Dynamic Responses of Ground-Dwelling Invertebrate Communities to Disturbance in Forest Ecosystems


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Disturbance in Forest Ecosystems
3. Responses of Ground-Dwelling Invertebrates to Patterns of Forest Disturbance
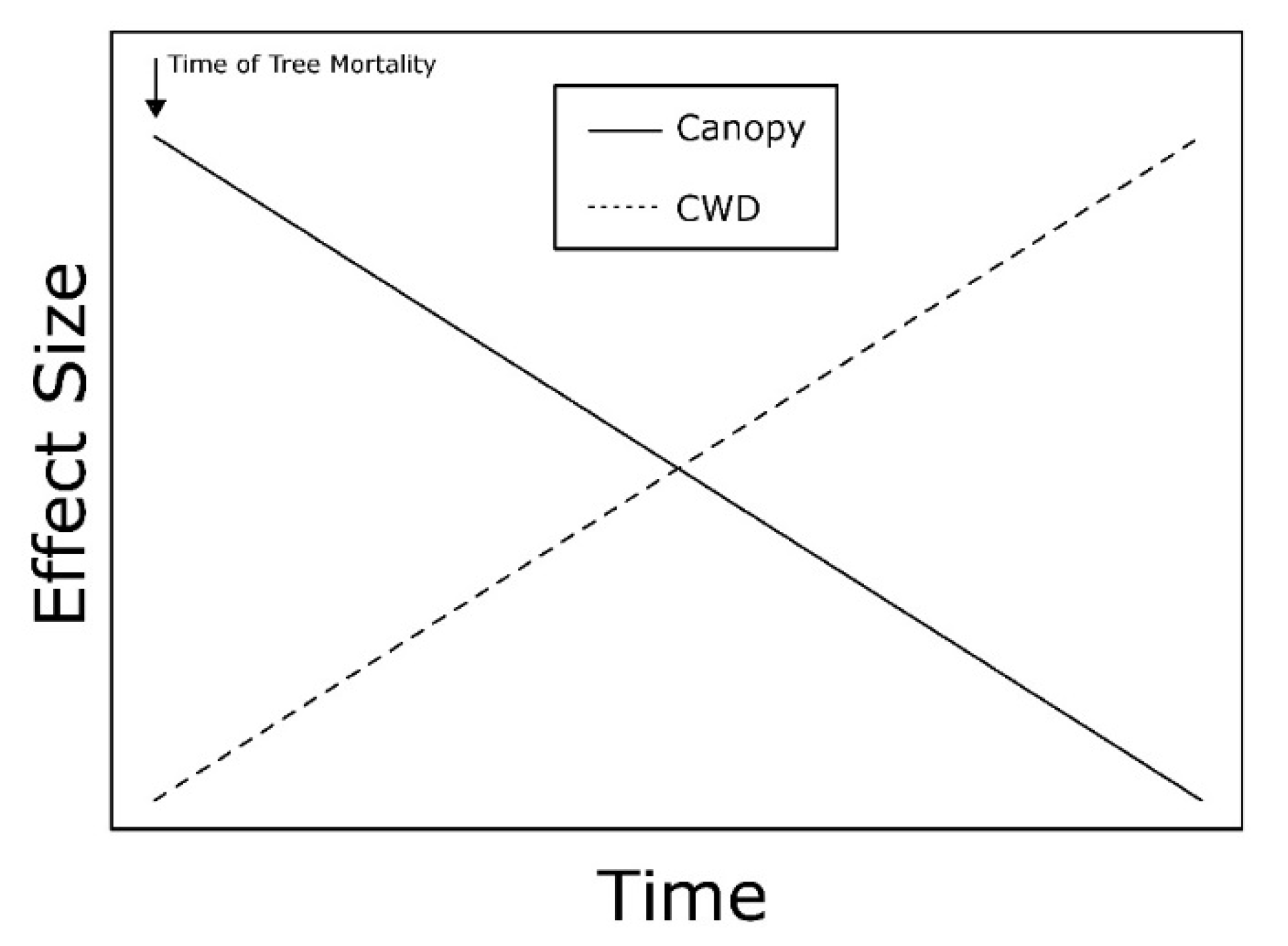
3.1. Invasive Insects
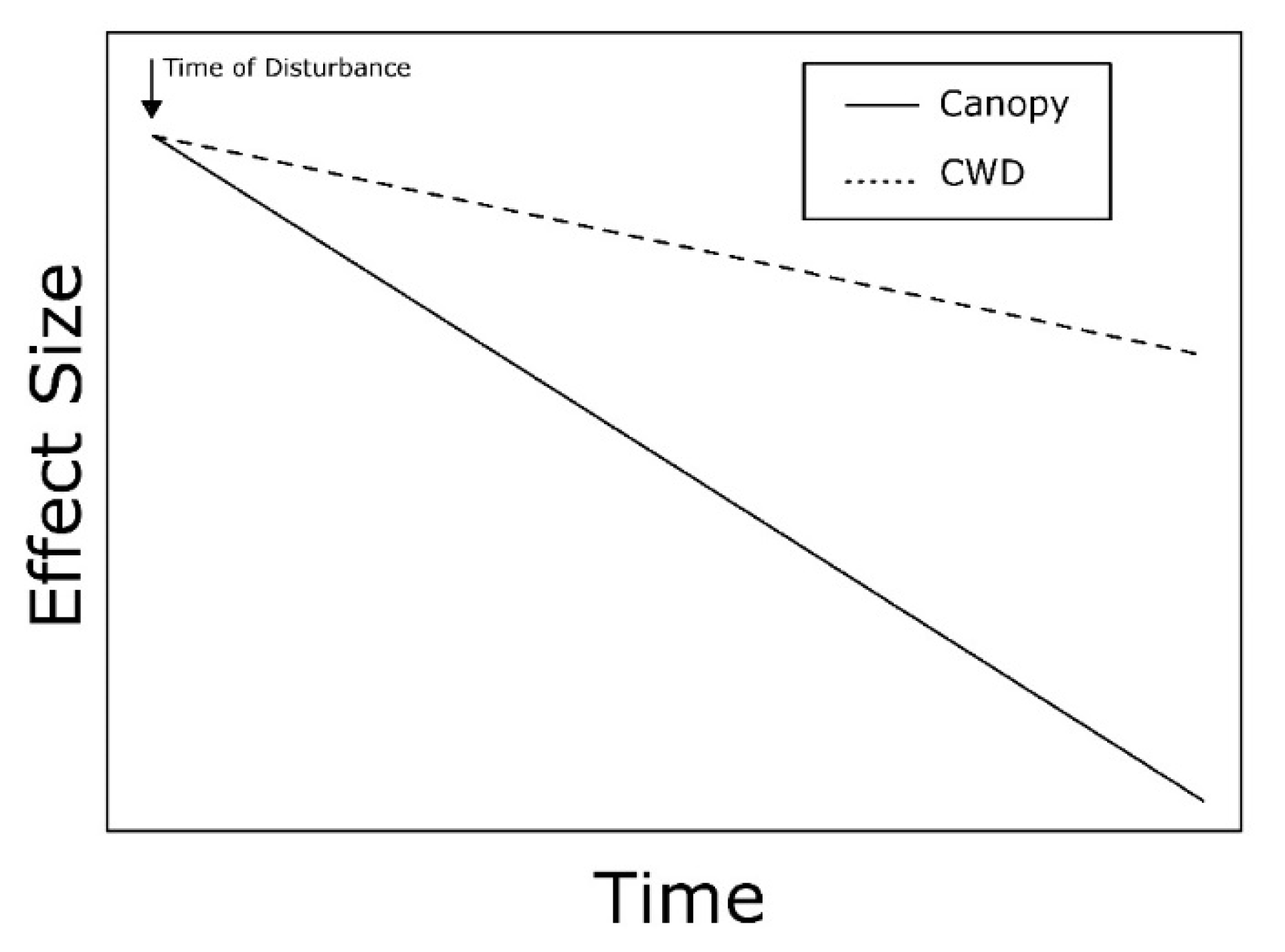
3.2. Wind
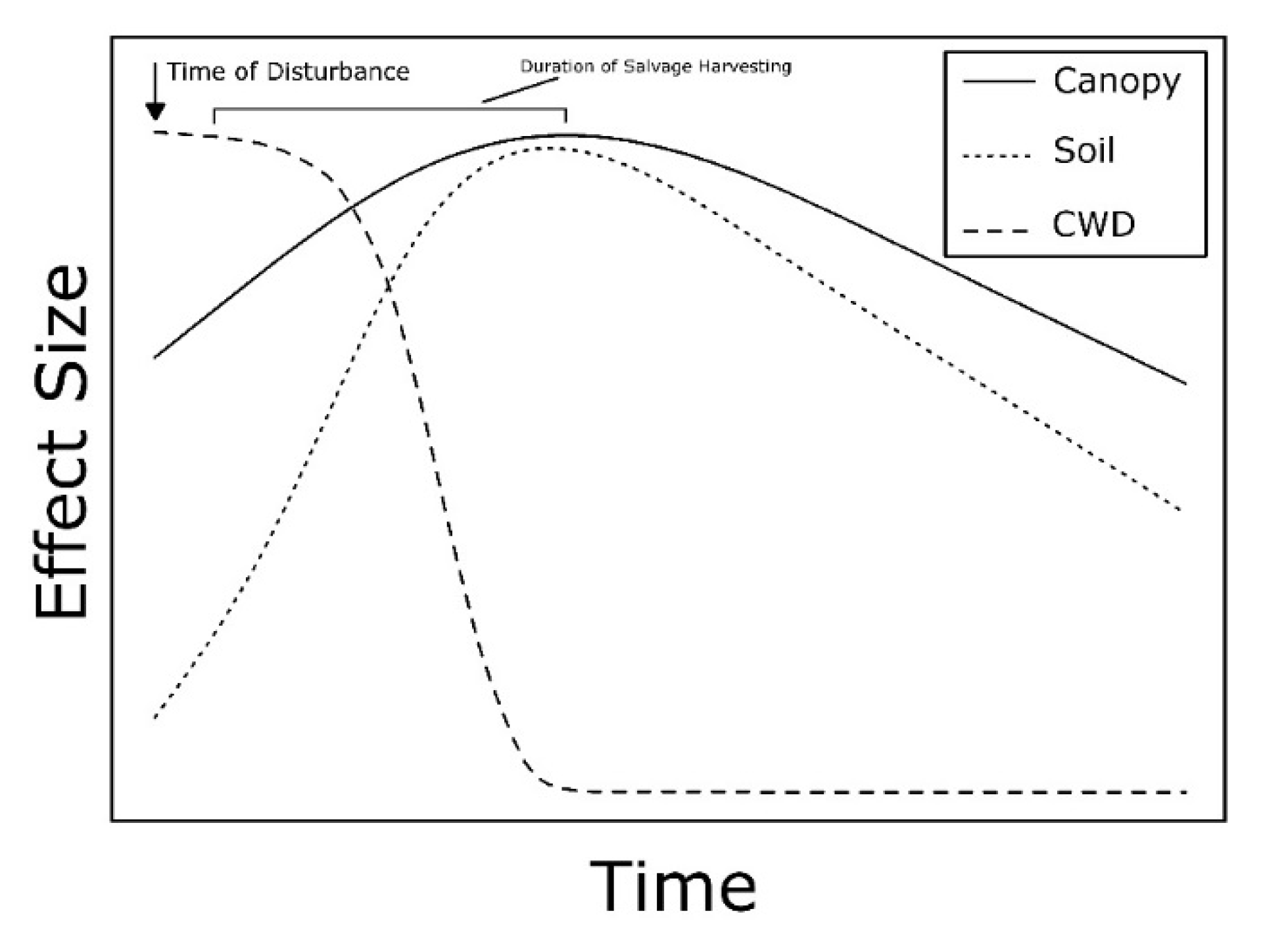
3.3. Salvage Logging
3.4. Responses of Invertebrate Taxa
4. Assessment of Conceptual Models and Conclusions
Supplementary Materials
Funding
Acknowledgements
Conflicts of Interest
References
- Frelich, L.E. Forest Dynamics and Disturbance Regimes: Studies from Temperate Evergreen-Deciduous Forests; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; John Wiley and Sons: New York, NY, USA, 1996. [Google Scholar]
- White, P.S.; Pickett, S.T.A. Natural disturbance and patch dynamics: An introduction. In The Ecology of Natural Disturbance and Patch Dynamics; Pickett, S.T.A., White, P.S., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1985; pp. 3–13. [Google Scholar]
- Runkle, J.R. Disturbance regimes in temperate forests. In The Ecology of Natural Disturbance and Patch Dynamics; Pickett, S.T.A., White, P.S., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1985; pp. 17–33. [Google Scholar]
- Pickett, S.T.A.; Kolasa, J.; Armesto, J.J.; Collins, S.L. The ecological concept of disturbance and its expression at various hierarchical levels. Oikos 1989, 54, 129–136. [Google Scholar] [CrossRef]
- Schowalter, T.D. Adaptations of insects to disturbance. In The Ecology of Natural Disturbance and Patch Dynamics; Pickett, S.T.A., White, P.S., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1985; pp. 235–252. [Google Scholar]
- Chapin, F.S., III; Matson, P.A.; Mooney, H.A. Principles of Terrestrial Ecosystem Ecology; Springer: New York, NY, USA, 2002. [Google Scholar]
- Schowalter, T.D. Insect responses to major landscape-level disturbance. Annu. Rev. Entomol. 2012, 57, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Franklin, J.F.; MacMahon, J.A. Messages from a mountain. Science 2000, 288, 1183–1184. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Noss, R.F. Salvage logging, ecosystem processes, and biodiversity conservation. Conserv. Biol. 2006, 20, 949–958. [Google Scholar] [CrossRef] [PubMed]
- Lindenmayer, D.B.; Burton, P.J.; Franklin, J.F. Salvage Logging and Its Ecological Consequences; Island Press: Washington, DC, USA, 2008. [Google Scholar]
- Franklin, J.F.; Spies, T.A.; Pelt, R.V.; Carey, A.B.; Thornburgh, D.A.; Berg, D.R.; Lindenmayer, D.B.; Harmon, M.E.; Keeton, W.S.; Shaw, D.C.; et al. Disturbances and structural development of natural forest ecosystems with silvicultural implications, using douglas-fir forests as an example. For. Ecol. Manag. 2002, 155, 399–423. [Google Scholar] [CrossRef]
- Foster, D.R.; Orwig, D.A. Preemptive and salvage harvesting of new England forests: When doing nothing is a viable alternative. Conserv. Biol. 2006, 20, 959–970. [Google Scholar] [CrossRef] [PubMed]
- Buddle, C.M.; Langor, D.W.; Pohl, G.R.; Spence, J.R. Arthropod responses to harvesting and wildfire: Implications for emulation of natural disturbance in forest management. Biol. Conserv. 2006, 128, 346–357. [Google Scholar] [CrossRef]
- Spies, T.A.; Turner, M.G. Dynamic forest mosaics. In Maintaining Biodiveristy in Forest Ecosystems; Hunter, M.L., Jr., Ed.; Cambridge University Press: Cambridge, UK, 1999; pp. 95–160. [Google Scholar]
- Petraitis, P.S.; Latham, R.E.; Niesenbaum, R.A. The maintenance of species diversity by disturbance. Q. Rev. Biol. 1989, 64, 393–418. [Google Scholar] [CrossRef]
- Boyd, I.L.; Freer-Smith, P.H.; Gilligan, C.A.; Godfray, H.C.J. The consequence of tree pests and diseases for ecosystem services. Science 2013, 342, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.J.; Spies, T.A.; Swanson, F.J.; Ohmann, J.L. Conserving biodiversity in managed forests. BioScience 1991, 41, 382–392. [Google Scholar] [CrossRef]
- Angelstam, P.K. Maintaining and restoring biodiversity in European boreal forests by developing natural disturbance regimes. J. Veg. Sci. 1998, 9, 593–602. [Google Scholar] [CrossRef]
- Bengtsson, J.; Nilsson, S.G.; Franc, A.; Menozzi, P. Biodiversity, disturbances, ecosystem function and management of european forests. For. Ecol. Manag. 2000, 132, 39–50. [Google Scholar] [CrossRef]
- Brussaard, L. Biodiversity and ecosystem functioning in soil. Ambio 1997, 26, 563–570. [Google Scholar]
