Queen Recognition Signals in Two Primitively Eusocial Halictid Bees: Evolutionary Conservation and Caste-Specific Perception

Abstract
1. Introduction
2. Materials and Methods
2.1. Bee Collection
2.2. Chemical Analyses
2.3. Measurement of Physiological State
2.4. Electrophysiological Analyses
2.5. Circle Tube Bioassays
2.6. Statistics
3. Results
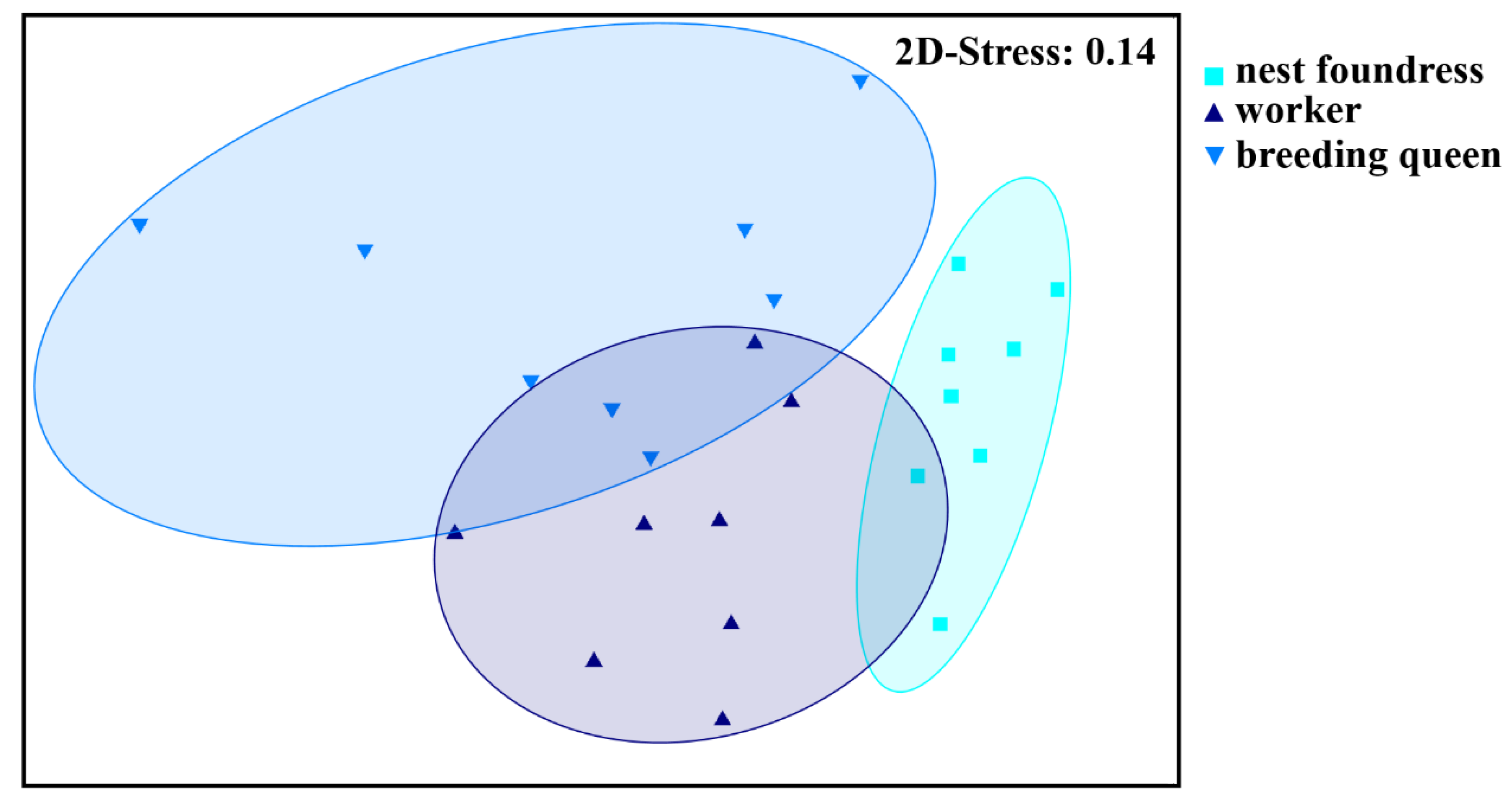
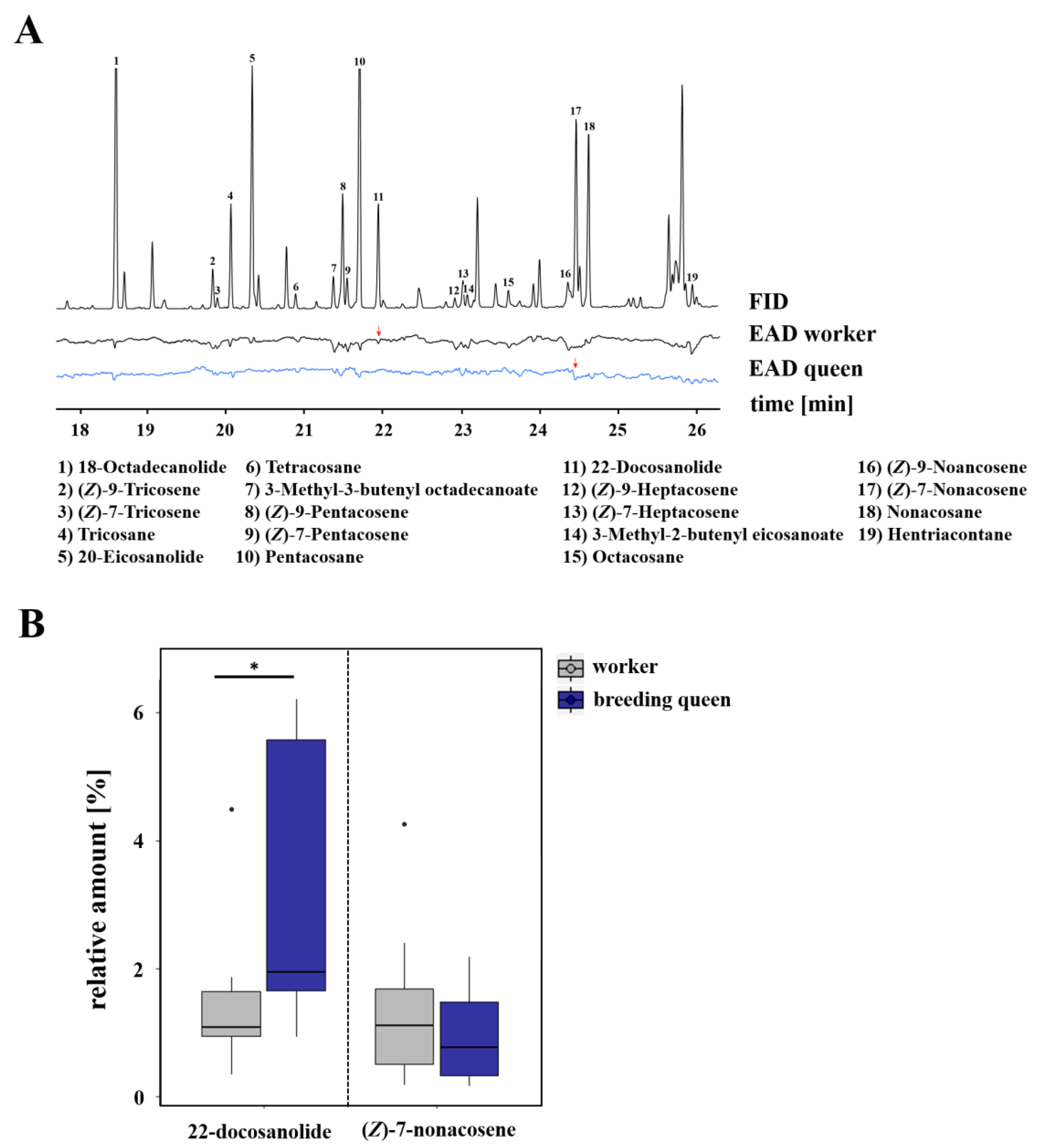
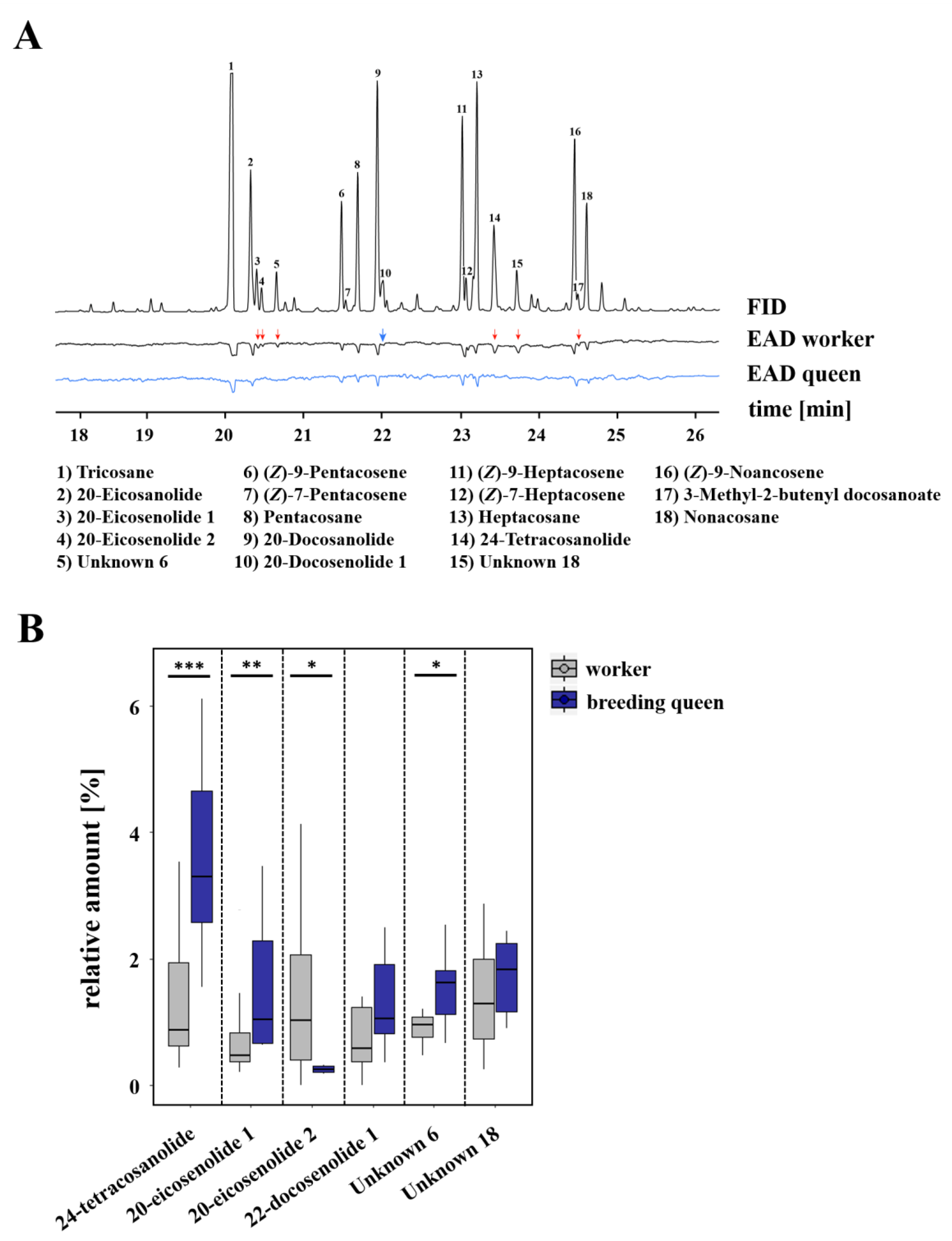
3.1. Caste-Specific and Female-Function-Specific Odor Profiles in L. pauxillum
3.2. Queen–Worker Interactions and Queen Recognition in L. pauxillum
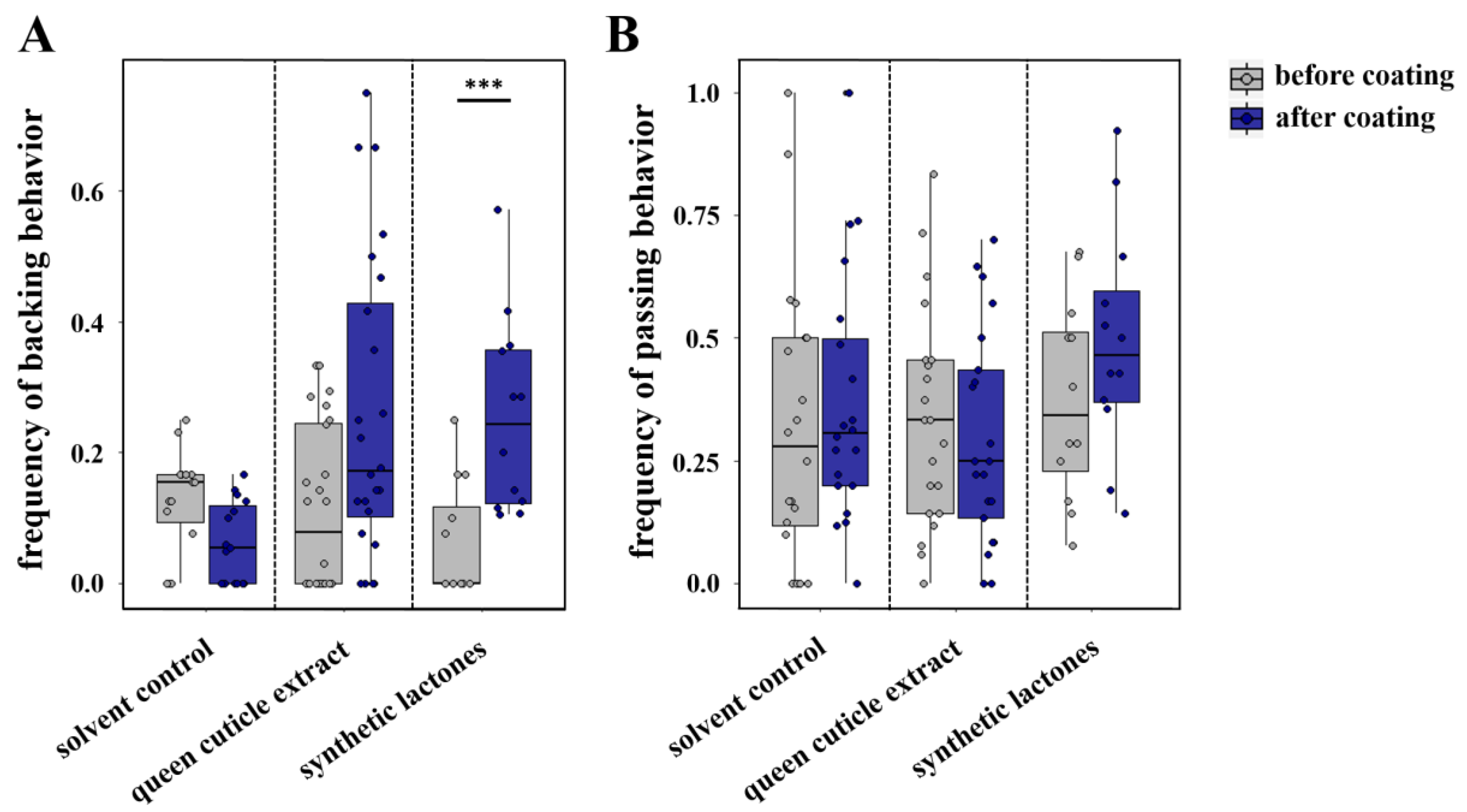
3.3. Caste-Specific Perception of Odor Signals
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wilson, E.O. The Insect Societies; Belknap Press of Harvard University Press: Cambridge, MA, USA, 1971. [Google Scholar]
- Ayasse, M.; Jarau, S. Chemical ecology of bumble bees. Annu. Rev. Entomol. 2014, 59, 299–319. [Google Scholar] [CrossRef] [PubMed]
