Analysis of Salivary Glands and Saliva from Aedes albopictus and Aedes aegypti Infected with Chikungunya Viruses


Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus and Cell Culture
2.2. Mosquitoes
2.3. Infectious Virus Titration by Plaque Assay
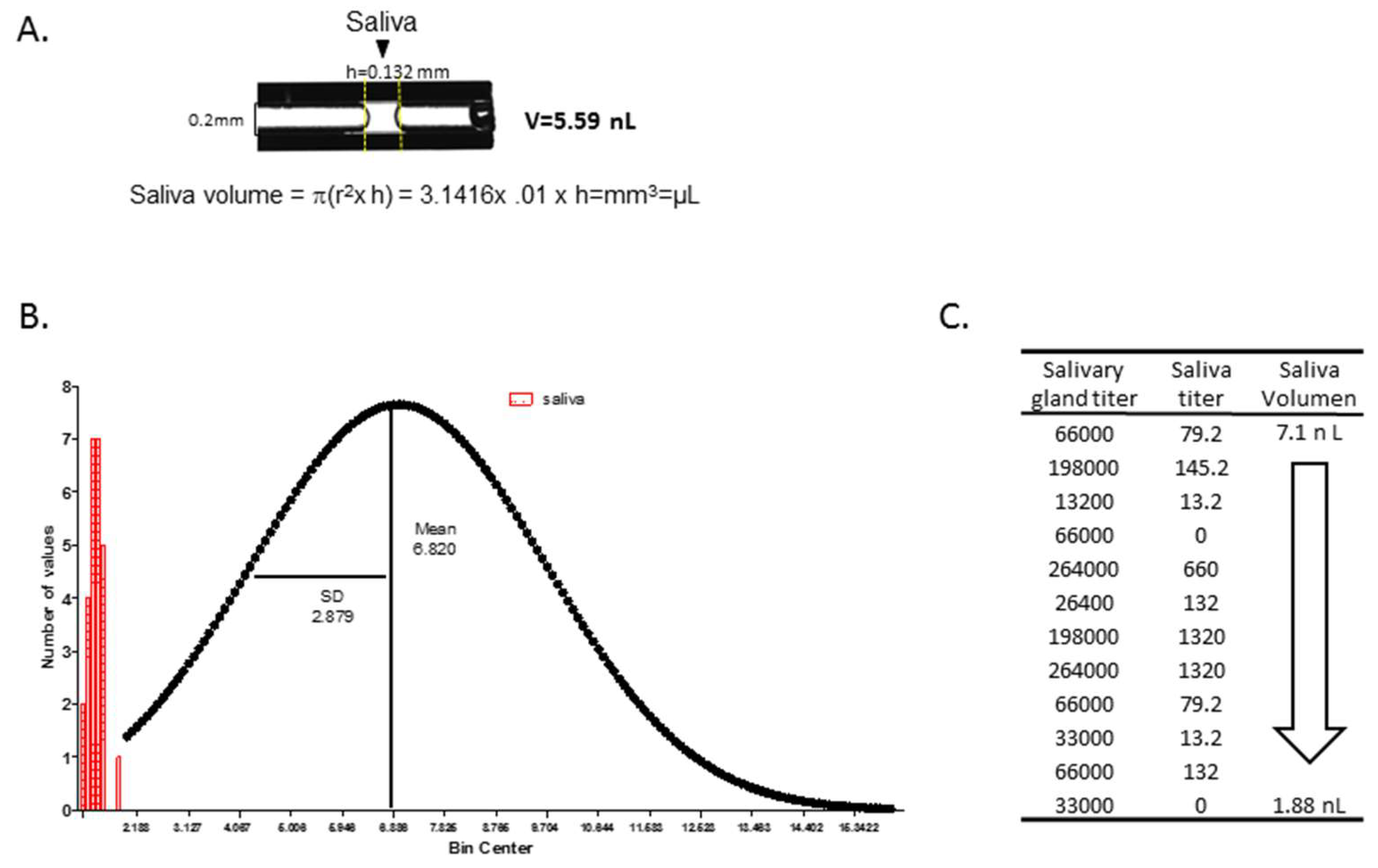
2.4. Saliva Collection and Salivary Gland Dissections
2.5. Statistical Analyses
3. Results
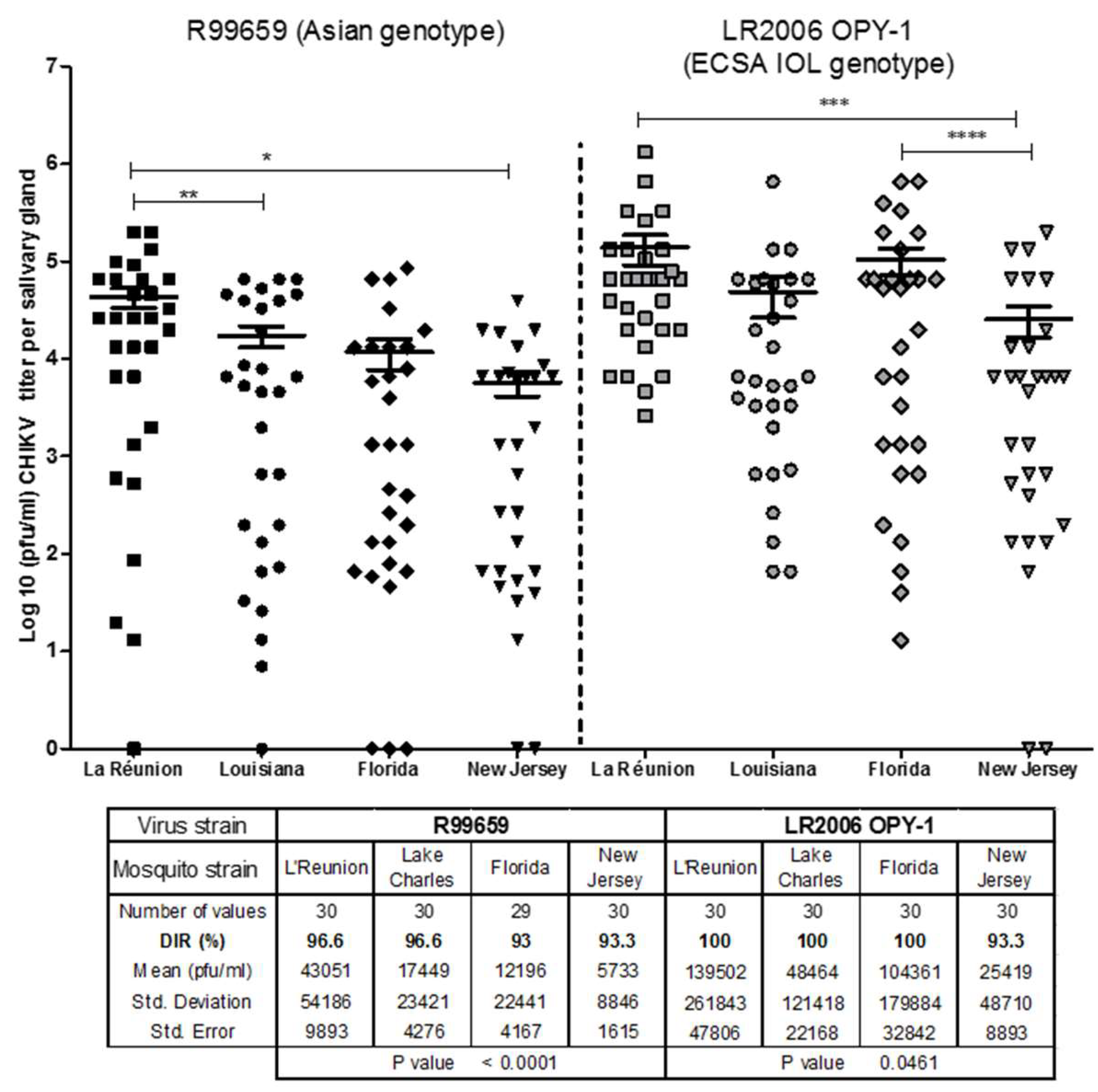
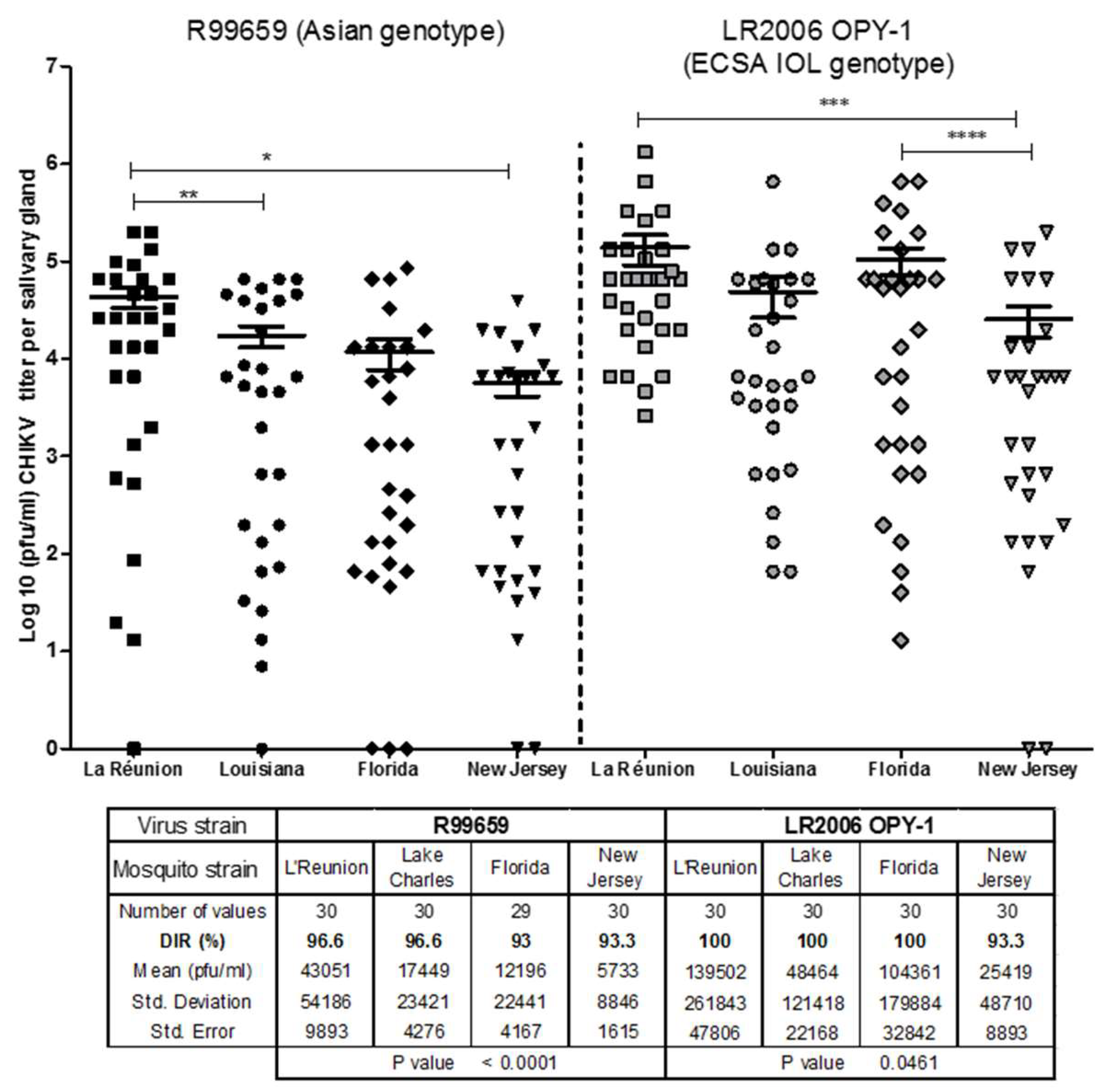
3.1. Dissemination of CHIKV R99659 and LR2006 OPY1 to Salivary Glands of Ae. albopictus Mosquito Strains
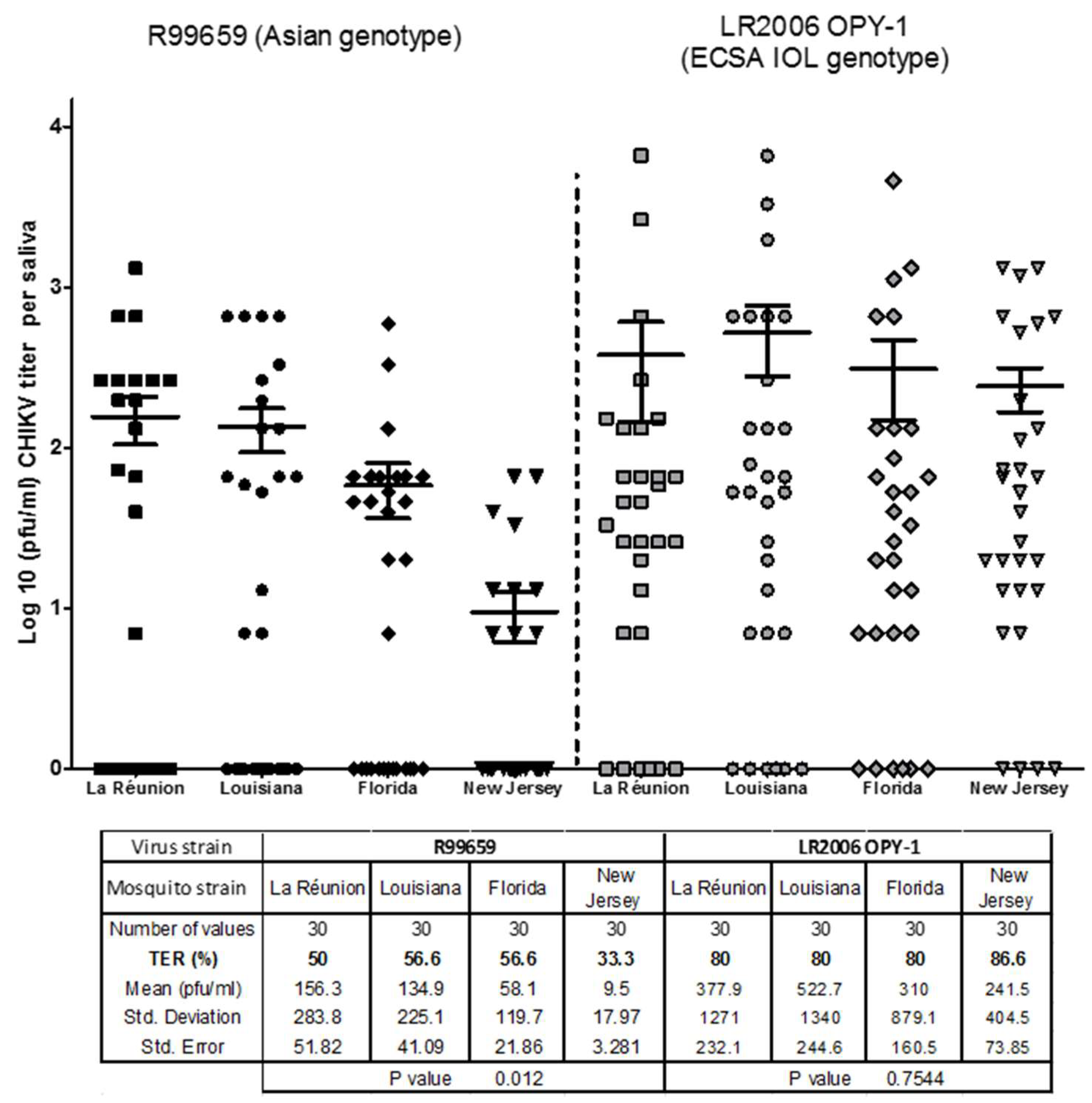
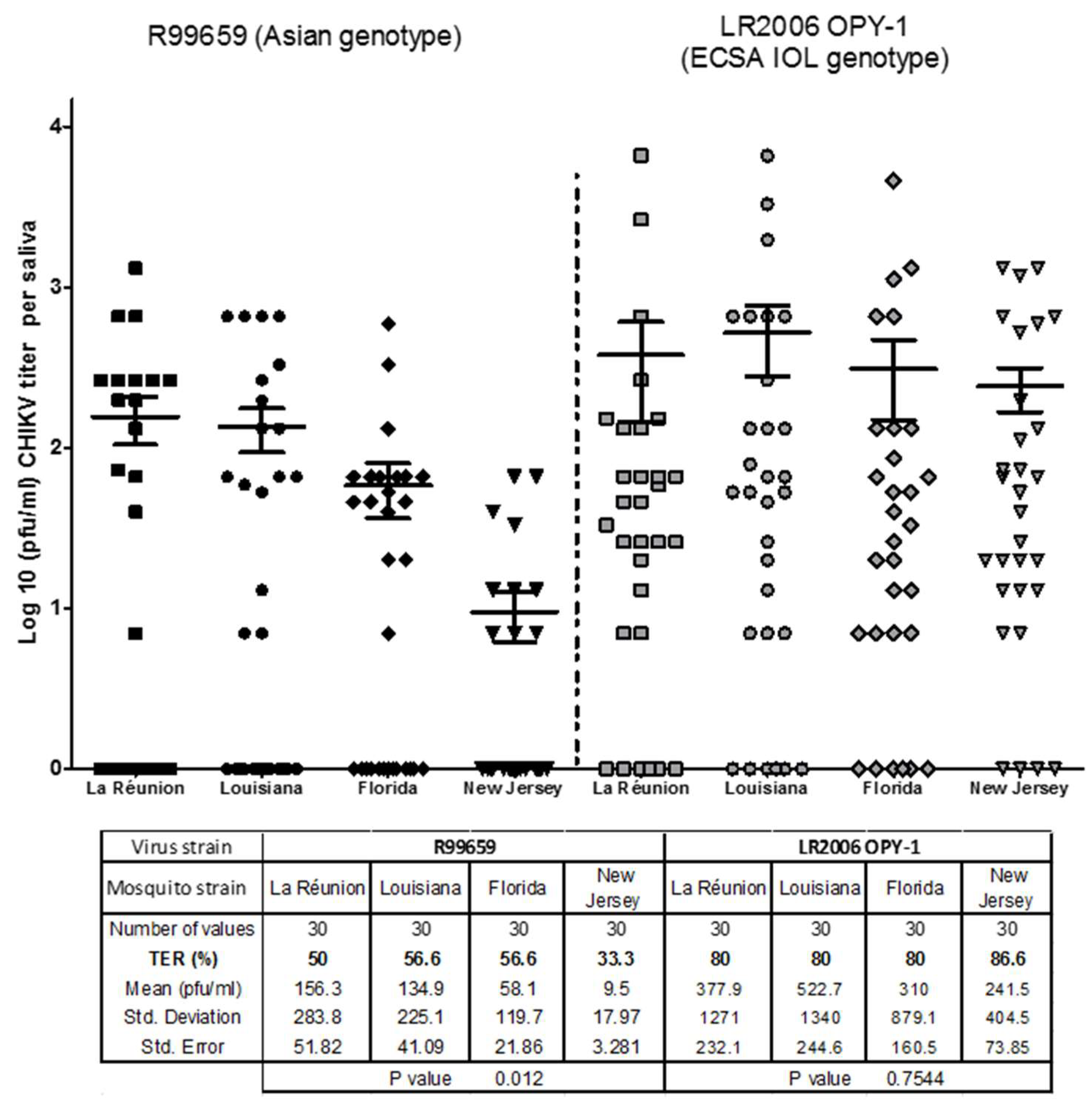
3.2. CHIKV R99659 and LR2006 OPY1 Viruses in Saliva of Ae. albopictus Mosquito Strains
3.3. Dissemination and Transmission Potential of CHIKV R99659 and LR2006 OPY1 in Ae. aegypti (Poza Rica)
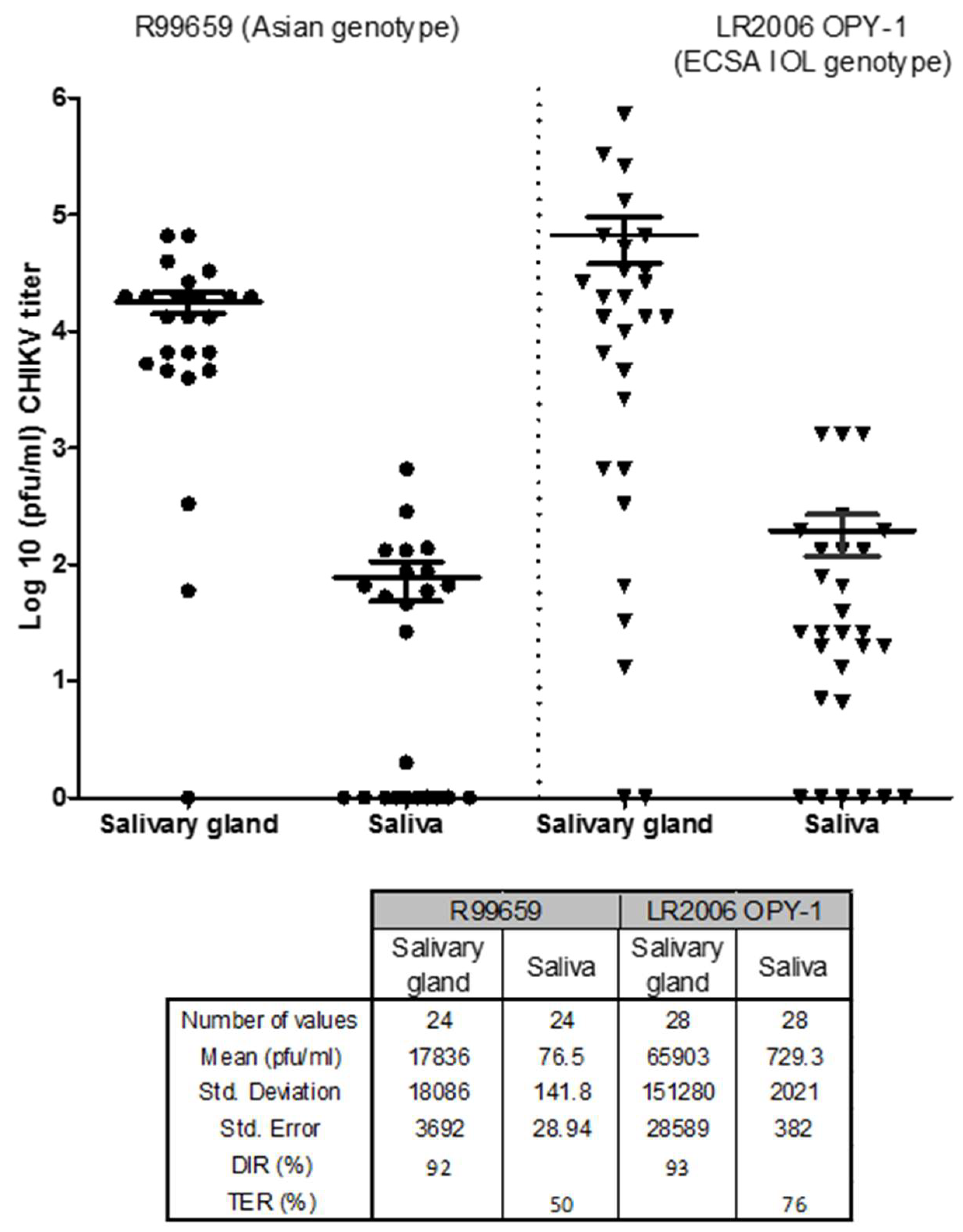
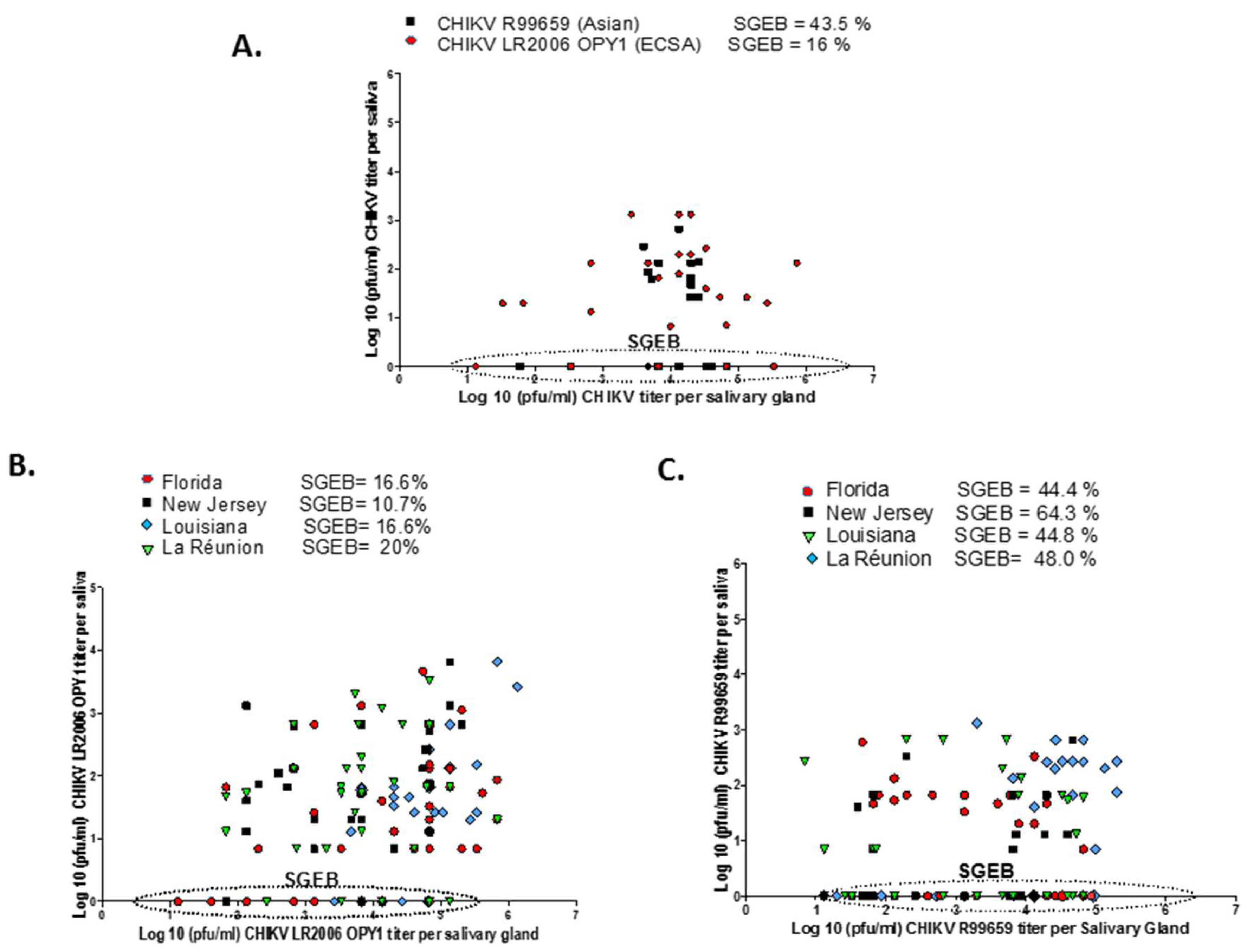
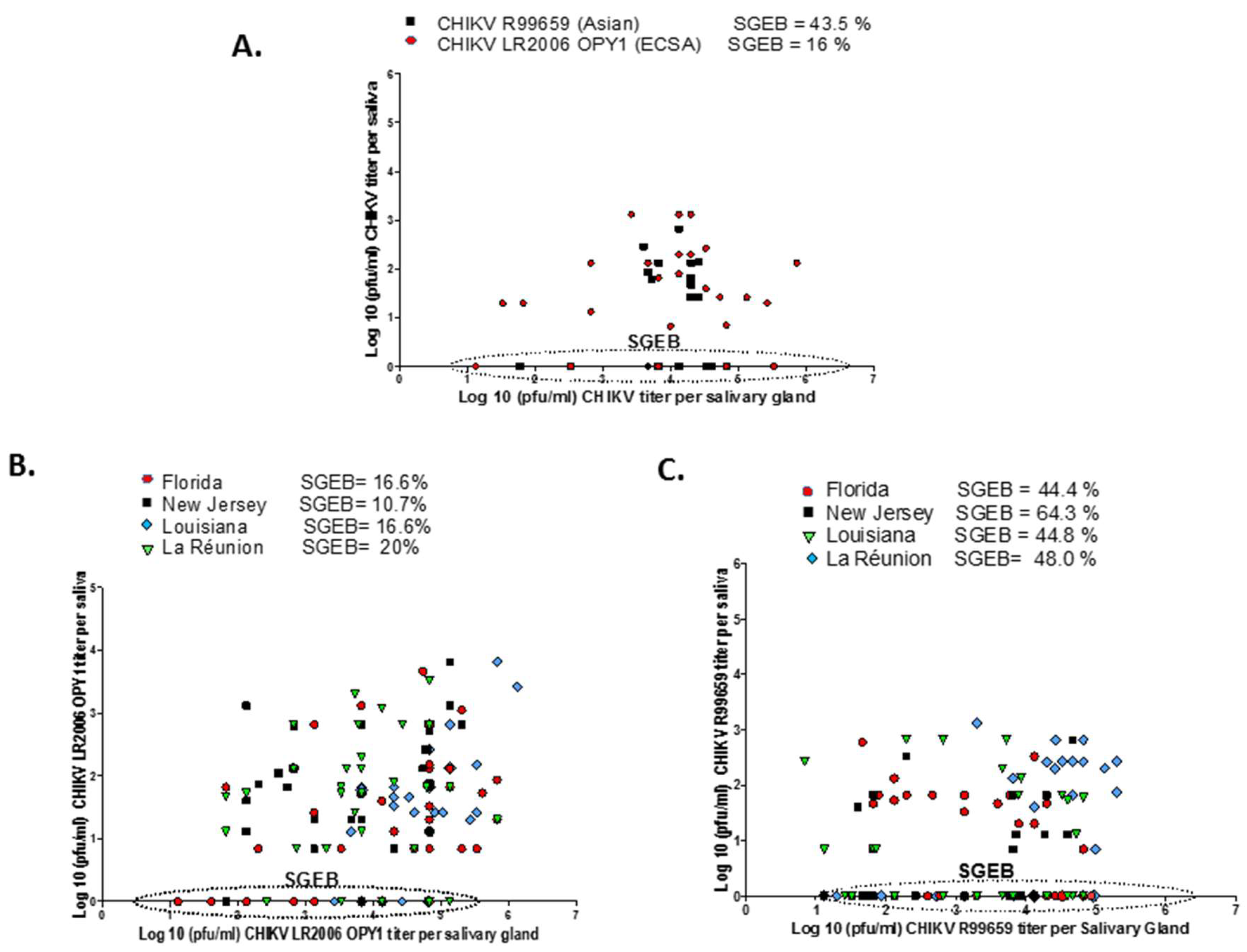
3.4. Relationship between CHIKV Infectious Particles in Saliva and Salivary Glands
3.5. Vector Competence of Ae. aegypti (Poza Rica) and Ae. albopictus Strains for R99659 versus LR2006 OPY1 Viruses
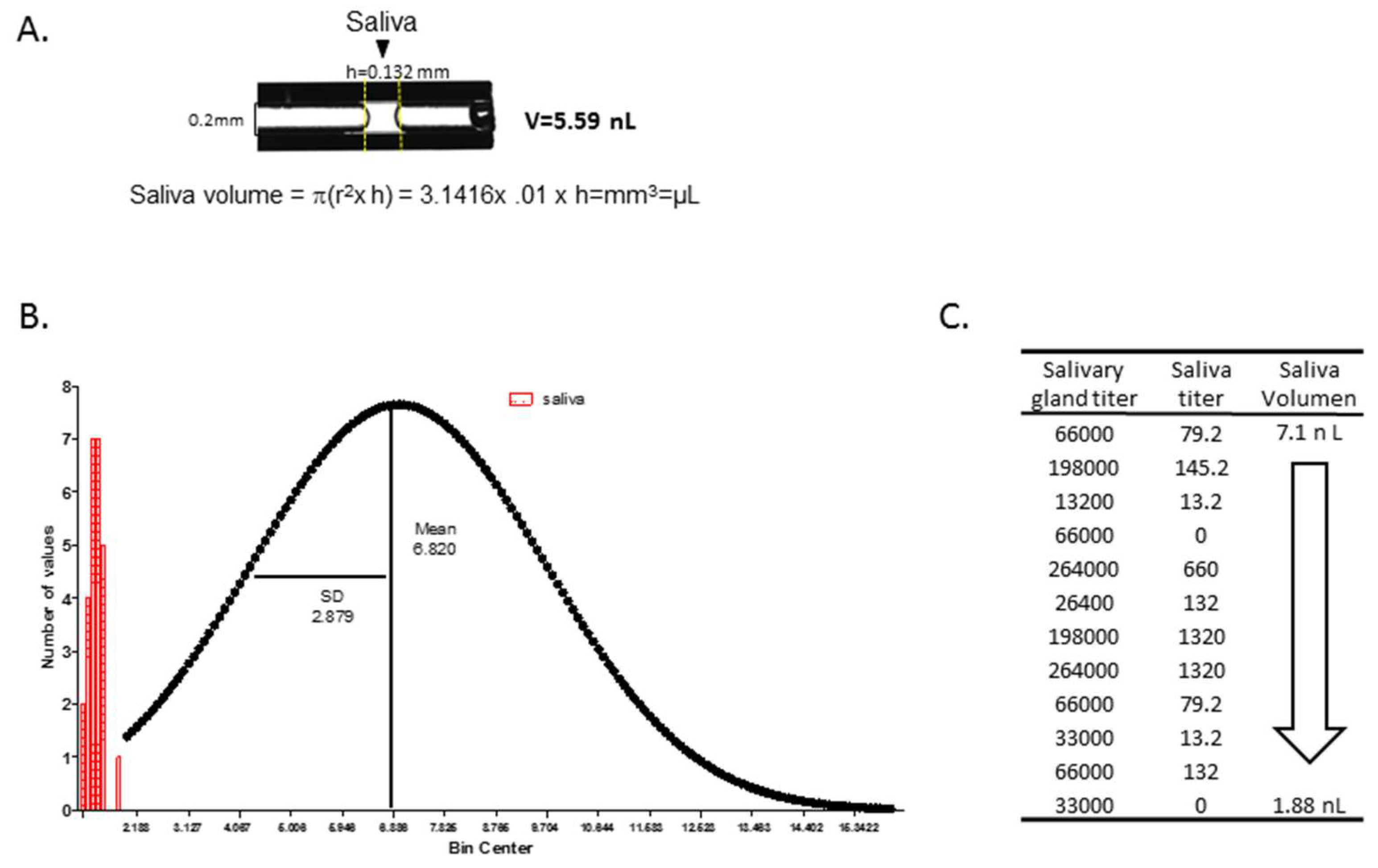