- Lavelle, P.; Decaëns, T.; Aubert, M.; Barot, S.; Blouin, M.; Bureau, F.; Margerie, P.; Mora, P.; Rossi, J.P. Soil invertebrates and ecosystem services. Eur. J. Soil Biol. 2006, 42, S3–S15. [Google Scholar] [CrossRef]
- Wall, D.H.; Bardgett, R.D.; Behan-Pelletier, V.; Herrick, J.E.; Jones, T.H.; Ritz, K.; Six, J.; Strong, D.R.; van der Putten, W.H. Soil Ecology and Ecosystem Services; Oxford: Oxford, UK, 2012. [Google Scholar]
- Bergeron, Y.; Harvey, B.; Leduc, A.; Gauthier, S. Forest management guidelines based on natural disturbance dynamics: Stand- and forest-level considerations. For. Chron. 1999, 75, 49–54. [Google Scholar] [CrossRef]
- Paine, T.R.; Tegner, J.M.; Johnson, A.E. Compounded perturbations yield ecological surprises. Ecosystems 1998, 1, 535–545. [Google Scholar] [CrossRef]
- Perry, K.I.; Wallin, K.F.; Wenzel, J.W.; Herms, D.A. Characterizing movement of ground-dwelling arthropods with a novel mark-capture method using fluorescent powder. J. Insect Behav. 2017, 30, 32–47. [Google Scholar] [CrossRef]
- Brouwers, N.C.; Newton, A.C. Movement rates of woodland invertebrates: A systematic review of empirical evidence. Insect Conserv. Divers. 2009, 2, 10–22. [Google Scholar] [CrossRef]
- Runkle, J.R. Patterns of disturbance in some old-growth mesic forests of eastern north America. Ecology 1982, 63, 1533–1546. [Google Scholar] [CrossRef]
- Frelich, L.E.; Lorimer, C.G. Natural disturbance regimes in hemlock-hardwood forests of the upper great lakes region. Ecol. Monogr. 1991, 61, 145–164. [Google Scholar] [CrossRef]
- van Lierop, P.; Lindquist, E.; Sathyapala, S.; Franceschini, G. Global forest area disturbance from fire, insect pests, diseases and severe weather events. For. Ecol. Manag. 2015, 352, 78–88. [Google Scholar] [CrossRef]
- Vitousek, P.M.; D’Antonio, C.M.; Loope, L.L.; Westbrooks, R. Biological invasions as global environmental change. Am. Sci. 1996, 84, 468–478. [Google Scholar]
- Liebhold, A.M.; MacDonald, W.L.; Bergdahl, D.; Mastro, V.C. Invasion by exotic forest pests: A threat to forest ecosystems. For. Sci. 1995, 41, 1–49. [Google Scholar] [CrossRef]
- Wilcove, D.S.; Rothstein, D.; Jason, D.; Phillips, A.; Losos, E. Quantifying threats to imperiled species in the United States. BioScience 1998, 48, 607–615. [Google Scholar] [CrossRef]
- Mack, M.C.; D’Antonio, C.M. Impacts of biological invasions on disturbance regimes. Trends Ecol. Evol. 1998, 13, 195–198. [Google Scholar] [CrossRef]
- Gandhi, K.J.K.; Herms, D.A. Direct and indirect effects of alien insect herbivores on ecological processes and interactions in forests of eastern North America. Biol. Invasions 2010, 12, 389–405. [Google Scholar] [CrossRef]
- Peterson, C.J. Catastrophic wind damage to North American forests and the potential impact of climate change. Sci. Total Environ. 2000, 262, 287–311. [Google Scholar] [CrossRef]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P. The interplay between climate change, forests, and disturbances. Sci. Total Environ. 2000, 262, 201–204. [Google Scholar] [CrossRef]
- Dale, V.H.; Joyce, L.A.; McNulty, S.; Neilson, R.P.; Ayres, M.P.; Flannigan, M.D.; Hanson, P.J.; Irland, L.C.; Lugo, A.E.; Peterson, C.J.; et al. Climate change and forest disturbances. BioScience 2001, 51, 723–734. [Google Scholar] [CrossRef]
- Greenberg, C.H.; Forrest, T.G. Seasonal abundance of ground-occurring macroarthropods in forest and canopy gaps in the southern Appalachians. Southeast. Nat. 2003, 2, 591–608. [Google Scholar] [CrossRef]
- Perry, K.I.; Herms, D.A. Short-term responses of ground beetles to forest changes caused by early stages of emerald ash borer (coleoptera: Buprestidae)-induced ash mortality. Environ. Entomol. 2016, 45, 616–626. [Google Scholar] [CrossRef] [PubMed]
- Richardson, B.A.; Richardson, M.J.; González, G.; Shiels, A.B.; Srivastava, D.S. A canopy trimming experiment in Puerto Rico: The response of litter invertebrate communities to canopy loss and debris deposition in a tropical forest subject to hurricanes. Ecosystems 2010, 13, 286–301. [Google Scholar] [CrossRef]
- Gray, A.N.; Spies, T.A.; Easter, M.J. Microclimatic and soil moisture responses to gap formation in coastal douglas-fir forests. Can. J. For. Res. 2002, 32, 332–343. [Google Scholar] [CrossRef]
- Collins, B.S.; Pickett, S.T.A. Influence of canopy opening on the environment and herb layer in a northern hardwoods forest. Vegetatio 1987, 70, 3–10. [Google Scholar]
