G.; Strangi, A.; Marinelli, L.; Del Nista, D.; Roversi, P.F. Thaumatotibia leucotreta (Meyrick) (Lepidoptera Tortricidae) intercepted for the first time in Italy. Redia 2014, 97, 147–149. [Google Scholar]
- Erichsen, C.; Schoeman, A.S. Moth pests of avocados. S. Afr. Avocado Grow. Assoc. Yearb. 1994, 17, 109–112. [Google Scholar]
- Venette, R.C.; Davis, E.E.; DaCosta, M.; Heisler, H.; Larson, M. Mini Risk Assessment: False Codling Moth, Thaumatotibia (=Cryptophlebia) leucotreta (Meyrick) [Lepidoptera: Tortricidae]; University of Minnesota, Department of Entomology: Falcon Heights, MN, USA, 2003. [Google Scholar]
- Boardman, L.; Grout, T.G.; Terblanche, J.S. False codling moth Thaumatotibia leucotreta (Lepidoptera, Tortricidae) larvae are chill-susceptible. Insect Sci. 2012, 19, 315–328. [Google Scholar] [CrossRef]
- Daiber, C.C. A study of the biology of the false codling moth Cryptophlebia leucotreta (Meyr.): The adult and generations during the year. Phytophylactica 1980, 12, 187–193. [Google Scholar]
- Moore, S.D. Moths and butterflies: False codling moth. Citrus Res. Int. IPM Prod. Guidel. 2012, 3, 1–9. [Google Scholar]
- Moore, S.D.; Kirkman, W. Citrus orchard sanitation with emphasis on false codling moth control. SA Fruit J. 2008, 7, 57–60. [Google Scholar]
- Makorere, R. An exploration of factors affecting development of citrus industry in Tanzania: Empirical evidence from Muheza District, Tanga Region. Int. J. Food Agric. Econ. 2014, 2, 135–154. [Google Scholar]
- Mounde, L.G.; Ateka, E.M.; Kihurani, A.W.; Wasilwa, L.; Thuranira, E.G. Occurrence and distribution of citrus gummosis (Phytophthora spp.) in Kenya. Afr. J. Hortic. Sci. 2009, 2, 315–328. [Google Scholar]
- Odanga, J.; Mohamed, S.; Mwalusepo, S.; Olubayo, F.; Nyankanga, R.; Khamis, F.; Rwomushana, I.; Johansson, T.; Ekesi, S. Spatial Distribution of Bactrocera dorsalis and Thaumatotibia leucotreta in Smallholder Avocado Orchards along Altitudinal Gradient of Taita Hills and Mount Kilimanjaro. Insects 2018, 9, 71. [Google Scholar] [CrossRef] [PubMed]
- Brehm, G.; Fiedler, K. Faunal composition of geometrid moths changes with altitude in an Andean montane rain forest. J. Biogeogr. 2003, 30, 431–440. [Google Scholar] [CrossRef]
- Hilt, N.; Fiedler, K. Arctiid moth ensembles along a successional gradient in the Ecuadorian montane rain forest zone: How different are subfamilies and tribes? J. Biogeogr. 2006, 33, 108–120. [Google Scholar] [CrossRef]
- Ashton, L.A.; Odell, E.H.; Burwell, C.J.; Maunsell, S.C.; Nakamura, A.; McDonald, W.J.F.; Kitching, R.L. Altitudinal patterns of moth diversity in tropical and subtropical Australian rainforests. Austral. Ecology 2016, 41, 197–208. [Google Scholar] [CrossRef]
- Highland, S.A.; Miller, J.C.; Jones, J.A. Determinants of moth diversity and community in a temperate mountain landscape: Vegetation, topography, and seasonality. Ecosphere 2013, 4, 1–22. [Google Scholar] [CrossRef]