3.6. Measurement of Salivation Using Ae. aegypti as A Model for Determining Saliva Volume from An Aedes Species
4. Discussion
5. Conclusions
- (1)
- We infected the Ae. albopictus strains and Ae. aegypti with blood meals containing 5–7 × 107 PFU/mL of R99659 (Asian genotype) and LR2006 OPY1 (ECSA-IOL genotype) and analyzed the mosquitoes nine days later to detect maximum infection of their salivary glands and saliva.
- (2)
- All four Ae. albopictus strains were highly susceptible to LR2006 OPY1 and R99659 viruses and their CHIKV disseminated infection rates (DIR) were statistically similar (p = 0.3916).
- (3)
- We demonstrated here that the TER in Ae. aegypti and Ae. albopictus vary according to their geographic origins of the mosquitoes and the CHIKV strain contributing to a salivary gland barrier for releasing R99659 virus into saliva.
- (4)
- Virus titers in salivary glands did not correlate with saliva volume, but Ae. aegypti expectorated on average a volume of 6.8 nL/mosquito.
- (5)
- If introduced, LR2006 OPY1 virus could increase the risk of CHIKV transmission by both Aedes species in the western hemisphere.
Author Contributions
Funding
Conflicts of Interest
References
- Strauss, J.H.; Strauss, E.G. The alphaviruses: Gene expression, replication, and evolution. Microbiol. Rev. 1994, 58, 491–562. [Google Scholar] [PubMed]
- Weaver, S.C.; Lecuit, M. Chikungunya virus and the global spread of a mosquito-borne disease. N. Engl. J. Med. 2015, 372, 1231–1239. [Google Scholar] [CrossRef]
- Riswari, S.F.; Ma’roef, C.N.; Djauhari, H.; Kosasih, H.; Perkasa, A.; Yudhaputri, F.A.; Artika, I.M.; Williams, M.; van der Ven, A.; Myint, K.S.; et al. Study of viremic profile in febrile specimens of chikungunya in Bandung, Indonesia. J. Clin. Virol. 2016, 74, 61–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Duijl-Richter, M.K.; Hoornweg, T.E.; Rodenhuis-Zybert, I.A.; Smit, J.M. Early events in chikungunya virus infection-from virus cell binding to membrane fusion. Viruses 2015, 7, 3647–3674. [Google Scholar] [CrossRef] [PubMed]
- Rai, K.S. Aedes albopictus in the Americas. Annu. Rev. Entomol. 1991, 36, 459–484. [Google Scholar] [CrossRef]
- Powers, A.M.; Brault, A.C.; Tesh, R.B.; Weaver, S.C. Re-emergence of Chikungunya and O’nyong-nyong viruses: Evidence for distinct geographical lineages and distant evolutionary relationships. J. Gen. Virol. 2000, 81 Pt 2, 471–479. [Google Scholar] [CrossRef]