- Kocher, S.D.; Grozinger, C.M. Cooperation, conflict, and the evolution of queen pheromones. J. Chem. Ecol. 2011, 37, 1263–1275. [Google Scholar] [CrossRef] [PubMed]
- Le Conte, Y.; Hefetz, A. Primer pheromones in social Hymenoptera. Annu. Rev. Entomol. 2008, 53, 523–542. [Google Scholar] [CrossRef] [PubMed]
- Monnin, T. Chemical recognition of reproductive status in social insects. Ann. Zool. Fenn. 2006, 43, 515–530. [Google Scholar]
- Oi, C.A.; van Zweden, J.S.; Oliveira, R.C.; Van Oystaeyen, A.; Nascimento, F.S.; Wenseleers, T. The origin and evolution of social insect queen pheromones: Novel hypotheses and outstanding problems. BioEssays 2015, 37, 808–821. [Google Scholar] [CrossRef]
- Rottler-Hoemann, A.M.; Schulz, S.; Ayasse, M. Nest wax triggers worker reproduction in the bumblebee Bombus terrestris. R. Soc. Open Sci. 2016, 3, 150599. [Google Scholar] [CrossRef]
- Holman, L.; Jørgensen, C.G.; Nielsen, J.; d’Ettorre, P. Identification of an ant queen pheromone regulating worker sterility. Proc. R. Soc. Lond. B Biol. Sci. 2010, 277, 3793–3800. [Google Scholar] [CrossRef]
- Holman, L.; Lanfear, R.; d’Ettorre, P. The evolution of queen pheromones in the ant genus Lasius. J. Evol. Biol. 2013, 26, 1549–1558. [Google Scholar] [CrossRef]
- Holman, L.; Hanley, B.; Millar, J.G. Highly specific responses to queen pheromone in three Lasius ant species. Behav. Ecol. Sociobiol. 2016, 70, 387–392. [Google Scholar] [CrossRef]
- Oi, C.A.; Millar, J.G.; van Zweden, J.S.; Wenseleers, T. Conservation of queen pheromones across two species of vespine wasps. J. Chem. Ecol. 2016, 42, 1175–1180. [Google Scholar] [CrossRef]
- Nunes, T.M.; Mateus, S.; Favaris, A.P.; Amaral, M.F.; Von Zuben, L.G.; Clososki, G.C.; Bento, J.M.S.; Oldroyd, B.P.; Silva, R.; Zucchi, R.; et al. Queen signals in a stingless bee: Suppression of worker ovary activation and spatial distribution of active compounds. Sci. Rep. 2014, 4, 7449. