Sickly Sweet: Insecticidal Polyols Induce Lethal Regurgitation in Dipteran Pests

 , , ,
, , ,  and
and 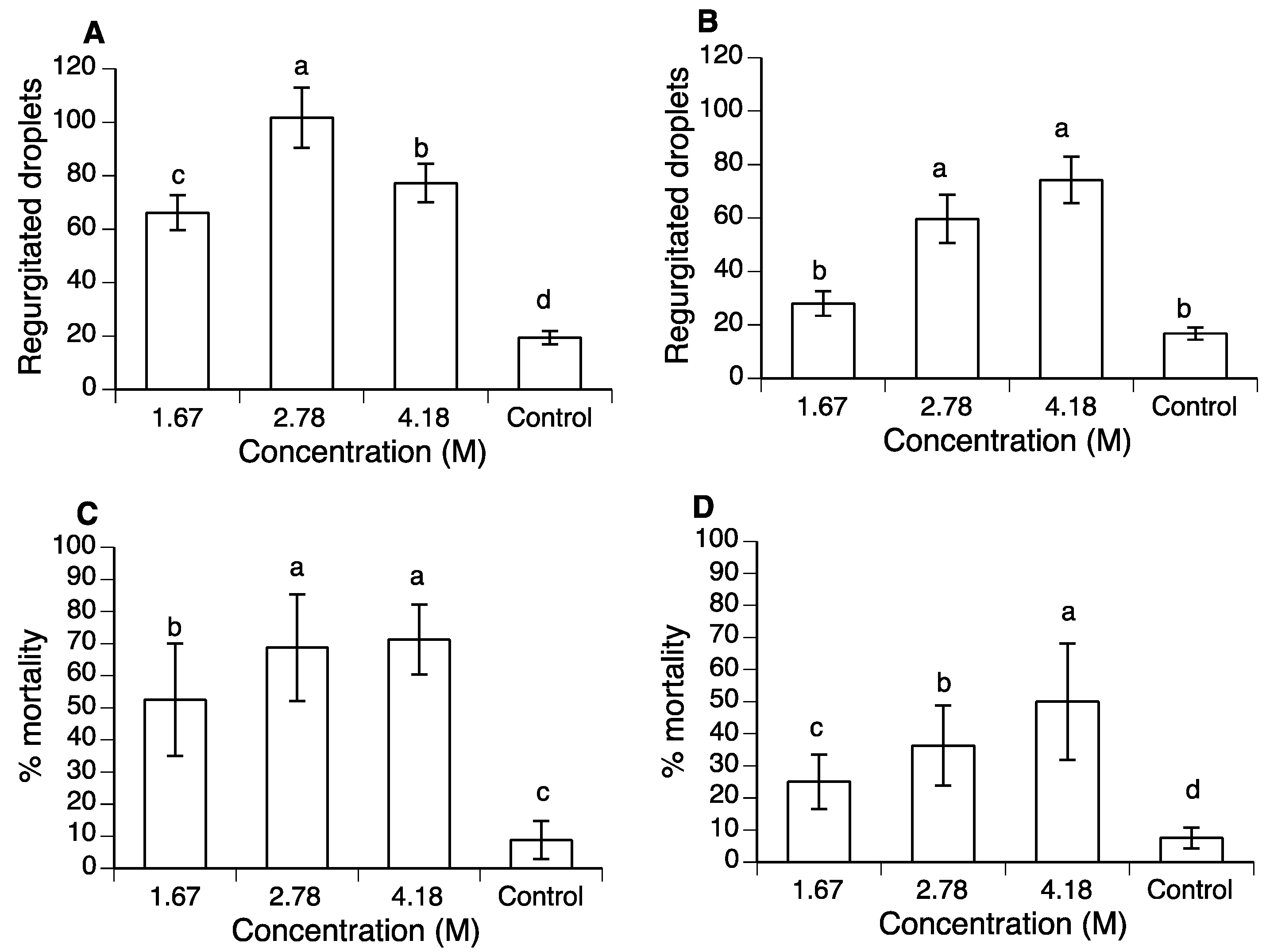{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects and Compounds
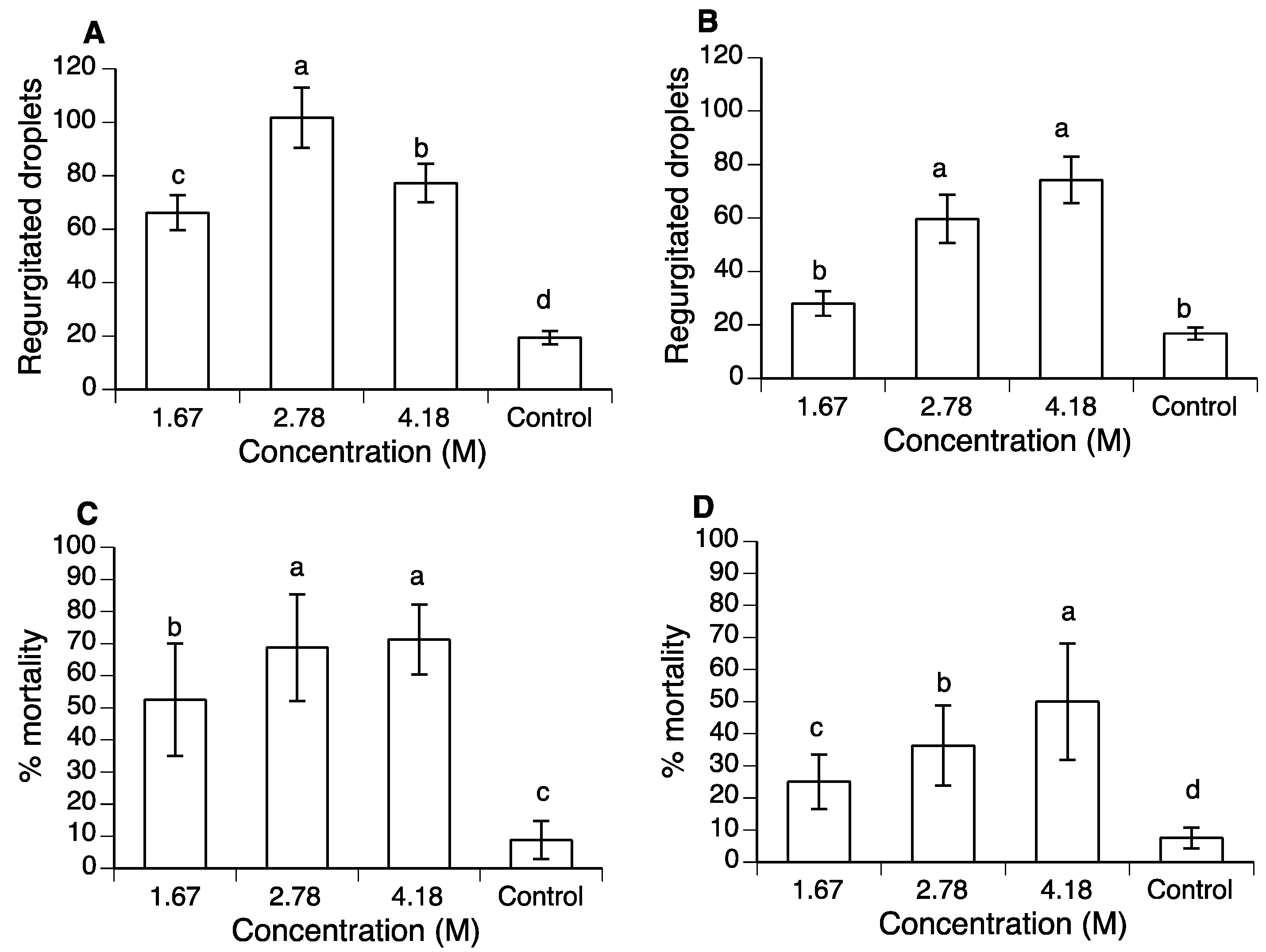
2.2. Concentration–Mortality Response and Regurgitation of Polyols in A. ludens
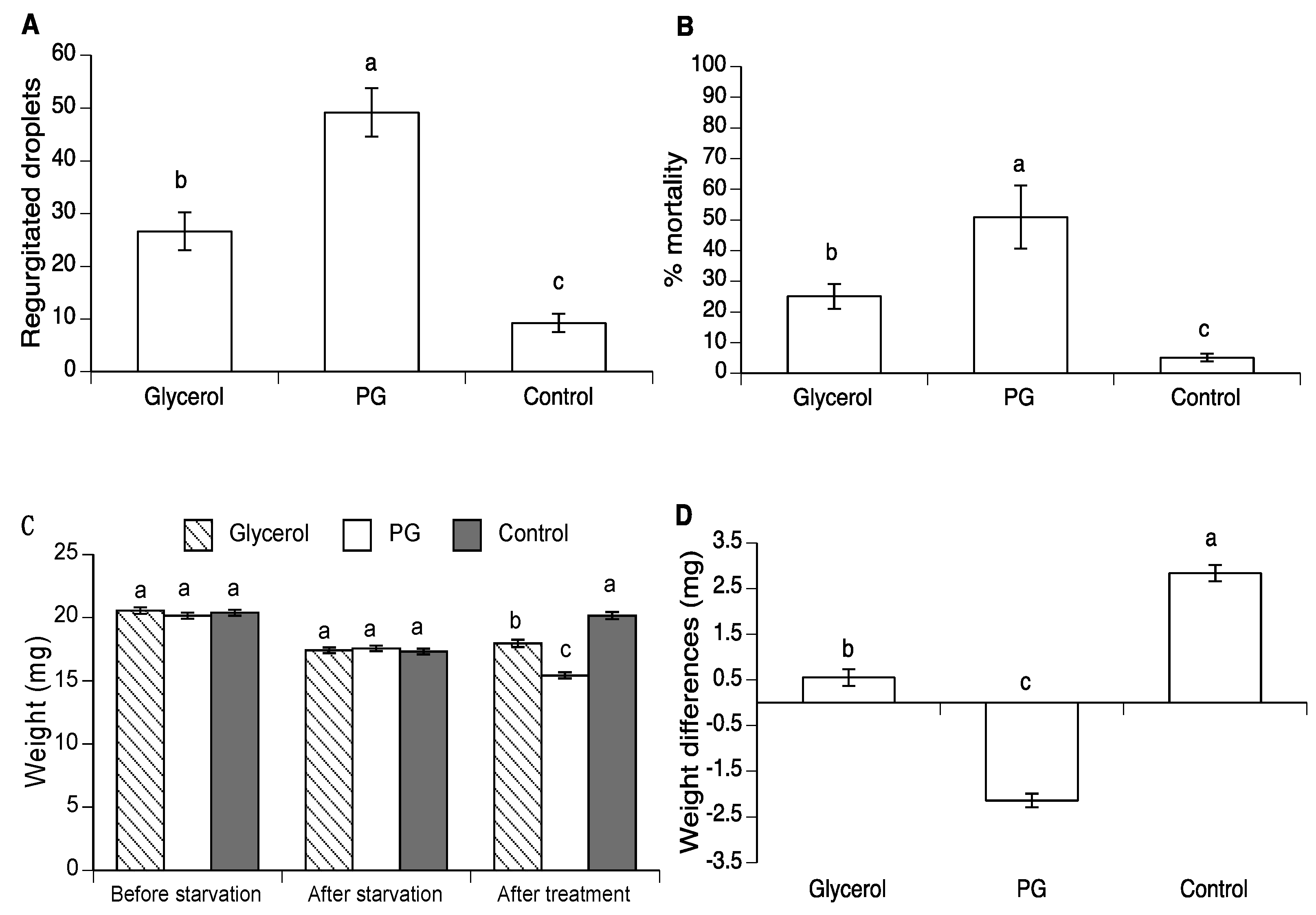
2.3. Effect of Glycerol and PG on Body Weight in A. ludens and A. obliqua
2.4. Effect of Glycerol and PG on Mortality and Regurgitation of D. suzukii
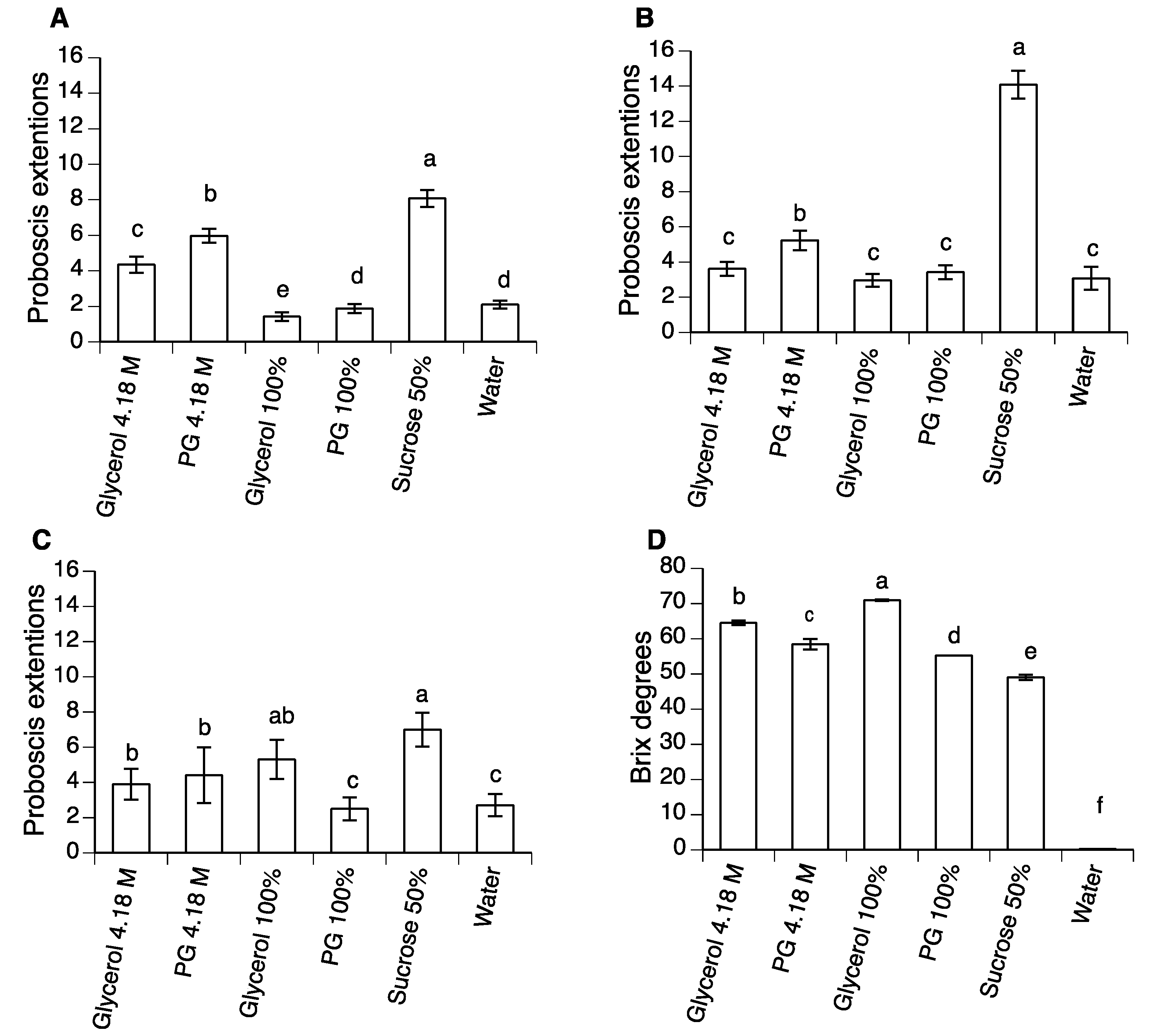
2.5. Proboscis Extension Reflex to PG, Glycerol and Sucrose
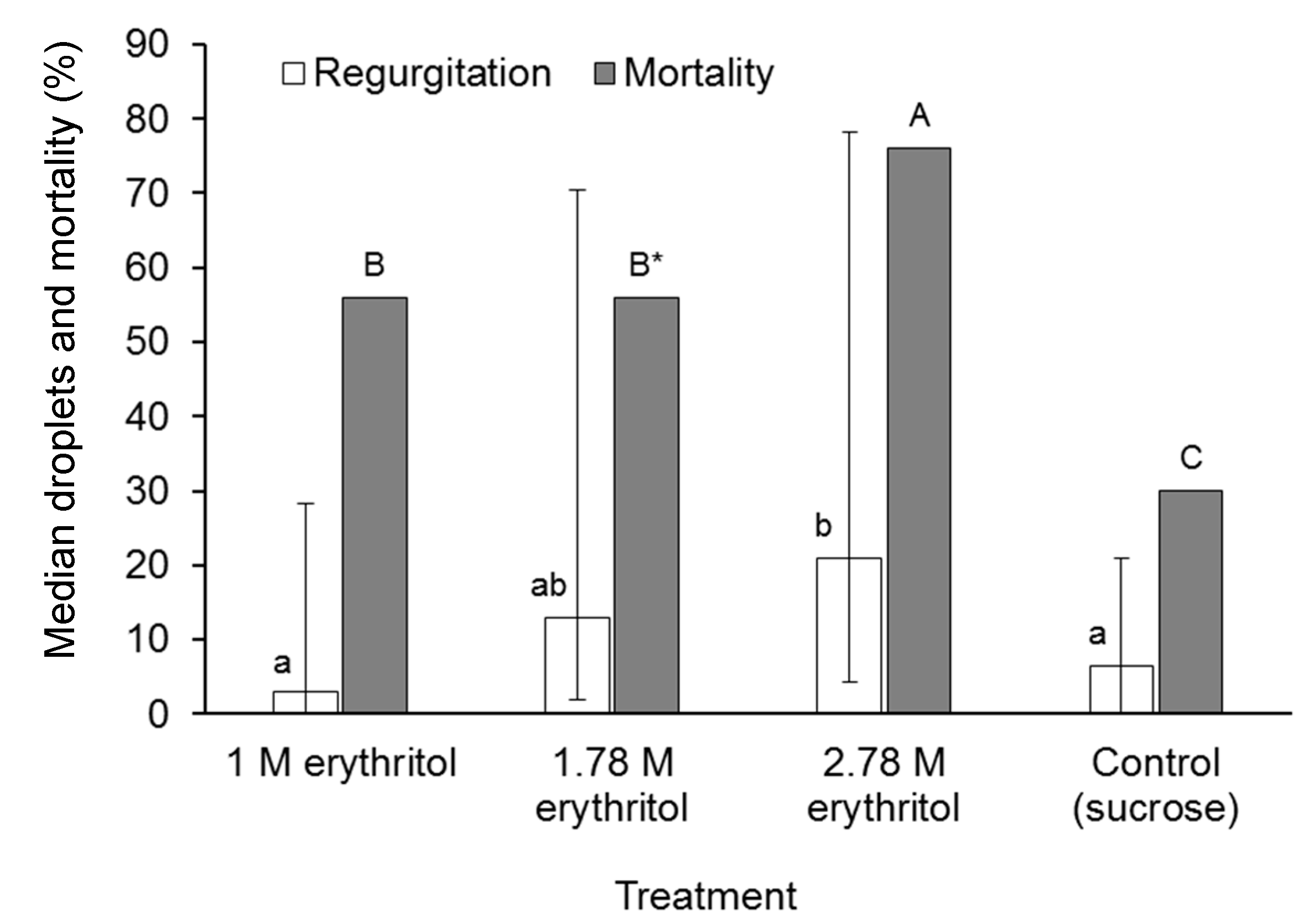
2.6. The Effect of Erythritol on Regurgitation and Mortality in A. ludens
2.7. Statistical Analyses
3. Results
3.1. Concentration–Mortality Response of Polyols in A. ludens
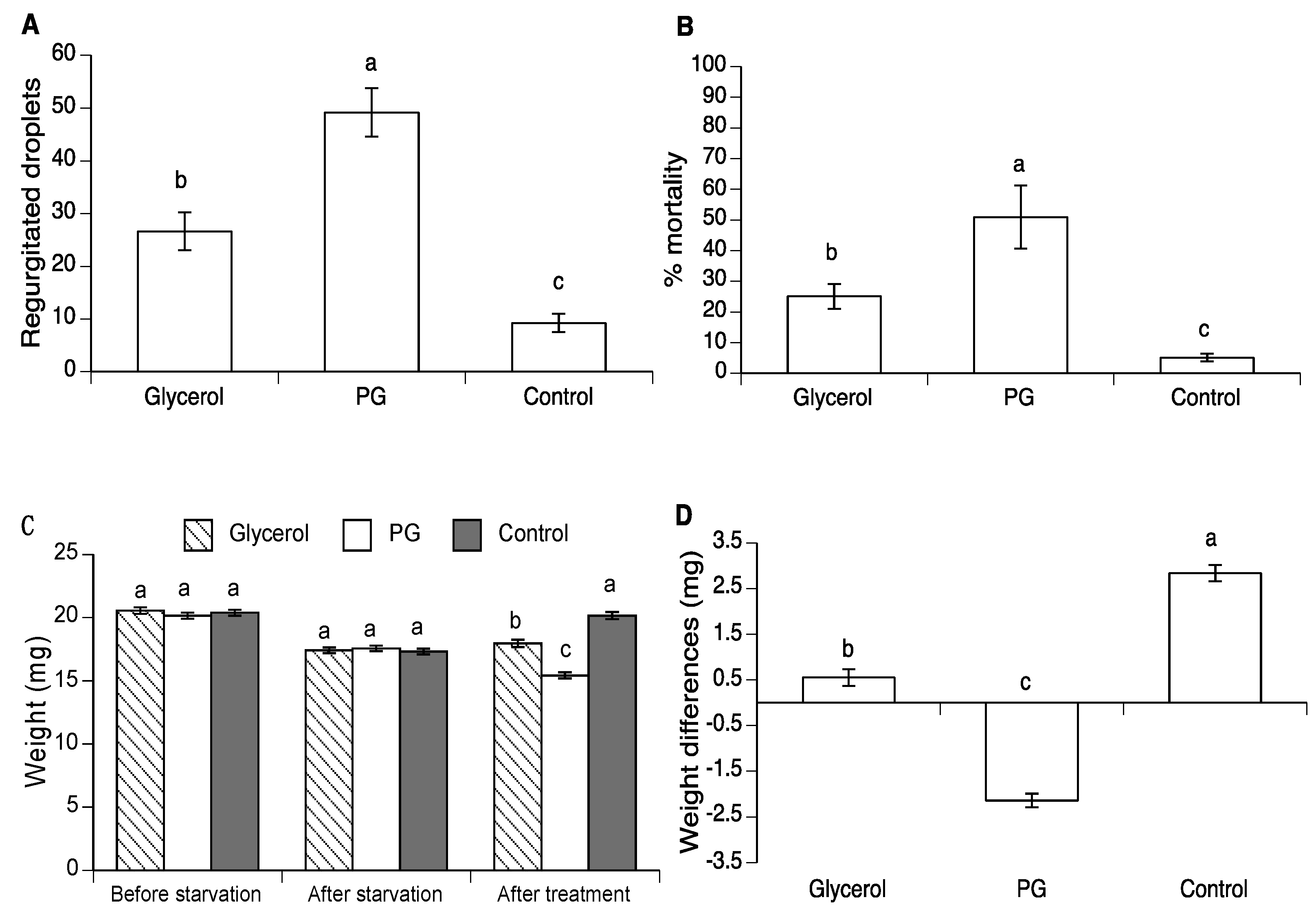
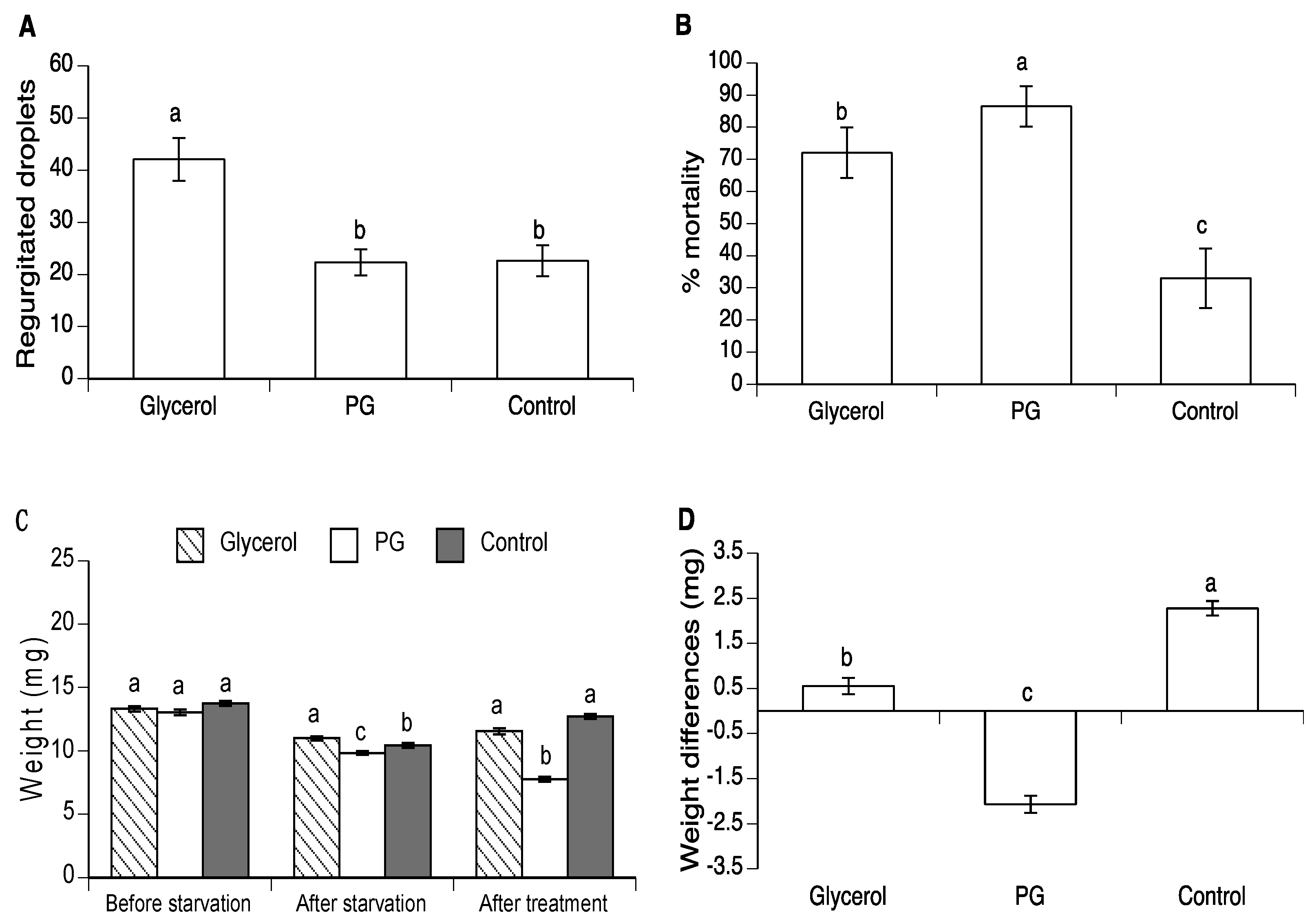
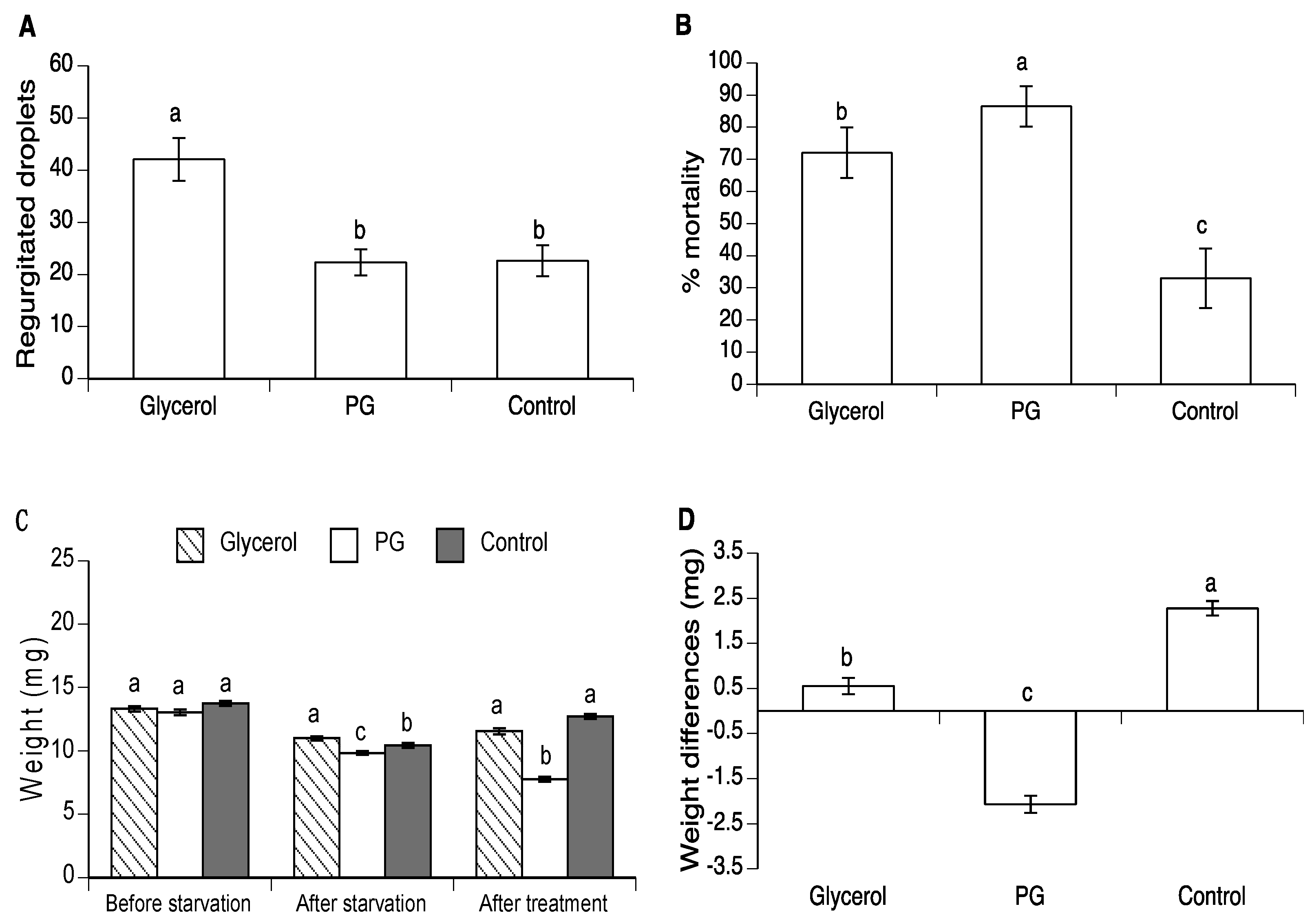
3.2. Effect of Glycerol and PG on Body Weight in A. ludens and A. obliqua