- Phillips, D.L.; Shure, D.J. Patch-size effects on early succession in southern Appalachian forests. Ecology 1990, 71, 204–212. [Google Scholar] [CrossRef]
- Fahey, R.T.; Puettmann, K.J. Ground-layer disturbance and initial conditions influence gap partitioning of understorey vegetation. J. Ecol. 2007, 95, 1098–1109. [Google Scholar] [CrossRef]
- Ishizuka, M.; Ochiai, Y.; Utsugi, H. Microenvironments and growth in gaps. In Diversity and Interaction in a Temperate Forest Community: Ogawa Forest Reserve of Japan; Nakashizuka, T., Matsumoto, Y., Eds.; Springer: Tokyo, Japan, 2002; pp. 229–244. [Google Scholar]
- Royo, A.A.; Carson, W.P. On the formation of dense understory layers in forests worldwide: Consequences and implications for forest dynamics, biodiversity, and succession. Can. J. For. Res. 2006, 36, 1345–1362. [Google Scholar] [CrossRef]
- Chen, J.; Saunders, S.C.; Crow, T.R.; Naiman, R.J.; Brosofske, K.D.; Mroz, G.D.; Brookshire, B.L.; Franklin, J.F. Microclimate in forest ecosystem and landscape ecology: Variations in local climate can be used to monitor and compare the effects of different management regimes. BioScience 1999, 49, 288–297. [Google Scholar] [CrossRef]
- Canham, C.D.; Denslow, J.S.; Platt, W.J.; Runkle, J.R.; Spies, T.A.; White, P.S. Light regimes beneath closed canopies and tree-fall gaps in temperate and tropical forests. Can. J. For. Res. 1990, 20, 620–631. [Google Scholar] [CrossRef]
- Perry, K.I.; Herms, D.A. Response of the forest floor invertebrate community to canopy gap formation caused by early stages of emerald ash borer-induced ash mortality. For. Ecol. Manag. 2016, 375, 259–267. [Google Scholar] [CrossRef]
- Perry, K.I.; Wallin, K.F.; Wenzel, J.W.; Herms, D.A. Forest disturbance and arthropods: Small-scale canopy gaps drive invertebrate community structure and composition. Ecosphere 2018, 9, e02463. [Google Scholar] [CrossRef]
- Shure, D.J.; Phillips, D.L. Patch size of forest openings and arthropod populations. Oecologia 1991, 86, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, K.J.K.; Gilmore, D.W.; Katovich, S.A.; Mattson, W.J.; Zasada, J.C.; Seybold, S.J. Catastrophic windstorm and fuel-reduction treatments alter ground beetle (Coleoptera: Carabidae) assemblages in a north American sub-boreal forest. For. Ecol. Manag. 2008, 256, 1104–1123. [Google Scholar] [CrossRef]
- Bouget, C.; Duelli, P. The effects of windthrow on forest insect communities: A literature review. Biol. Conserv. 2004, 118, 281–299. [Google Scholar] [CrossRef]
- Getz, L.L. Notes on the ecology of slugs: Arion circumscriptus, Deroceras reticulatum, and D. laeve. Am. Midl. Nat. 1959, 61, 485–498. [Google Scholar] [CrossRef]
- Chikoski, J.M.; Ferguson, S.H.; Meyer, L. Effects of water addition on soil arthropods and soil characteristics in a precipitation-limited environment. Acta Oecologica 2006, 30, 203–211. [Google Scholar] [CrossRef]
- Entling, W.; Schmidt, M.H.; Bacher, S.; Brandl, R.; Nentwig, W. Niche properties of central European spiders: Shading, moisture and the evolution of the habitat niche. Glob. Ecol. Biogeogr. 2007, 16, 440–448. [Google Scholar] [CrossRef]
- Levings, S.C.; Windsor, D.M. Litter moisture content as a determinant of litter arthropod distribution and abundance during the dry season on Barro Colorado Island, Panama. Biotropica 1984, 16, 125–131. [Google Scholar] [CrossRef]
- Verhoef, H.A. Soil moisture and the population dynamics of coexisting Collembola. Ecol. Bull. 1977, 25, 480–482. [Google Scholar]
- Verhoef, H.A.; van Selm, A.J. Distribution and population dynamics of Collembola in relation to soil moisture. Ecography 1983, 6, 387–388. [Google Scholar] [CrossRef]
- Choi, W.; Moorhead, D.; Neher, D.; Ryoo, M. A modeling study of soil temperature and moisture effects on population dynamics of Paronychiurus kimi (Collembola: Onychiuridae). Biol. Fertil. Soils 2006, 43, 69–75. [Google Scholar] [CrossRef]
- Neve, G. Influence of temperature and humidity on the activity of three Carabus species. In Carabid Beetles: Ecology and Evolution; Desender, K., Dufre’ne, M., Loreau, M., Luft, M.L., Maelfait, J.-P., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1994. [Google Scholar]
- Thiele, H.-U. Carabid Beetles in Their Environments: A Study on Habitat Selection by Adaptations in Physiology and Behavior; Springer: Berlin, Germany, 1977. [Google Scholar]