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.A.; Hölldobler, B.; Liebig, J. Cuticular hydrocarbons reliably identify cheaters and allow enforcement of altruism in a social insect. Curr. Biol. 2009, 19, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.A.; Millar, J.G.; Hanks, L.M.; Suarez, A.V. Experimental evidence that workers recognize reproductives through cuticular hydrocarbons in the ant Odontomachus brunneus. Behav. Ecol. Sociobiol. 2012, 66, 1267–1276. [Google Scholar] [CrossRef]
- Van Oystaeyen, A.; Oliveira, R.C.; Holman, L.; van Zweden, J.S.; Romero, C.; Oi, C.A.; d’Ettorre, P.; Khalesi, M.; Billen, J.; Wäckers, F.; et al. Conserved class of queen pheromones stops social insect workers from reproducing. Science 2014, 343, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Butler, C.G.; Callow, R.K.; Johnston, N.C. The isolation and synthesis of queen substance, 9-oxodec-trans-2-enoic acid, a honeybee pheromone. Proc. R. Soc. Lond. B Biol. Sci. 1962, 155, 417–432. [Google Scholar]
- Hoover, S.E.R.; Keeling, C.I.; Winston, M.L.; Slessor, K.N. The effect of queen pheromones on worker honey bee ovary development. Naturwissenschaften 2003, 90, 477–480. [Google Scholar] [CrossRef]
- Matsuura, K.; Himuro, C.; Yokoi, T.; Yamamoto, Y.; Vargo, E.L.; Keller, L. Identification of a pheromone regulating caste differentiation in termites. Proc. Natl. Acad. Sci. USA 2010, 107, 12963–12968. [Google Scholar] [CrossRef]
- Smith, A.A.; Millar, J.G.; Suarez, A.V. Comparative analysis of fertility signals and sex-specific cuticular chemical profiles of Odontomachus trap-jaw ants. J. Exp. Biol. 2016, 219, 419–430. [Google Scholar] [CrossRef]
- Steitz, I.; Ayasse, M. Macrocyclic lactones act as a queen pheromone in a primitively eusocial sweat bee. Curr. Biol. under review.