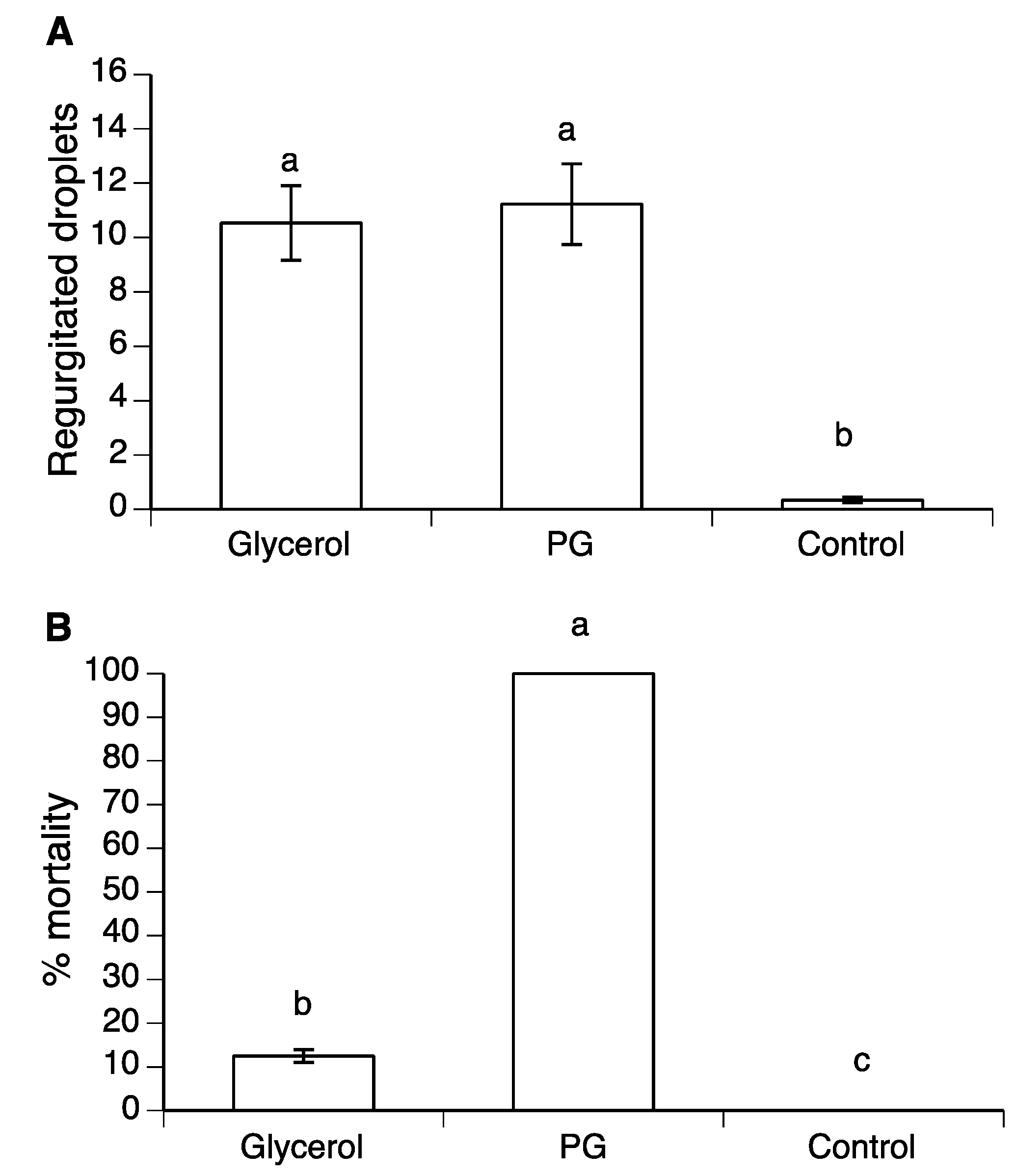
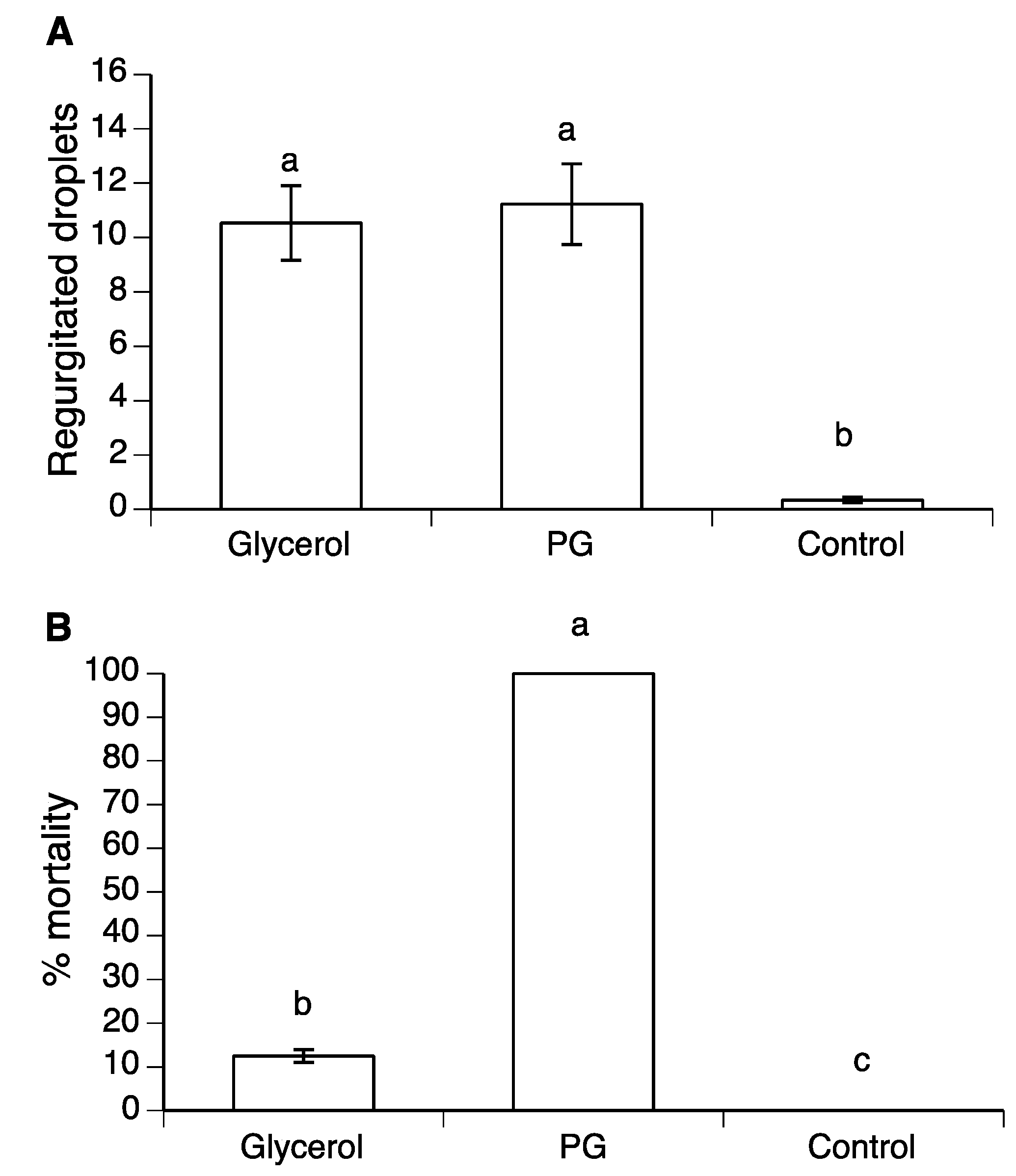
3.3. Effect of Glycerol and PG on Mortality and Regurgitation of Drosophila suzukii
3.4. Proboscis Extension Reflex Studies
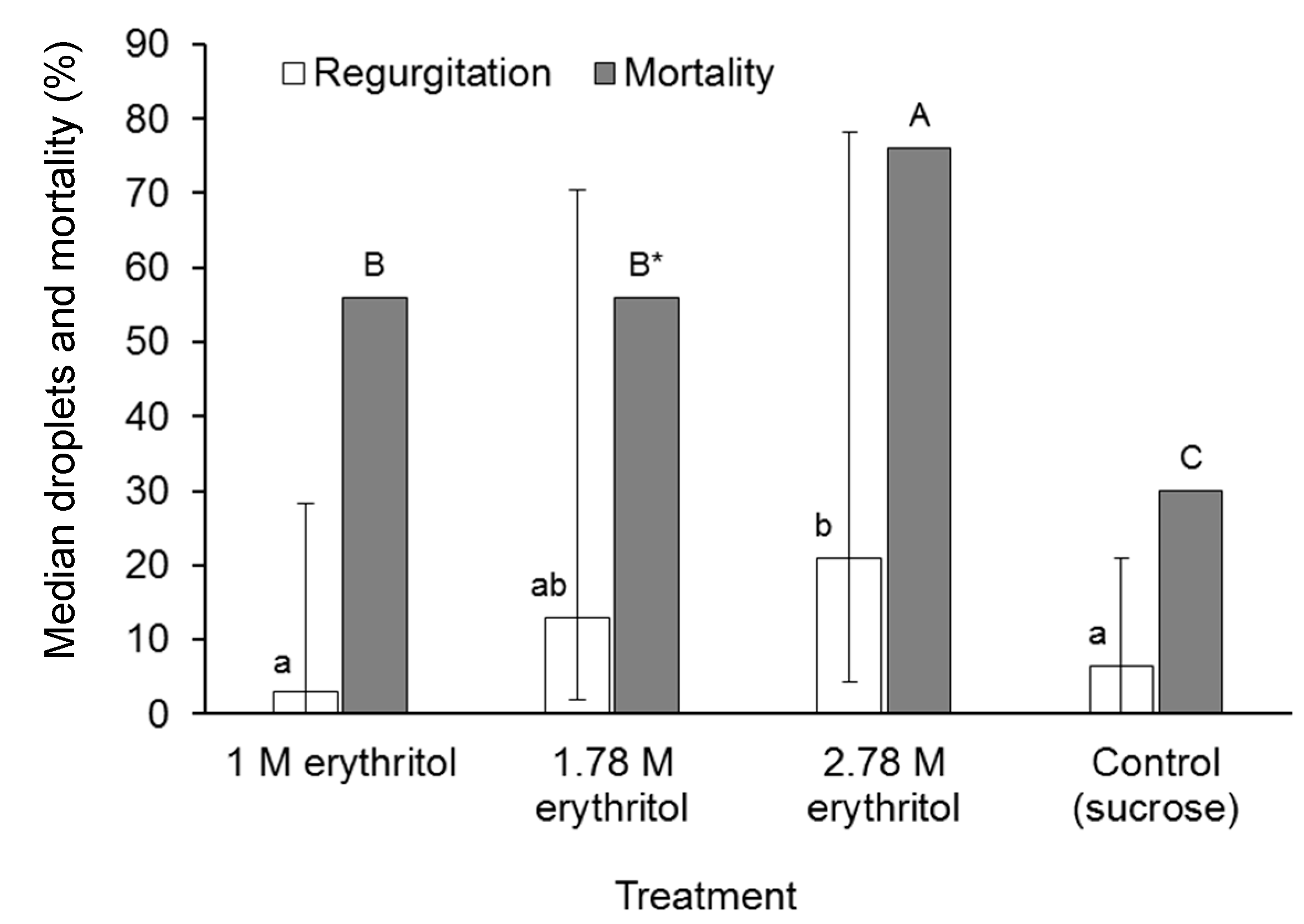
3.5. The Effect of Erythritol on Regurgitation and Mortality in A. ludens
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Ansari, M.; Moraiet, M.; Ahmad, S. Insecticides: Impact on the Environment and Human Health. In Environmental Deterioration and Human Health; Malik, A., Grohmann, E., Akhtar, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 99–123. [Google Scholar]
- Nauen, R.; Bretschneider, T. New modes of action of insecticides. Pestic Outlook 2002, 13, 241–245. [Google Scholar] [CrossRef]
- Doucet, D.; Retnakaran, A. Insect chitin: Metabolism, genomics and pest management. Adv. Insect Physiol. 2012, 43, 437–511. [Google Scholar]
- Federici, B.A. Insecticidal bacterial proteins identify the midgut epithelium as a source of novel target sites for insect control. Arch. Insect Biochem. Physiol. 1993, 22, 357–371. [Google Scholar] [CrossRef]