- Ruiter, P.C.D.; Griffiths, B.; Moore, J.C. Biodiversity and stability in soil ecosystems: Patterns, processes and the effects of disturbance. In Biodiversity and Ecosystem Functioning: Synthesis and Perspectives; Loreau, M., Naeem, S., Inchausti, P., Eds.; Oxford University Press: Oxford, UK, 2002; pp. 102–113. [Google Scholar]
- McElhinny, C.; Gibbons, P.; Brack, C.; Bauhus, J. Forest and woodland stand structural complexity: Its definition and measurement. For. Ecol. Manag. 2005, 218, 1–24. [Google Scholar] [CrossRef]
- Siitonen, J. Microhabitats. In Biodiversity in Dead Wood; Stokland, J.N., Siitonen, J., Jonsson, B.G., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 150–182. [Google Scholar]
- Harmon, M.E.; Franklin, J.F.; Swanson, F.J.; Sollins, P.; Gregory, S.V.; Lattin, J.D.; Anderson, N.H.; Cline, S.P.; Aumen, N.G.; Sedell, J.R.; et al. Ecology of coarse woody debris in temperate ecosystems. In Advances in Ecological Research; MacFadyen, A., Ford, E.D., Eds.; Academic Press: Cambridge, MA, USA, 1986; Volume 15, pp. 133–302. [Google Scholar]
- Tews, J.; Brose, U.; Grimm, V.; Tielborger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef]
- Stokland, J.N.; Siitonen, J. Mortality factors and decay succession. In Biodiversity in Dead Wood; Stokland, J.N., Siitonen, J., Jonsson, B.G., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 110–149. [Google Scholar]
- McComb, W.; Lindenmayer, D.B. Dying, dead, and downed trees. In Maintaining Biodiversity in Forest Ecosystems; Hunter, M.L., Jr., Ed.; Cambridge University Press: Cambridge, UK, 1999; pp. 335–372. [Google Scholar]
- Jonsson, B.G.; Stokland, J.N. The surrounding environment. In Biodiversity in Dead Wood; Stokland, J.N., Siitonen, J., Jonsson, B.G., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 194–217. [Google Scholar]
- Jabin, M.; Mohr, D.; Kappes, H.; Topp, W. Influence of deadwood on density of soil macro-arthropods in a managed oak-beech forest. For. Ecol. Manag. 2004, 194, 61–69. [Google Scholar] [CrossRef]
- Harmon, M.E.; Sexton, J. Guidelines for Measurements of Woody Detritus in Forest Ecosystems; Publication No. 20; U.S. LTER Network Office: Seattle, WA, USA, 1996; pp. 1–34. [Google Scholar]
- Moldenke, A.; Pajutee, M.; Ingham, E. The functional roles of forest soil arthropods: The soil is a lively place. In Proceedings of the California Forest Soils Council Conference on Forest Soils Biology and Forest Management; Gen Tech Rep PSW-GTR-178; USDA Forest Service, Pacific Southwest Research Station: Berkeley, CA, USA, 2000; pp. 7–22. [Google Scholar]
- Ulyshen, M.D.; Hanula, J.L. Litter-dwelling arthropod abundance peaks near coarse woody debris in loblolly pine forests of the southeastern United States. Fla. Entomol. 2009, 92, 163–164. [Google Scholar] [CrossRef]
- Koivula, M.; Punttila, P.; Haila, Y.; Niemelä, J. Leaf litter and the small-scale distribution of carabid beetles (Coleoptera, Carabidae) in the boreal forest. Ecography 1999, 22, 424–435. [Google Scholar] [CrossRef]
- Pearce, J.L.; Venier, L.A. The use of ground beetles (Coleoptera: Carabidae) and spiders (Araneae) as bioindicators of sustainable forest management: A review. Ecol. Indic. 2006, 6, 780–793. [Google Scholar] [CrossRef]
- Rainio, J.; Niemelä, J. Ground beetles (Coleoptera: Carabidae) as bioindicators. Biodivers. Conserv. 2003, 12, 487–506. [Google Scholar] [CrossRef]
- Greenslade, P. The potential of Collembola to act as indicators of landscape stress in Australia. Aust. J. Exp. Agric. 2007, 47, 424–434. [Google Scholar] [CrossRef]
- Folgarait, P.J. Ant biodiversity and its relationship to ecosystem functioning: A review. Biodivers. Conserv. 1998, 7, 1221–1244. [Google Scholar] [CrossRef]
- Dunn, J.P.; Potter, D.A.; Kimmerer, T.W. Carbohydrate reserves, radial growth, and mechanisms of resistance of oak trees to phloem-boring insects. Oecologia 1990, 83, 458–468. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.F. The relation between host condition and attacks by the bronzed birch borer. J. Econ. Entomol. 1944, 37, 588–596. [Google Scholar] [CrossRef]
- Blais, J.R. Trends in the frequency, extent, and severity of spruce budworm outbreaks in eastern Canada. Can. J. For. Res. 1983, 13, 539–547. [Google Scholar] [CrossRef]
- Royama, T.; MacKinnon, W.E.; Kettela, E.G.; Carter, N.E.; Hartling, L.K. Analysis of spruce budworm outbreak cycles in New Brunswick, Canada, since 1952. Ecology 2005, 86, 1212–1224. [Google Scholar] [CrossRef]
- Sippell, W. Outbreaks of the forest tent caterpillar, Malacosoma disstria Hbn., a periodic defoliator of broad-leaved trees in Ontario. Can. Entomol. 1962, 94, 408–416. [Google Scholar] [CrossRef]