- Steitz, I.; Kingwell, C.; Paxton, R.J.; Ayasse, M. Evolution of caste-specific chemical profiles in halictid bees. J. Chem. Ecol. 2018, 44, 827–837. [Google Scholar] [CrossRef]
- Johansson, I.; Svensson, B.G.; Tengö, J.; Bergström, G. Systematic relationship of halictinae bees based on the pattern of macrocyclic lactones in the Dufour gland secretion. Insect Biochem. 1982, 12, 161–170. [Google Scholar] [CrossRef]
- Cane, J.H. Dufour’s gland secretion in the cell linings of bees (Hymenoptera: Apoidea). J. Chem. Ecol. 1981, 7, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Duffield, R.M.; Fernandes, A.; Lamb, C.; Wheeler, J.W.; Eickwort, G.C. Macrocyclic lactones and isopentenyl esters in the Dufour’s gland secretion of halictine bees (Hymenoptera: Halictidae). J. Chem. Ecol. 1981, 7, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Hefetz, A. The role of Dufour’s gland secretions in bees. Physiol. Entomol. 1987, 12, 243–253. [Google Scholar] [CrossRef]
- Hefetz, A.; Bergström, G.; Tengö, J. Species, individual and kin specific blends in Dufour’s gland secretions of halictine bees. J. Chem. Ecol. 1986, 12, 197–208. [Google Scholar] [CrossRef]
- Smith, B.H.; Carlson, R.G.; Frazier, J. Identification and bioassay of macrocyclic lactone sex pheromone of the halictine bee Lasioglossum zephyrum. J. Chem. Ecol. 1985, 11, 1447–1456. [Google Scholar] [CrossRef]
- Ayasse, M.; Engels, W.; Lübke, G.; Taghizadeh, T.; Francke, W. Mating expenditures reduced via female sex pheromone modulation in the primitively eusocial halictine bee, Lasioglossum (Evylaeus) malachurum (Hymenoptera: Halictidae). Behav. Ecol. Sociobiol. 1999, 45, 95–106. [Google Scholar] [CrossRef]
- Ayasse, M.; Paxton, R.J.; Tengö, J. Mating behavior and chemical communication in the order Hymenoptera. Annu. Rev. Entomol. 2001, 46, 31–78. [Google Scholar] [CrossRef]
- Wcislo, W.T. The role of learning in the mating biology of a sweat bee Lasioglossum zephyrum (Hymenoptera: Halictidae). Behav. Ecol. Sociobiol. 1987, 20, 179–185. [Google Scholar] [CrossRef]
- Smith, B.; Weller, C. Social competition among gynes in halictine bees: The influence of bee size and pheromones on behavior. J. Insect Behav. 1989, 2, 397–411. [Google Scholar] [CrossRef]
- Keller, L.; Nonacs, P. The role of queen pheromones in social insects: Queen control or queen signal? Anim. Behav. 1993, 45, 787–794. [Google Scholar] [CrossRef]
- Babu, M.J.; Ankolekar, S.M.; Rajashekhar, K.P. Castes of the weaver ant Oecophylla smaragdina (Fabricius) differ in the organization of sensilla in their antennae and mouthparts. Curr. Sci. 2011, 101, 755–764. [Google Scholar]
- Fialho, M.C.Q.; Guss-Matiello, C.P.; Zanuncio, J.C.; de Oliveira Campos, L.A.; Serrao, J.E. A comparative study of the antennal sensilla in corbiculate bees. J. Apicult. Res. 2014, 53, 392–403. [Google Scholar] [CrossRef]