- Piermarini, P.M.; Esquivel, C.J.; Denton, J.S. Malpighian tubules as novel targets for mosquito control. Int. J. Environ. Res. Public Health. 2017, 14, 111. [Google Scholar] [CrossRef] [PubMed]
- Stoffolano, J.G., Jr.; Haselton, A.T. The adult dipteran crop: A unique and overlooked organ. Annu. Rev. Entomol. 2013, 58, 205–225. [Google Scholar] [CrossRef] [PubMed]
- Terra, W.R. Evolution of digestive systems of insects. Annu. Rev. Entomol. 1990, 35, 181–200. [Google Scholar] [CrossRef]
- Gomes, G.; Köberle, R.; Von Zuben, C.J.; Andrade, D.V. Droplet bubbling evaporatively cools a blowfly. Sci. Rep. 2018, 8, 5464. [Google Scholar] [CrossRef] [PubMed]
- Baudier, K.M.; Kaschock-Marenda, S.D.; Patel, N.; Diangelus, K.L.; O’Donnell, S.; Marenda, D.R. Erythritol, a non-nutritive sugar alcohol sweetener and the main component of Truvia®, is a palatable ingested insecticide. PLoS ONE 2014, 9, e98949. [Google Scholar] [CrossRef]
- Choi, M.-Y.; Tang, S.B.; Ahna, S.-J.; Amarasekare, K.G.; Shearere, P.N.D.; Lee, J.C. Effect of non-nutritive sugars to decrease the survivorship of spotted wing drosophila, Drosophila suzukii. J. Insect Physiol. 2017, 99, 86–94. [Google Scholar] [CrossRef]
- Fisher, M.L.; Fowler, F.E.; Denning, S.S.; Watson, D.W. Survival of the House Fly (Diptera: Muscidae) on truvia and other sweeteners. J. Med. Entomol. 2017, 54, 999–1005. [Google Scholar] [CrossRef]
- Zheng, C.Y.; Zeng, L.; Xu, Y.J. Effect of sweeteners on the survival and behaviour of Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Pest Manag. Sci. 2016, 72, 990–996. [Google Scholar] [CrossRef] [PubMed]
- Gilkey, P.L.; Bolshakov, D.T.; Kowala, J.G.; Taylor, L.A.; O’Donnell, S.; Marenda, D.R.; Sirot, L.K. Lethal effects of erythritol on the mosquito Aedes aegypti Linnaeus (Diptera: Culicidae). J. Appl. Entomol. 2018. [Google Scholar] [CrossRef]
- Boesten, D.M.; den Hartog, G.J.; de Cock, P.; Bosscher, D.; Bonnema, A.; Bast, A. Health effects of erythritol. Nutrafoods 2015, 14, 3–9. [Google Scholar] [CrossRef]