- Love, C.N.; Hill, M.P.; Moore, S.D. Thaumatotibia leucotreta and the Navel orange: Ovipositional preferences and host susceptibility. J. Appl. Entomol. 2014, 138, 600–611. [Google Scholar] [CrossRef]
- Thompson, J.N.; Pellmyr, O. Evolution of oviposition behavior and host preference in Lepidoptera. Annu. Rev. Entomol. 1991, 36, 65–89. [Google Scholar] [CrossRef]
- Daiber, C.C. A study of the biology of the false codling moth [Cryptophlebia leucotreta (Meyr.)]: The cocoon. Phytophylactica 1979, 11, 151–157. [Google Scholar]
- Daiber, C.C. A study of the biology of the false codling moth [Cryptophlebia leucotreta (Meyr.)]: The egg. Phytophylactica 1979, 11, 129–132. [Google Scholar]
- Daiber, C.C. A study of the biology of the false codling moth [Cryptophlebia leucotreta (Meyr.)]: The larva. Phytophylactica 1979, 11, 141–144. [Google Scholar]
- De Jager, Z.M. Biology and Ecology of the False Codling Moth, Thaumatotibia leucotreta (Meyrick). Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, 2013. [Google Scholar]
- Moore, S.D.; Richards, G.I.; Chambers, C.; Hendry, D. An improved larval diet for commercial mass rearing of the false codling moth, Thaumatotibia leucotreta (Meyrick) (Lepidoptera: Tortricidae). Afr. Entomol. 2014, 22, 216–219. [Google Scholar] [CrossRef]
- Opoku-Debrah, J.K.; Hill, M.P.; Knox, C.; Moore, S.D. Comparison of the biology of geographically distinct populations of the citrus pest, Thaumatotibia leucotreta (Meyrick) (Lepidoptera: Tortricidae), in South Africa. Afr. Entomol. 2014, 22, 530–537. [Google Scholar] [CrossRef]
- Haile, A.; Hofsvang, T. Host plant preference of the stem borer Busseola fusca (Fuller) (Lepidoptera: Noctuidae). Crop Prot. 2002, 21, 227–233. [Google Scholar] [CrossRef]
- Mkiga, A.M.; Mwatawala, M.W. Developmental Biology of Zeugodacus cucurbitae (Diptera: Tephritidae) in Three Cucurbitaceous Hosts at Different Temperature Regimes. J. Insect Sci. 2015, 15. [Google Scholar] [CrossRef]
- Carey, J.R. Applied Demography for Biologists: With Special Emphasis on Insects; Oxford University Press: Oxford, UK, 1993. [Google Scholar]
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Hofmeyr, J.H.; Pringle, K.L. Resistance of false codling moth, Cryptophlebia leucotreta (Meyrick) (Lepidoptera: Tortricidae), to the chitin synthesis inhibitor, triflumuron. Afr. Entomol. 1998, 6, 373–375. [Google Scholar]
- Catung, H.A. Population studies of the false codling moth, Cryptophlebia lellcolrela Meyr., on citrus in the Transvaal. Phytophylactica 1974, 6, 31–38. [Google Scholar]
- Jaenike, J. On optimal oviposition behavior in phytophagous insects. Theor. Popul. Biol. 1978, 14, 350–356. [Google Scholar] [CrossRef]