- Cooke, B.J.; Lorenzetti, F. The dynamics of forest tent caterpillar outbreaks in Québec, Canada. For. Ecol. Manag. 2006, 226, 110–121. [Google Scholar] [CrossRef]
- Birch, M. Aggregation in bark beetles. In Chemical Ecology of Insects; Bell, W.J., Cardé, R.T., Eds.; Springer: Berlin/Heidelberg, Germany, 1984; pp. 331–353. [Google Scholar]
- Berryman, A.A.; Raffa, K.F.; Millstein, J.A.; Nils Chr, S. Interaction dynamics of bark beetle aggregation and conifer defense rates. Oikos 1989, 56, 256–263. [Google Scholar] [CrossRef]
- Vité, J.P.; Pitman, G.B. Bark beetle aggregation: Effects of feeding on the release of pheromones in Dendroctonus and Ips. Nature 1968, 218, 169–170. [Google Scholar] [CrossRef]
- Herms, D.A.; McCullough, D.G. Emerald ash borer invasion of North America: History, biology, ecology, impacts, and management. Annu. Rev. Entomol. 2014, 59, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Klooster, W.S.; Herms, D.A.; Knight, K.S.; Herms, C.P.; McCullough, D.G.; Smith, A.; Gandhi, K.J.K.; Cardina, J. Ash (Fraxinus spp.) mortality, regeneration, and seed bank dynamics in mixed hardwood forests following invasion by emerald ash borer (Agrilus planipennis). Biol. Invasions 2014, 16, 859–873. [Google Scholar] [CrossRef]
- Gandhi, K.J.K.; Smith, A.; Hartzler, D.M.; Herms, D.A. Indirect effects of emerald ash borer-induced ash mortality and canopy gap formation on epigaeic beetles. Environ. Entomol. 2014, 43, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, K.J.K.; Herms, D.A. North American arthropods at risk due to widespread Fraxinus mortality caused by the alien emerald ash borer. Biol. Invasions 2010, 12, 1839–1846. [Google Scholar] [CrossRef]
- Klooster, W.S.; Gandhi, K.J.K.; Long, L.C.; Perry, K.I.; Rice, K.B.; Herms, D.A. Ecological impacts of emerald ash borer in forests at the epicenter of the invasion in North America. Forests 2018, 9, 250. [Google Scholar] [CrossRef]
- Holmes, T.P.; Aukema, J.E.; Von Holle, B.; Liebhold, A.; Sills, E. Economic impacts of invasive species in forests. Ann. N. Y. Acad. Sci. 2009, 1162, 18–38. [Google Scholar] [CrossRef] [PubMed]
- Higham, M.; Hoven, B.M.; Gorchov, D.L.; Knight, K.S. Patterns of coarse woody debris in hardwood forests across a chronosequence of ash mortality due to the emerald ash borer (Agrilus planipennis). Nat. Areas J. 2017, 37, 406–411. [Google Scholar] [CrossRef]
- Perry, K.I.; Herms, D.A.; Klooster, W.S.; Smith, A.; Hartzler, D.M.; Coyle, D.R.; Gandhi, K.J.K. Downed coarse woody debris dynamics in ash (Fraxinus spp.) stands invaded by emerald ash borer (Agrilus planipennis Fairmaire). Forests 2018, 9, 191. [Google Scholar] [CrossRef]
- Perry, K.I.; Herms, D.A. Effects of late stages of emerald ash borer (Coleoptera: Buprestidae)-induced ash mortality on forest floor invertebrate communities. J. Insect Sci. 2017, 17, 119. [Google Scholar] [CrossRef]
- Perry, K.I.; Herms, D.A. Responses of ground-dwelling invertebrates to gap formation and accumulation of woody debris from invasive species, wind, and salvage logging. Forests 2017, 8, 174. [Google Scholar] [CrossRef]
- Herms, D.A.; McCullough, D.G. Emerald ash borer: Ecology and management. Encycl. Pest Manag. 2013, 1–14. [Google Scholar] [CrossRef]
- Ulyshen, M.D.; Klooster, W.S.; Barrington, W.T.; Herms, D.A. Impacts of emerald ash borer-induced tree mortality on leaf litter arthropods and exotic earthworms. Pedobiologia 2011, 54, 261–265. [Google Scholar] [CrossRef]
- Long, L.C. Direct and Indirect Impacts of Emerald Ash Borer on Forest Bird Communities. Master’s Thesis, The Ohio State University, Columbus, OH, USA, 2013. [Google Scholar]
- Kendrick, J.A.; Ribbons, R.R.; Classen, A.T.; Ellison, A.M. Changes in canopy structure and ant assemblages affect soil ecosystem variables as a foundation species declines. Ecosphere 2015, 6, 1–20. [Google Scholar] [CrossRef]
- Ellison, A.M.; Barker-Plotkin, A.A.; Foster, D.R.; Orwig, D.A. Experimentally testing the role of foundation species in forests: The Harvard Forest hemlock removal experiment. Methods Ecol. Evol. 2010, 1, 168–179. [Google Scholar] [CrossRef]
- Sackett, T.E.; Record, S.; Bewick, S.; Baiser, B.; Sanders, N.J.; Ellison, A.M. Response of macroarthropod assemblages to the loss of hemlock (Tsuga canadensis), a foundation species. Ecosphere 2011, 2, 1–16. [Google Scholar] [CrossRef]