- Mortensen, A.; Aguilar, F.; Crebelli, R.; Di Domenico, A.; Dusemund, B.; Frutos, M.J.; Galtier, P.; Gott, D.; Gundert-Remy, U.; Leblanc, J.C. Re-evaluation of glycerol (E 422) as a food additive. EFSA J. 2017, 15, 4720. [Google Scholar]
- Thomas, D.B.; Robacker, D.C. Trapping for Mexican fruit fly (Diptera: Tephritidae) with torula yeast and propylene glycol. Subtrop. Plant Sci. 2006, 58, 23–25. [Google Scholar]
- Gordesky-Gold, B.; Rivers, N.; Ahmed, O.M.; Breslin, P.A. Drosophila melanogaster prefers compounds perceived sweet by humans. Chem. Senses. 2008, 33, 301–309. [Google Scholar] [CrossRef]
- Aluja, M.; Piñero, J.; Jácome, I.; Díaz-Fleischer, F.; Sivinski, J. Behavior of flies in the genus Anastrepha (Trypetinae: Toxotrypanini). In Fruit Flies (Tephritidae): Phylogeny and Evolution of Behavior; Aluja, M., Norrbom, A., Eds.; CRC Press: Boca Raton, FL, USA, 1999; pp. 375–406. [Google Scholar]