- Volk, S.M.; Chen, R.; Tsetsarkin, K.A.; Adams, A.P.; Garcia, T.I.; Sall, A.A.; Nasar, F.; Schuh, A.J.; Holmes, E.C.; Higgs, S.; et al. Genome-scale phylogenetic analyses of chikungunya virus reveal independent emergences of recent epidemics and various evolutionary rates. J. Virol. 2010, 84, 6497–6504. [Google Scholar] [CrossRef]
- Dupont-Rouzeyrol, M.; Caro, V.; Guillaumot, L.; Vazeille, M.; D’Ortenzio, E.; Thiberge, J.M.; Baroux, N.; Gourinat, A.C.; Grandadam, M.; Failloux, A.B. Chikungunya virus and the mosquito vector Aedes aegypti in New Caledonia (South Pacific Region). Vector Borne Zoonotic Dis. 2012, 12, 1036–1041. [Google Scholar] [CrossRef]
- Tsetsarkin, K.A.; Vanlandingham, D.L.; McGee, C.E.; Higgs, S. A single mutation in chikungunya virus affects vector specificity and epidemic potential. PLoS Pathog. 2007, 3, 0030201. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.A.; McGee, C.E.; Volk, S.M.; Vanlandingham, D.L.; Weaver, S.C.; Higgs, S. Epistatic roles of E2 glycoprotein mutations in adaption of chikungunya virus to Aedes albopictus and Ae. aegypti mosquitoes. PLoS ONE 2009, 4, 0006835. [Google Scholar] [CrossRef]