- Record, S.; McCabe, T.; Baiser, B.; Ellison, A.M. Identifying foundation species in North American forests using long-term data on ant assemblage structure. Ecosphere 2018, 9, e02139. [Google Scholar] [CrossRef]
- Orwig, D.A.; Barker Plotkin, A.A.; Davidson, E.A.; Lux, H.; Savage, K.E.; Ellison, A.M. Foundation species loss affects vegetation structure more than ecosystem function in a northeastern USA forest. PeerJ 2013, 1, e41. [Google Scholar] [CrossRef] [PubMed]
- Orwig, D.A.; Foster, D.R. Forest response to the introduced hemlock woolly adelgid in southern New England, USA. J. Torrey Bot. Soc. 1998, 125, 60–73. [Google Scholar] [CrossRef]
- Ford, C.R.; Elliott, K.J.; Clinton, B.D.; Kloeppel, B.D.; Vose, J.M. Forest dynamics following eastern hemlock mortality in the southern Appalachians. Oikos 2012, 121, 523–536. [Google Scholar] [CrossRef]
- Orwig, D.A.; Foster, D.R.; Mausel, D.L. Landscape patterns of hemlock decline in New England due to the introduced hemlock woolly adelgid. J. Biogeogr. 2002, 29, 1475–1487. [Google Scholar] [CrossRef]
- McClure, M.S. Density-dependent feedback and population cycles in Adelges tsugae (Homoptera: Adelgidae) on Tsuga canadensis. Environ. Entomol. 1991, 20, 258–264. [Google Scholar] [CrossRef]
- Garneau, D.E.; Lawler, M.E.; Rumpf, A.S.; Weyburne, E.S.; Cuppernull, T.M.; Boe, A.G. Potential effects of beech bark disease on small mammals and invertebrates in northeastern US forests. Northeast. Nat. 2012, 19, 391–410. [Google Scholar] [CrossRef]
- Gandhi, K.J.K.; Gilmore, D.W.; Katovich, S.A.; Mattson, W.J.; Spence, J.R.; Seybold, S.J. Physical effects of weather events on the abundance and diversity of insects in North American forests. Environ. Rev. 2007, 15, 113–152. [Google Scholar] [CrossRef]
- Everham, E.M.; Brokaw, N.V.L. Forest damage and recovery from catastrophic wind. Bot. Rev. 1996, 62, 113–185. [Google Scholar] [CrossRef]
- Yamamoto, S.-I. The gap theory in forest dynamics. Bot. Mag. 1992, 105, 375–383. [Google Scholar] [CrossRef]
- Nelson, J.L.; Groninger, J.W.; Battaglia, L.L.; Ruffner, C.M. Bottomland hardwood forest recovery following tornado disturbance and salvage logging. For. Ecol. Manag. 2008, 256, 388–395. [Google Scholar] [CrossRef]
- Liechty, H.O.; Jurgensen, M.F.; Mroz, G.D.; Gale, M.R. Pit and mound topography and its influence on storage of carbon, nitrogen, and organic matter within an old-growth forest. Can. J. For. Res. 1997, 27, 1992–1997. [Google Scholar] [CrossRef]
- Clinton, B.D.; Baker, C.R. Catastrophic windthrow in the southern Appalachians: Characteristics of pits and mounds and initial vegetation responses. For. Ecol. Manag. 2000, 126, 51–60. [Google Scholar] [CrossRef]
- Schaetzl, R.J.; Burns, S.F.; Johnson, D.L.; Small, T.W. Tree uprooting: Review of impacts on forest ecology. Vegetatio 1989, 79, 165–176. [Google Scholar] [CrossRef]
- Sobhani, V.M.; Barrett, M.; Peterson, C.J. Robust prediction of treefall pit and mound sizes from tree size across 10 forest blowdowns in eastern North America. Ecosystems 2014, 17, 837–850. [Google Scholar] [CrossRef]
- Beatty, S.W.; Stone, E.L. The variety of soil microsites created by tree falls. Can. J. For. Res. 1986, 16, 539–548. [Google Scholar] [CrossRef]
- Cooper-Ellis, S.; Foster, D.R.; Carlton, G.; Lezberg, A. Forest response to catastrophic wind: Results from an experimental hurricane. Ecology 1999, 80, 2683–2696. [Google Scholar] [CrossRef]
- Meigs, G.W.; Keeton, W.S. Intermediate-severity wind disturbance in mature temperate forests: Legacy structure, carbon storage, and stand dynamics. Ecol. Appl. 2018, 28, 798–815. [Google Scholar] [CrossRef] [PubMed]
- Quine, C.; Coutts, M.; Gardiner, B.; Pyatt, G. Forests and Wind: Management to Minimise Damage; Forestry Commission, Bulletin 114; HMSO: London, UK, 1995. [Google Scholar]
- Foster, D.R.; Aber, J.D.; Melillo, J.M.; Bowden, R.D.; Bazzaz, F.A. Forest response to disturbance and anthropogenic stress: Rethinking the 1938 hurricane and the impact of physical disturbance vs. chemical and climate stress on forest ecosystems. BioScience 1997, 47, 437–445. [Google Scholar] [CrossRef]
- Young, T.P.; Hubbell, S.P. Crown asymmetry, treefalls, and repeat disturbance of broad-leaved forest gaps. Ecology 1991, 72, 1464–1471. [Google Scholar] [CrossRef]
- Perry, K.I. Responses of Ground-Dwelling Invertebrate Communities to Disturbance in Forest Ecosystems. Ph.D. Thesis, Ohio State University, Columbus, OH, USA, 2016. [Google Scholar]