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; Isaacs, R. Invasion biology of spotted wing Drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Domínguez, J.; Artiaga-López, T.; Solís, E.; Hernández, E. Métodos de colonización y cría masiva. In Moscas de la Fruta: Fundamentos y Procedimientos para su Manejo; Montoya, P., Toledo, J., Hernández, E., Eds.; S y G. Editores: Mexico City, Mexico, 2010; pp. 259–276. [Google Scholar]
- Liedo, P.; Carey, J.R.; Celedonio, H.; Guillen, J. Size specific demography of three species of Anastrepha fruit flies. Entomol. Exp. Appl. 1992, 63, 135–142. [Google Scholar] [CrossRef]
- Dalton, D.T.; Walton, V.M.; Shearer, P.W.; Walsh, D.B.; Capriled, J.; Isaacs, R. Laboratory survival of Drosophila suzukii under simulated winter conditions of the Pacific Northwest and seasonal field trapping in five primary regions of small and stone fruit production in the United States. Pest Manag. Sci. 2011, 67, 1368–1374. [Google Scholar] [CrossRef]
- Lasa, R.; Tadeo, E.; Toledo-Hernandez, R.A.; Carmona, L.; Lima, I.; Williams, T. Improved capture of Drosophila suzukii by a trap baited with two attractants in the same device. PLoS ONE 2017, 12, e0188350. [Google Scholar] [CrossRef] [PubMed]
- Hendrichs, J.; Cooley, S.; Prokopy, R.J. Post-feeding bubbling behavior in fluid feeding Diptera: Concentration of crop contents by oral evaporation of excess water. Physiol. Entomol. 1992, 17, 153–161. [Google Scholar] [CrossRef]
- Burgess, E.R., IV; King, B.H. Insecticidal potential of two sugar alcohols to Musca domestica (Diptera: Muscidae). J. Econ. Entomol. 2017, 110, 2252–2258. [Google Scholar] [CrossRef] [PubMed]
- Le Bot, Y.; Gouy, P.A. Polyols from starch. In Handbook of Starch Hydrolysis Products and Their Derivatives; Dziedzic, S.Z., Kearsley, M.W., Eds.; Springer: Boston, MA, USA, 1995; pp. 155–177. [Google Scholar]
- Crawley, M.J. GLIM for Ecologists; Blackwell: Oxford, UK, 1993. [Google Scholar]
- Agresti, A. An Introduction to Categorical Data Analysis; John Wiley: New York, NY, USA, 1996; p. 290. [Google Scholar]
- Jamovi—Jamovi Project Version 0.9 [Computer Software]. Available online: https://www.jamovi.org/about.html (accessed on 7 February 2019).