- Vazeille, M.; Moutailler, S.; Coudrier, D.; Rousseaux, C.; Khun, H.; Huerre, M.; Thiria, J.; Dehecq, J.S.; Fontenille, D.; Schuffenecker, I.; et al. Two Chikungunya isolates from the outbreak of La Reunion (Indian Ocean) exhibit different patterns of infection in the mosquito, Aedes albopictus. PLoS ONE 2007, 2, 0001168. [Google Scholar] [CrossRef] [PubMed]
- Schuffenecker, I.; Iteman, I.; Michault, A.; Murri, S.; Frangeul, L.; Vaney, M.C.; Lavenir, R.; Pardigon, N.; Reynes, J.M.; Pettinelli, F.; et al. Genome microevolution of chikungunya viruses causing the Indian Ocean outbreak. PLoS Med. 2006, 3, 23. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.A.; Weaver, S.C. Sequential adaptive mutations enhance efficient vector switching by Chikungunya virus and its epidemic emergence. PLoS Pathog. 2011, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Waggoner, J.J.; Pinsky, B.A. How great is the threat of chikungunya virus? Expert Rev. Anti Infect. Ther. 2015, 13, 291–293. [Google Scholar] [CrossRef] [PubMed]
- Morrison, T.E. Reemergence of chikungunya virus. J. Virol. 2014, 88, 11644–11647. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.V.J., Jr.; Ludwig-Begall, L.F.; de Oliveira-Filho, E.F.; Oliveira, R.A.S.; Duraes-Carvalho, R.; Lopes, T.R.R.; Silva, D.E.; Gil, L.H. A scoping review of Chikungunya virus infection: Epidemiology, clinical characteristics, viral co-circulation complications, and control. Acta Trop. 2018, 6, 30937–30939. [Google Scholar] [CrossRef] [PubMed]