- Newton, P.J. Inversely density-dependent egg parasitism in patchy distributions of the citrus pest Cryptophlebia leucotreta (Lepidoptera: Tortricidae) and its agricultural efficiency. J. Appl. Ecol. 1988, 25, 145–162. [Google Scholar] [CrossRef]
- Gripenberg, S.; Mayhew, P.J.; Parnell, M.; Roslin, T. A meta-analysis of preference–performance relationships in phytophagous insects. Ecol. Lett. 2010, 13, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Muthuthantri, S.; Clarke, A.R. Five commercial citrus rate poorly as hosts of the polyphagous fruit fly Bactrocera tryoni (Froggatt) (D iptera: Tephritidae) in laboratory studies. Aust. J. Entomol. 2012, 51, 289–298. [Google Scholar] [CrossRef]
- König, M.A.; Wiklund, C.; Ehrlén, J. Butterfly oviposition preference is not related to larval performance on a polyploid herb. Ecol. Evol. 2016, 6, 2781–2789. [Google Scholar] [CrossRef] [PubMed]
- Hanif, R.; Iqbal, Z.; Iqbal, M.; Hanif, S.; Rasheed, M. Use of vegetables as nutritional food: Role in human health. J. Agric. Biol. Sci. 2006, 1, 18–22. [Google Scholar]
- Eze, S.O.; Kanu, C.Q. Phytochemical and nutritive composition analysis of Solanum aethopicum L. J. Pharm. Sci. Innov. 2014, 4, 358–361. [Google Scholar] [CrossRef]
- Ananthan, R.; Subash, K.; Longvah, T. Assessment of nutrient composition and capsaicinoid content of some red chilies. Int. Food Nutr. Technol. 2014, 72, 1–4. [Google Scholar]
- Traore, F.; Dabire-Binso, C.L.; Ba, N.M.; Sanon, A.; Pittendrigh, B.R. Feeding preferences of the legume pod borer Maruca vitrata (Lepidoptera: Crambidae) larvae and suitability of different flower parts for larval development. Int. J. Trop. Insect Sci. 2013, 33, 107–113. [Google Scholar] [CrossRef]
- Gould, J.; Venette, R.; Winograd, D. Effect of temperature on development and population parameters of Copitarsia decolora (Lepidoptera: Noctuidae). Environ. Entomol. 2005, 34, 548–556. [Google Scholar] [CrossRef]
- Vargas, R.I.; Walsh, W.A.; Kanehisa, D.; Jang, E.B.; Armstrong, J.W. Demography of four Hawaiian fruit flies (Diptera: Tephritidae) reared at five constant temperatures. Ann. Entomol. Soc. Am. 1997, 90, 162–168. [Google Scholar] [CrossRef]
- Vayssières, J.F.; Carel, Y.; Coubes, M.; Duyck, P.F. Development of immature stages and comparative demography of two cucurbit-attacking fruit flies in Reunion Island: Bactrocera cucurbitae and Dacus ciliatus (Diptera Tephritidae). Environ. Entomol. 2008, 37, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Birch, L. The intrinsic rate of natural increase of an insect population. J. Anim. Ecol. 1948, 17, 15–26. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Village/Orchard | Latitude | Longitude | Vegetables Sampled Nearby the Orchard | Distance from the Orchard |
|---|---|---|---|---|
| Mlali | S06°56′55.3″ | E037°31′55.4″ | Sweet pepper, African eggplant, okra, chili pepper | 67–72 m |
| Mlali | S06°56′33.6″ | E037°31′59.1″ | Sweet pepper, African eggplant, okra, chili pepper | 69–75 m |