- Patrick, M.; Fowler, D.; Dunn, R.R.; Sanders, N.J. Effects of treefall gap disturbances on ant assemblages in a tropical montane cloud forest. Biotropica 2012, 44, 472–478. [Google Scholar] [CrossRef]
- Basu, P. Seasonal and spatial patterns in ground foraging ants in a rain forest in the western Ghats, India. Biotropica 1997, 29, 489–500. [Google Scholar] [CrossRef]
- Feener, H.D., Jr.; Schupp, W.E. Effect of treefall gaps on the patchiness and species richness of neotropical ant assemblages. Oecologia 1998, 116, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Sklodowski, J.; Garbalinska, P. Ground beetle (Coleoptera, Carabidae) assemblages inhabiting scots pine stands of Puszcza Piska forest: Six-year responses to a tornado impact. ZooKeys 2011, 100, 371–392. [Google Scholar] [CrossRef] [PubMed]
- Barber, N.A.; Widick, W.L. Localized effects of tornado damage on ground beetle communities and vegetation in a forested preserve. Nat. Areas J. 2017, 37, 489–496. [Google Scholar] [CrossRef]
- Garbalińska, P.; Skłodowski, J. Body size differentiation in selected carabid species inhabiting Puszcza Piska forest stands disturbed by the hurricane. Balt. J. Coleopterol. 2008, 8, 101–114. [Google Scholar]
- Skłodowski, J.; Garbalińska, P. Ground beetle assemblages (Coleoptera, Carabidae) in the third year of regeneration after a hurricane in the Puszcza Piska pine forests. Balt. J. Coleopterol. 2007, 7, 17–36. [Google Scholar]
- Peterson, C.J.; Leach, A.D. Limited salvage logging effects on forest regeneration after moderate-severity windthrow. Ecol. Appl. 2008, 18, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Franklin, J.F.; Mitchell, R.J.; Palik, B.J. Natural Disturbance and Stand Development Principles for Ecological Forestry; General Technical Report NRS-19; USDA Forest Service: Newtown Square, PA, USA, 2007. [Google Scholar]
- Fettig, C.J.; Klepzig, K.D.; Billings, R.F.; Munson, A.S.; Nebeker, T.E.; Negrón, J.F.; Nowak, J.T. The effectiveness of vegetation management practices for prevention and control of bark beetle infestations in coniferous forests of the western and southern united states. For. Ecol. Manag. 2007, 238, 24–53. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Foster, D.R.; Franklin, J.F.; Hunter, M.L.; Noss, R.F.; Schmiegelow, F.A.; Perry, D. Salvage harvesting policies after natural disturbance. Science 2004, 303, 1303. [Google Scholar] [CrossRef] [PubMed]
- Lousier, J.D. Impacts of Forest Harvesting and Regeneration on Forest Sites; Land Management Report 67; Research Branch Ministry of Forests: Victoria, BC, Canada, 1990; pp. 1–103. [Google Scholar]
- McNabb, D.H.; Startsev, A.D.; Nguyen, H. Soil wetness and traffic level effects on bulk density and air-filled porosity of compacted boreal forest soils. Soil Sci. Soc. Am. J. 2001, 65, 1238–1247. [Google Scholar] [CrossRef]
- Thom, D.; Seidl, R. Natural disturbance impacts on ecosystem services and biodiversity in temperate and boreal forests. Biol. Rev. 2016, 91, 760–781. [Google Scholar] [CrossRef] [PubMed]
- Leverkus, A.B.; Rey Benayas, J.M.; Castro, J.; Boucher, D.; Brewer, S.; Collins, B.M.; Donato, D.; Fraver, S.; Kishchuk, B.E.; Lee, E.-J.; et al. Salvage logging effects on regulating and supporting ecosystem services—A systematic map. Can. J. For. Res. 2018, 48, 983–1000. [Google Scholar] [CrossRef]
- Likens, G.E.; Bormann, F.H.; Pierce, R.S.; Reiners, W.A. Recovery of a deforested ecosystem. Science 1978, 199, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Bormann, F.H.; Likens, G.E. Catastrophic disturbance and the steady state in northern hardwood forests: A new look at the role of disturbance in the development of forest ecosystems suggests important implications for land-use policies. Am. Sci. 1979, 67, 660–669. [Google Scholar]
- Stokland, J.N. The coarse woody debris profile: An archive of recent forest history and an important biodiversity indicator. Ecol. Bull. 2001, 49, 71–83. [Google Scholar]
- Jonsson, B.G.; Siitonen, J. Dead wood and sustainable forest management. In Biodiversity in Dead Wood; Stokland, J.N., Siitonen, J., Jonsson, B.G., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 302–337. [Google Scholar]
- Work, T.T.; Brais, S.; Harvey, B.D. Reductions in downed deadwood from biomass harvesting alter composition of spiders and ground beetle assemblages in jack-pine forests of western Quebec. For. Ecol. Manag. 2014, 321, 19–28. [Google Scholar] [CrossRef]