- Gratz, N.G. Critical review of the vector status of Aedes albopictus. Med. Vet. Entomol. 2004, 18, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, K.E.; Lessler, J.; Moloney, R.M.; Kmush, B.; Cummings, D.A. Incubation periods of mosquito-borne viral infections: A systematic review. Am. J. Trop Med. Hyg. 2014, 90, 882–891. [Google Scholar] [CrossRef]
- Vanlandingham, D.L.; Higgs, S.; Huang, Y.J. Aedes albopictus (Diptera: Culicidae) and mosquito-borne viruses in the United States. J. Med. Entomol. 2016, 53, 1024–1028. [Google Scholar] [CrossRef]
- Thavara, U.; Tawatsin, A.; Pengsakul, T.; Bhakdeenuan, P.; Chanama, S.; Anantapreecha, S.; Molito, C.; Chompoosri, J.; Thammapalo, S.; Sawanpanyalert, P.; et al. Outbreak of chikungunya fever in Thailand and virus detection in field population of vector mosquitoes, Aedes aegypti (L.) and Aedes albopictus Skuse (Diptera: Culicidae). Southeast. Asian J. Trop Med. Public Health. 2009, 40, 951–962. [Google Scholar]
- Vera-Maloof, F.Z.; Saavedra-Rodriguez, K.; Elizondo-Quiroga, A.E.; Lozano-Fuentes, S.; Black IV, W.C. Coevolution of the Ile1,016 and Cys1,534 mutations in the voltage gated sodium channel gene of Aedes aegypti in Mexico. PLoS Negl. Trop Dis. 2015, 9, e0004263. [Google Scholar] [CrossRef]