| Mlali | S06°57′20.5″ | E037°32′02.9″ | Sweet pepper, African eggplant, okra, chili pepper | 58–69 m |
| Mlali | S06°57′46.8″ | E037°32′14.1’′ | Sweet pepper, African eggplant, okra, chili pepper | 65–70 m |
| Mlali | S06°57′35.2″ | E037°31′51.8″ | Sweet pepper, African eggplant, okra, chili pepper | 68–74 m |
| Mkuyuni | S06°58′41.9″ | E037°31′58.0″ | Sweet pepper, African eggplant, okra, chili pepper | 63–69 m |
| Kipera | S06°56′14.3″ | E037°31′40.2″ | Sweet pepper, African eggplant, okra, chili pepper | 66–74 m |
| Vitonga | S06°57′15.9″ | E037°29′35.9″ | Sweet pepper, African eggplant, okra, chili pepper | 55–67 m |
| Vitonga | S06°58′36.8″ | E037°29′23.0″ | Sweet pepper, African eggplant, okra, chili pepper | 66–72 m |
| Vitonga | S06°57′52.4″ | E037°30′38.5″ | Sweet pepper, African eggplant, okra, chili pepper | 56–63 m |
| Parameter | Altitude | Fruit Sampling Method | Country | Average | |
|---|---|---|---|---|---|
| Kenya | Tanzania | ||||
| A% fruit with damage symptoms | >1201 masl | Ground | 42.16 ± 3.40 h | 33.92 ± 3.05 fgh | 38.04 ± 2.29 e |
| Tree without shaking | 12.72 ± 1.69 bcd | 9.36 ± 1.35 bc | 11.04 ± 1.10 b | ||
| Tree shaking | 28.48 ± 2.88 fg | 22.08 ± 2.12 def | 25.28 ± 1.75 c | ||
| 501–1200 masl | Ground | 40.24 ± 2.28 gh | 28.58 ± 2.72 fgh | 34.38 ± 1.91 de | |
| Tree without shaking | 7.72 ± 1.26 abc | 5.52 ± 1.96 bc | 6.62 ± 0.80 a | ||
| Tree branch shaking | 27.56 ± 2.20 efg | 21.40 ± 1.95 def | 24.48 ± 1.49 c | ||
| 0–500 masl | Ground | 35.88 ± 1.87 gh | 21.76 ± 2.49 def | 28.82 ± 1.82 cd | |
| Tree without shaking | 5.76 ± 1.21 ab | 2.88 ± 0.76 a | 4.32 ± 0.74 a | ||
| Tree shaking | 16.08 ± 1.82 cde | 14.16 ± 1.69 cd | 15.12 ± 1.21 b | ||
| Average | 24.07A | 17.73B | |||
| B% Larval infested fruit | |||||
| >1201 masl | Ground | 33.26 ± 3.59 efg | 21.00 ± 2.75 cde | 27.12 ± 2.41de | |
| Tree without shaking | 16.00 ± 0.99 bcd | 12.08 ± 0.75 abc | 14.04 ± 0.68 bc | ||
| Tree shaking | 46.48 ± 4.38 g | 41.80 ± 3.69 fg | 44.14 ± 2.85 g | ||
| 501–1200 masl | Ground | 29.00 ± 3.02 def | 16.40 ± 1.84 bcd | 22.62 ± 1.96 cd | |
| Tree without shaking | 14.93 ± 1.27 bc | 8.96 ± 0.63 abc | 11.90 ± 0.82 ab | ||
| Tree shaking | 45.04 ± 3.75 g | 37.44 ± 3.75 fg | 41.24 ± 2.64 fg | ||
| 0–500 masl | Ground | 15.46 ± 1.80 bc | 11.44 ± 1.27 abc | 13.40 ± 1.13 b | |
| Tree without shaking | 8.27 ± 0.83 ab | 4.92 ± 0.49 a | 6.64 ± 0.54 a | ||
| Tree shaking | 34.24 ± 3.99 efg | 33.00 ± 3.78 efg | 33.62 ±2.72 ef | ||
| Average | 26.93A | 20.79B | |||
| Host Plant | Scientific Name | Percentage Infestation |
|---|---|---|
| Okra | Abelmoschus esculentus | 3.08 ± 0.90 a |
| Sweet pepper | Capsicum spp. | 8.72 ± 1.81 b |
| African eggplant | Solanum aethiopicum | 12.00 ± 1.46 b |
| Chili pepper | Capsicum spp. | 10.96 ± 1.28 b |
| Parameter | Host Plant | |||