- Sampson, B.J.; Werle, C.T.; Stringer, S.J.; Adamczyk, J.J. Ingestible insecticides for spotted wing Drosophila control: A polyol, erythritol, and an insect growth regulator, lufenuron. J. Appl. Entomol. 2016, 141, 8–18. [Google Scholar] [CrossRef]
- O’Donnell, S.; Baudier, K.; Marenda, D. Non-nutritive polyol sweeteners differ in insecticidal activity when ingested by adult Drosophila melanogaster (Diptera: Drosophilidae). J. Insect Sci. 2016, 16, 47. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.P.; Terra, W.R. An α-glucosidase from perimicrovillar membranes of Dysdercus peruvianus (Hemiptera: Pyrrhocoridae) midgut cells. Purification and properties. Insect Biochem. Mol. Biol. 1995, 25, 487–494. [Google Scholar] [CrossRef]
- Tang, S.B.; Lee, J.C.; Jung, J.K.; Choi, M.Y. Effect of erythritol formulation on the mortality, fecundity and physiological excretion in Drosophila suzukii. J. Insect Physiol. 2017, 101, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Fowles, J.R.; Banton, M.I.; Pottenger, L.H. A toxicological review of the propylene glycols. Crit. Rev. Toxicol. 2013, 43, 363–390. [Google Scholar] [CrossRef]
- Lim, T.Y.; Poole, R.L.; Pageler, N.M. Propylene glycol toxicity in children. J. Pediatr. Pharmacol. Ther. 2014, 19, 277–282. [Google Scholar]
- Zar, T.; Graeber, C.; Perazella, M.A. Recognition, treatment, and prevention of propylene glycol toxicity. Semin. Dialysis. 2007, 20, 217–219. [Google Scholar] [CrossRef]
- Uche, E.M.; Arowolo, R.O.; Akinyemi, J.O. Toxic effects of glycerol in Swiss albino rats. Res. Comm. Chem. Pathol. Pharmacol. 1987, 56, 125–128. [Google Scholar]
- Daltona, P.; Soretha, B.; Mautea, V.; Novaleskia, C.; Bantonb, M. Lack of respiratory and ocular effects following acute propylene glycol exposure in healthy humans. Inhal. Toxicol. 2018, 30, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Staerkle, M.; Kölliker, M. Maternal food regurgitation to nymphs in earwigs (Forficula auricularia). Ethology 2008, 114, 844–850. [Google Scholar]
- Horn, C.C. Why is the neurobiology of nausea and vomiting so important? Appetite 2008, 50, 430–434. [Google Scholar] [CrossRef] [PubMed]
- Hendrichs, J.; Fletcher, B.S.; Prokopy, R.J. Feeding behavior of Rhagoletis pomonella flies (Diptera: Tephritidae): Effect of initial food quantity and quality on food foraging, handling costs, and bubbling. J. lnsect Behav. 1993, 6, 43–64. [Google Scholar] [CrossRef]
- Stoffolano, J.G., Jr.; Acaron, A.; Conway, M. “Bubbling” or droplet regurgitation in both sexes of adult Phormia regina (Diptera: Calliphoridae) fed various concentrations of sugar and protein solutions. Ann. Entomol. Soc. Am. 2008, 101, 964–970. [Google Scholar] [CrossRef]
- Desurmont, G.A.; Köhler, A.; Maag, D.; Laplanche, D.; Xu, H.; Baumann, J.; Demairé, C.; Devenoges, D.; Glavan, M.; Mann, L.; Turlings, T.C. The spitting image of plant defenses: Effects of plant secondary chemistry on the efficacy of caterpillar regurgitant as an anti-predator defense. Ecol. Evol. 2017, 7, 6304–6313. [Google Scholar] [CrossRef]
- Koseki, T.; Koganezawa, M.; Furuyama, A.; Isono, K.; Shimada, I. A specific receptor site for glycerol, a new sweet tastant for Drosophila: Structure–taste relationship of glycerol in the labellar sugar receptor cell. Chem. Senses 2004, 29, 703–711. [Google Scholar] [CrossRef]
- Burke, C.J.; Waddell, S. Remembering nutrient quality of sugar in Drosophila. Curr. Biol. 2011, 21, 746–750. [Google Scholar] [CrossRef]
- Fujii, S.; Yavuz, A.; Slone, J.; Jagge, C.; Song, X.; Amrein, H. Drosophila sugar receptors in sweet taste perception, olfaction, and internal nutrient sensing. Curr. Biol. 2015, 25, 621–627. [Google Scholar] [CrossRef]
- Fontellas, T.M.d.L.; Zucoloto, F.S. Nutritive value of diets with different carbohydrates for adult Anastrepha obliqua (Macquart) (Diptera, Tephritidae). Rev. Bras. Zool. 1999, 16, 1135–1147. [Google Scholar] [CrossRef]
- Medeiros, L.; Zucoloto, F.S. Nutritional balancing in fruit flies: Performance of wild adult females of Anastrepha obliqua (Diptera: Tephritidae) fed on single-food or food-pair treatments. J. Insect Physiol. 2006, 52, 1121–1127. [Google Scholar] [CrossRef] [PubMed]
- Cowles, R.S.; Rodriguez-Saona, C.; Holdcraft, R.; Loeb, G.M.; Elsensohn, J.E.; Hesler, S.P. Sucrose improves insecticide activity against Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2015, 108, 640–653. [Google Scholar] [CrossRef] [PubMed]
- Aluja, M.; Guillén, L.; Jácome, I.; Righini, N. Long term feeding patterns highlight preference for sucrose in the fruit fly Anastrepha serpentina when given a choice over other more nutritious food sources. J. Insect Behav. 2016, 29, 719–734. [Google Scholar] [CrossRef]
- Muüller, G.C.; Kravchenko, V.D.; Schlein, Y. Decline of Anopheles sergentii and Aedes caspius populations following presentation of attractive, toxic (Spinosad), sugar bait stations in an oasis. J. Am. Mosq. Control Assoc. 2008, 24, 147–149. [Google Scholar] [CrossRef]
- Muüller, G.C.; Beier, J.C.; Traore, S.F.; Toure, M.B.; Traore, M.M.; Bah, S.; Doumbia, S.; Schlein, Y. Successful field trial of attractive toxic sugar bait (ATSB)plant-spraying methods against malaria vectors in the Anopheles gambiaecomplex in Mali, West Africa. Malaria J. 2010, 9, 210. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Fleischer, F.; Pérez-Staples, D.; Cabrera-Mireles, H.; Montoya, P.; Liedo, P. Long-term attraction and toxic effects of tephritid insecticide-bait mixtures by applying the Torricelli’s barometer principle in a trapping device. Pest Manag. Sci. 2016, 72, 1346–1349. [Google Scholar] [CrossRef] [PubMed]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-Fleischer, F.; Arredondo, J.; Lasa, R.; Bonilla, C.; Debernardi, D.; Pérez-Staples, D.; Williams, T. Sickly Sweet: Insecticidal Polyols Induce Lethal Regurgitation in Dipteran Pests. Insects 2019, 10, 53. https://doi.org/10.3390/insects10020053
Díaz-Fleischer F, Arredondo J, Lasa R, Bonilla C, Debernardi D, Pérez-Staples D, Williams T. Sickly Sweet: Insecticidal Polyols Induce Lethal Regurgitation in Dipteran Pests. Insects. 2019; 10(2):53. https://doi.org/10.3390/insects10020053
Chicago/Turabian StyleDíaz-Fleischer, Francisco, José Arredondo, Rodrigo Lasa, Carlos Bonilla, Diana Debernardi, Diana Pérez-Staples, and Trevor Williams. 2019. "Sickly Sweet: Insecticidal Polyols Induce Lethal Regurgitation in Dipteran Pests" Insects 10, no. 2: 53. https://doi.org/10.3390/insects10020053
APA StyleDíaz-Fleischer, F., Arredondo, J., Lasa, R., Bonilla, C., Debernardi, D., Pérez-Staples, D., & Williams, T. (2019). Sickly Sweet: Insecticidal Polyols Induce Lethal Regurgitation in Dipteran Pests. Insects, 10(2), 53. https://doi.org/10.3390/insects10020053