- Bosio, C.F.; Beaty, B.J.; Black, W.C., IV. Quantitative genetics of vector competence for dengue-2 virus in Aedes aegypti. Am. J. Trop Med. Hyg. 1998, 59, 965–970. [Google Scholar] [CrossRef]
- Sladowski, D.; Steer, S.J.; Clothier, R.H.; Balls, M. An improved MTT assay. J. Immunol. Methods. 1993, 157, 203–207. [Google Scholar] [CrossRef]
- Takeuchi, H.; Baba, M.; Shigeta, S. An application of tetrazolium (MTT) colorimetric assay for the screening of anti-herpes simplex virus compounds. J. Virol. Methods. 1991, 33, 61–71. [Google Scholar] [CrossRef]
- Dubrulle, M.; Mousson, L.; Moutailler, S.; Vazeille, M.; Failloux, A.B. Chikungunya virus and Aedes mosquitoes: Saliva is infectious as soon as two days after oral infection. PLoS ONE 2009, 4, 0005895. [Google Scholar] [CrossRef]
- Hurlbut, H.S. Mosquito salivation and virus transmission. Am. J. Trop. Med. Hyg. 1966, 15, 989–993. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vargas, I.; Olson, K.E.; Black, W.C., IV. Genetics of salivary gland infection barriers in population of Aedes aegypti. Infect. Genet. Evol. (In preparation).