- Baker, S.C.; Richardson, A.M.M.; Seeman, O.D.; Barmuta, L.A. Does clearfell, burn and sow silviculture mimic the effect of wildfire? A field study and review using litter beetles. For. Ecol. Manag. 2004, 199, 433–448. [Google Scholar] [CrossRef]
- Fail, J., Jr. Production and decomposition rates of a coastal plain forest following the impact of hurricane Hugo. J. Elisha Mitchell Sci. Soc. 1999, 115, 47–54. [Google Scholar]
- Worrell, R.; Hampson, A. The influence of some forest operations on the sustainable management of forest soils—A review. Forestry 1997, 70, 61–85. [Google Scholar] [CrossRef]
- Hayes, J.P.; Schoenholtz, S.H.; Hartley, M.J.; Murphy, G.; Powers, R.F.; Berg, D.; Radosevich, S.R. Environmental consequences of intensively managed forest plantations in the pacific northwest. J. For. 2005, 103, 83–87. [Google Scholar]
- McIver, J.D.; Starr, L. A literature review on the environmental effects of postfire logging. West. J. Appl. For. 2001, 16, 159–168. [Google Scholar]
- Wagenbrenner, J.W.; MacDonald, L.H.; Coats, R.N.; Robichaud, P.R.; Brown, R.E. Effects of post-fire salvage logging and a skid trail treatment on ground cover, soils, and sediment production in the interior western United States. For. Ecol. Manag. 2015, 335, 176–193. [Google Scholar] [CrossRef]
- Greacen, E.; Sands, R. Compaction of forest soils. A review. Soil Res. 1980, 18, 163–189. [Google Scholar] [CrossRef]
- Page-Dumroese, D.S.; Jurgensen, M.F.; Tiarks, A.E.; Ponder, J.F.; Sanchez, F.G.; Fleming, R.L.; Kranabetter, J.M.; Powers, R.F.; Stone, D.M.; Elioff, J.D.; et al. Soil physical property changes at the North American long-term soil productivity study sites: 1 and 5 years after compaction. Can. J. For. Res. 2006, 36, 551–564. [Google Scholar] [CrossRef]
- Sands, R.; Greacen, E.; Gerard, C. Compaction of sandy soils in radiata pine forests. Soil Res. 1979, 17, 101–113. [Google Scholar] [CrossRef]
- Wingate-Hill, R.; Jakobsen, B. Increased mechanisation and soil damage in forests—A review. N. Z. J. For. Sci. 1982, 12, 380–393. [Google Scholar]
- Urbanovičová, V.; Kováč, Ľ.; Miklisová, D. Epigeic arthropod communities of spruce forest stands in the High Tatra Mts. (Slovakia) with special reference to Collembola–first year after windthrow. Acta Soc. Zool. Bohem. 2010, 74, 21–29. [Google Scholar]
- Urbanovičová, V.; Miklisová, D.; Kováč, Ľ. Forest disturbance enhanced the activity of epedaphic Collembola in windthrown stands of the High Tatra Mountains. J. Mt. Sci. 2014, 11, 449–463. [Google Scholar] [CrossRef]
- Wermelinger, B.; Moretti, M.; Duelli, P.; Lachat, T.; Pezzatti, G.B.; Obrist, M.K. Impact of windthrow and salvage-logging on taxonomic and functional diversity of forest arthropods. For. Ecol. Manag. 2017, 391, 9–18. [Google Scholar] [CrossRef]
- Phillips, I.D.; Cobb, T.P.; Spence, J.R.; Brigham, R.M. Salvage logging, edge effects, and carabid beetles: Connections to conservation and sustainable forest management. Environ. Entomol. 2006, 35, 950–957. [Google Scholar] [CrossRef]
- Koivula, M.; Spence, J.R. Effects of post-fire salvage logging on boreal mixed-wood ground beetle assemblages (Coleoptera, Carabidae). For. Ecol. Manag. 2006, 236, 102–112. [Google Scholar] [CrossRef]
- Thorn, S.; Bußler, H.; Fritze, M.-A.; Goeder, P.; Müller, J.; Weiß, I.; Seibold, S. Canopy closure determines arthropod assemblages in microhabitats created by windstorms and salvage logging. For. Ecol. Manag. 2016, 381, 188–195. [Google Scholar] [CrossRef]
- Thorn, S.; Bässler, C.; Brandl, R.; Burton, P.J.; Cahall, R.; Campbell, J.L.; Castro, J.; Choi, C.-Y.; Cobb, T.; Donato, D.C.; et al. Impacts of salvage logging on biodiversity: A meta-analysis. J. Appl. Ecol. 2017, 55, 279–289. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perry, K.I.; Herms, D.A. Dynamic Responses of Ground-Dwelling Invertebrate Communities to Disturbance in Forest Ecosystems. Insects 2019, 10, 61. https://doi.org/10.3390/insects10030061
Perry KI, Herms DA. Dynamic Responses of Ground-Dwelling Invertebrate Communities to Disturbance in Forest Ecosystems. Insects. 2019; 10(3):61. https://doi.org/10.3390/insects10030061
Chicago/Turabian StylePerry, Kayla I., and Daniel A. Herms. 2019. "Dynamic Responses of Ground-Dwelling Invertebrate Communities to Disturbance in Forest Ecosystems" Insects 10, no. 3: 61. https://doi.org/10.3390/insects10030061
APA StylePerry, K. I., & Herms, D. A. (2019). Dynamic Responses of Ground-Dwelling Invertebrate Communities to Disturbance in Forest Ecosystems. Insects, 10(3), 61. https://doi.org/10.3390/insects10030061