|---|---|---|---|---|
| Orange | Sweet Pepper | African Eggplant | Chili Pepper | |
| * Sex ratio (male: female) | 0.38:0.62 | 0.32:0.68 | 0.37:0.63 | 0.34:0.66 |
| * Pre-oviposition period | 1.20 ± 0.08 | 1.21 ± 0.09 | 1.17 ± 0.12 | 1.23 ± 0.06 |
| * Oviposition period | 8.00 ± 0.4 | 7.75 ± 0.29 | 8.00 ± 0.41 | 7.75 ± 0.25 |
| * Female longevity (days) | 16.02 ± 0.89 | 16.21 ± 0.71 | 16.61 ± 0.73 | 15.90 ± 0.82 |
| * Male longevity (days) | 15.67 ± 0.69 | 15.34 ± 0.60 | 15.51 ± 0.90 | 15.61 ± 0.81 |
| Fecundity | 415.00 ± 3.70c | 341.00 ± 5.37a | 378.00 ± 8.03b | 364.00 ± 3.53b |
| Orange | Chili Pepper | African Eggplant | Sweet Pepper | |
|---|---|---|---|---|
| Day 3 | ||||
| Orange | - | 0.823 | 1.000 | 0.460 |
| Chili pepper | - | 0.829 | 0.928 | |
| African eggplant | - | 0.459 | ||
| Day 4 | ||||
| Orange | - | 0.003 | 0.009 | 0.001 |
| Chili pepper | - | 0.988 | 0.822 | |
| African eggplant | - | 0.630 | ||
| Day 5 | ||||
| Orange | - | 0.6815 | 0.177 | 0.010 |
| Chili pepper | - | 1.000 | 0.182 | |
| African eggplant | - | 0.177 |
| Parameter | Host Plant | |||
|---|---|---|---|---|
| Orange | Sweet Pepper | African Eggplant | Chili Pepper | |
| Net reproductive rate | 419.7 ± 3.844c | 348.00 ± 5.484a | 385.90 ± 7.722b | 371.5 ± 3.611b |
| * Mean generation time | 48.19 ± 1.103 | 48.90 ± 1.301 | 47.29 ± 0.807 | 48.42 ± 0.261 |
| * Intrinsic rate of increase | 0.13 ± 0.002 | 0.12 ± 0.003 | 0.13 ± 0.002 | 0.12 ± 0.001 |
| * Doubling time | 5.53 ± 0.128 | 5.79 ± 0.162 | 5.50 ± 0.086 | 5.67 ± 0.024 |
| * Finite rate of increase | 1.40 ± 0.002 | 1.13 ± 0.004 | 1.13 ± 0.002 | 1.12 ± 0.002 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mkiga, A.M.; Mohamed, S.A.; du Plessis, H.; Khamis, F.M.; Ekesi, S. Field and Laboratory Performance of False Codling Moth, Thaumatotibia Leucotreta (Lepidoptera: Troticidae) on Orange and Selected Vegetables. Insects 2019, 10, 63. https://doi.org/10.3390/insects10030063
Mkiga AM, Mohamed SA, du Plessis H, Khamis FM, Ekesi S. Field and Laboratory Performance of False Codling Moth, Thaumatotibia Leucotreta (Lepidoptera: Troticidae) on Orange and Selected Vegetables. Insects. 2019; 10(3):63. https://doi.org/10.3390/insects10030063
Chicago/Turabian StyleMkiga, Abdullah Mohamed, Samira Abuelgasim Mohamed, Hannalene du Plessis, Fathiya Mbarak Khamis, and Sunday Ekesi. 2019. "Field and Laboratory Performance of False Codling Moth, Thaumatotibia Leucotreta (Lepidoptera: Troticidae) on Orange and Selected Vegetables" Insects 10, no. 3: 63. https://doi.org/10.3390/insects10030063
APA StyleMkiga, A. M., Mohamed, S. A., du Plessis, H., Khamis, F. M., & Ekesi, S. (2019). Field and Laboratory Performance of False Codling Moth, Thaumatotibia Leucotreta (Lepidoptera: Troticidae) on Orange and Selected Vegetables. Insects, 10(3), 63. https://doi.org/10.3390/insects10030063