- Vega-Rua, A.; Zouache, K.; Girod, R.; Failloux, A.B.; Lourenco-de-Oliveira, R. High level of vector competence of Aedes aegypti and Aedes albopictus from ten American countries as a crucial factor in the spread of Chikungunya virus. J. Virol. 2014, 88, 6294–6306. [Google Scholar] [CrossRef] [PubMed]
- Alto, B.W.; Wiggins, K.; Eastmond, B.; Velez, D.; Lounibos, L.P.; Lord, C.C. Transmission risk of two chikungunya lineages by invasive mosquito vectors from Florida and the Dominican Republic. PLoS Negl. Trop Dis. 2017, 11, e0005724. [Google Scholar] [CrossRef]
- Honorio, N.A.; Wiggins, K.; Camara, D.C.P.; Eastmond, B.; Alto, B.W. Chikungunya virus vector competency of Brazilian and Florida mosquito vectors. PLoS Negl. Trop. Dis. 2018, 12, e0006521. [Google Scholar] [CrossRef]
- Ngoagouni, C.; Kamgang, B.; Kazanji, M.; Paupy, C.; Nakoune, E. Potential of Aedes aegypti and Aedes albopictus populations in the Central African Republic to transmit enzootic chikungunya virus strains. Parasit Vectors. 2017, 10, 164. [Google Scholar] [CrossRef]
- Lounibos, L.P.; Kramer, L.D. Invasiveness of Aedes aegypti and Aedes albopictus and vectorial capacity for chikungunya virus. J. Infect. Dis. 2016, 214 (Suppl. 5), S453–S458. [Google Scholar] [CrossRef]
- Reiskind, M.H.; Pesko, K.; Westbrook, C.J.; Mores, CN. Susceptibility of Florida mosquitoes to infection with chikungunya virus. Am. J. Trop. Med. Hyg. 2008, 78, 422–425. [Google Scholar] [CrossRef] [PubMed]
- Ruckert, C.; Ebel, G.D. How do virus-mosquito interactions lead to viral emergence? Trends Parasitol. 2018, 34, 310–321. [Google Scholar] [CrossRef]
- Forrester, N.L.; Coffey, L.L.; Weaver, S.C. Arboviral bottlenecks and challenges to maintaining diversity and fitness during mosquito transmission. Viruses 2014, 6, 3991–4004. [Google Scholar] [CrossRef] [PubMed]
- Beaty, B.J.; Holterman, M.; Tabachnick, W.; Shope, R.E.; Rozhon, E.J.; Bishop, D.H. Molecular basis of bunyavirus transmission by mosquitoes: Role of the middle-sized RNA segment. Science 1981, 211, 1433–1435. [Google Scholar] [CrossRef] [PubMed]
- Grimstad, P.R.; Paulson, S.L.; Craig, G.B., Jr. Vector competence of Aedes hendersoni (Diptera: Culicidae) for La Crosse virus and evidence of a salivary-gland escape barrier. J. Med. Entomol. 1985, 22, 447–453. [Google Scholar] [CrossRef]
- Jupp, P.G. Culex theileri and Sindbis virus; salivary glands infection in relation to transmission. J. Am. Mosq. Control Assoc. 1985, 1, 374–376. [Google Scholar]
- Paulson, S.L.; Poirier, S.J.; Grimstad, P.R.; Craig, G.B., Jr. Vector competence of Aedes hendersoni (Diptera: Culicidae) for La Crosse virus: Lack of impaired function in virus-infected salivary glands and enhanced virus transmission by sporozoite-infected mosquitoes. J. Med. Entomol. 1992, 29, 483–488. [Google Scholar] [CrossRef]
- Turell, M.J.; Britch, S.C.; Aldridge, R.L.; Kline, D.L.; Boohene, C.; Linthicum, K.J. Potential for mosquitoes (Diptera: Culicidae) from Florida to transmit Rift Valley fever virus. J. Med. Entomol. 2013, 50, 1111–1117. [Google Scholar] [CrossRef] [PubMed]
- Gaidamovich, S.Y.; Khutoretskaya, N.V.; Lvova, A.I.; Sveshnikova, N.A. Immunofluorescent staining study of the salivary glands of mosquitoes infected with group A arboviruses. Intervirology 1973, 1, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Janzen, H.G.; Rhodes, A.J.; Doane, F.W. Chikungunya virus in salivary glands of Aedes aegypti (L.): An electron microscope study. Can. J. Microbiol. 1970, 16, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Salazar, M.I.; Richardson, J.H.; Sanchez-Vargas, I.; Olson, K.E.; Beaty, B.J. Dengue virus type 2: Replication and tropisms in orally infected Aedes aegypti mosquitoes. BMC Microbiol. 2007, 7, 1471–2180. [Google Scholar] [CrossRef] [PubMed]
- Coffey, L.L.; Failloux, A.B.; Weaver, S.C. Chikungunya virus-vector interactions. Viruses 2014, 6, 4628–4663. [Google Scholar] [CrossRef] [PubMed]
- Kendrick, K.; Stanek, D.; Blackmore, C. Transmission of chikungunya virus in the continental United States—Florida, 2014. MMWR Morb. Mortal. Wkly. 2014, 63, 1137. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| R99659 CHIKV R99659 | |||
| Ae. albopictus | Dissemination Rates (DIR) | Transmission Efficiency Rate (TER) | Vector Competence (VC) |
| La Réunion | 96.6 | 50 | 48.3 |
| Louisiana | 96.6 | 56.6 | 54.7 |
| Florida | 93 | 56.6 | 52.7 |
| New Jersey | 93.3 | 33.3 | 31.1 |
| LR2006 OPY1 CHIKVOPY1 ZIKVOPY-1 | |||
| Ae. albopictus | Dissemination Rates (DIR) | Transmission Efficiency Rate (TER) | Vector Competence (VC) |
| La Réunion | 100 | 80 | 80 |
| Louisiana | 100 | 80 | 80 |
| Florida | 100 | 80 | 80 |
| New Jersey | 93.3 | 86.6 | 80.1 |
| Aedes aegypti (Poza Rica) | |||
| ZIKV | Dissemination Rates (DIR) | Transmission Efficiency Rate (TER) | Vector Competence (VC) |
| R99659 | 92 | 50 | 46 |
| LR2006 OPY1 | 93 | 76 | 70.6 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanchez-Vargas, I.; Harrington, L.C.; Black, W.C., IV; Olson, K.E. Analysis of Salivary Glands and Saliva from Aedes albopictus and Aedes aegypti Infected with Chikungunya Viruses. Insects 2019, 10, 39. https://doi.org/10.3390/insects10020039
Sanchez-Vargas I, Harrington LC, Black WC IV, Olson KE. Analysis of Salivary Glands and Saliva from Aedes albopictus and Aedes aegypti Infected with Chikungunya Viruses. Insects. 2019; 10(2):39. https://doi.org/10.3390/insects10020039
Chicago/Turabian StyleSanchez-Vargas, Irma, Laura C. Harrington, William C. Black, IV, and Ken E. Olson. 2019. "Analysis of Salivary Glands and Saliva from Aedes albopictus and Aedes aegypti Infected with Chikungunya Viruses" Insects 10, no. 2: 39. https://doi.org/10.3390/insects10020039
APA StyleSanchez-Vargas, I., Harrington, L. C., Black, W. C., IV, & Olson, K. E. (2019). Analysis of Salivary Glands and Saliva from Aedes albopictus and Aedes aegypti Infected with Chikungunya Viruses. Insects, 10(2), 39. https://doi.org/10.3390/insects10020039