- Ravaiano, S.V.; de Paiva Ferreira, R.; de Oliveira Campos, L.A.; Martins, G.F. The antennal sensilla of Melipona quadrifasciata (Hymenoptera: Apidae: Meliponini): A study of different sexes and castes. Naturwissenschaften 2014, 101, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Leonhardt, S.D.; Menzel, F.; Nehring, V.; Schmitt, T. Ecology and evolution of communication in social insects. Cell 2016, 164, 1277–1287. [Google Scholar] [CrossRef]
- Smith, A.A.; Liebig, J. The evolution of cuticular fertility signals in eusocial insects. Curr. Opin. Insect Sci. 2017, 22, 79–84. [Google Scholar] [CrossRef]
- Villalta, I.; Abril, S.; Cerdá, X.; Boulay, R. Queen control or queen signal in ants: What remains of the controversy 25 years after Keller and Nonacs’ seminal paper? J. Chem. Ecol. 2018, 44, 805–817. [Google Scholar] [CrossRef]
- Westrich, P. Die Wildbienen Baden-Württembergs; Verlag Eugen Ulmer: Stuttgart, Germany, 1989. [Google Scholar]
- Hölldobler, B.; Michener, C.D. Mechanisms of identification and discrimination in social Hymenoptera. In Evolution of Social Behavior: Hypotheses and Empirical Tests; Markl, H., Ed.; Verlag Chemie: Weinheim, Germany, 1980. [Google Scholar]
- Soro, A.; Ayasse, M.; Zobel, M.U.; Paxton, R.J. Complex sociogenetic organization and the origin of unrelated workers in a eusocial sweat bee, Lasioglossum malachurum. Insect Soc. 2009, 56, 55–63. [Google Scholar] [CrossRef]
- Duchateau, M.J.; Velthuis, H.H.W. Ovarian development and egg laying in workers of Bombus terrestris. Entomol. Exp. Appl. 1989, 51, 199–213. [Google Scholar] [CrossRef]
- Dani, F.R.; Jones, G.R.; Corsi, S.; Beard, R.; Pradella, D.; Turillazzi, S. Nestmate recognition cues in the honey bee: Differential importance of cuticular alkanes and alkenes. Chem. Senses 2005, 30, 477–489. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER V6: User Manual-Tutorial; Plymouth Marine Laboratory: Plymouth, UK, 2006. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Ayasse, M.; Engels, W.; Hefetz, A.; Tengö, J.; Lübke, G.; Francke, W. Ontogenetic patterns of volatiles identified in Dufour’s gland extracts from queens and workers of the primitively eusocial halictine bee, Lasioglossum malachurum (Hymenoptera: Halictidae). Insect Soc. 1993, 40, 41–58. [Google Scholar] [CrossRef]
- Soro, A.; Ayasse, M.; Zobel, M.U.; Paxton, R.J. Kin discriminators in the eusocial sweat bee Lasioglossum malachurum: The reliability of cuticular and Dufour’s gland odours. Behav. Ecol. Sociobiol. 2011, 65, 641–653. [Google Scholar] [CrossRef]
- Blomquist, G.J.; Bagnères, A.G. Insect Hydrocarbons: Biology, Biochemistry and Chemical Ecology; Cambridge University Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Sprenger, P.P.; Burkert, L.H.; Abou, B.; Federle, W.; Menzel, F. Coping with the climate: Cuticular hydrocarbon acclimation of ants under constant and fluctuating conditions. J. Exp. Biol. 2018, 221, jeb171488. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, A.G.; Pomonis, J.G. Physical properties of insect cuticular hydrocarbons: The effects of chain length, methyl-branching and unsaturation. Comp. Biochem. Physiol. 1995, 112, 243–249. [Google Scholar] [CrossRef]
- Chapuisat, M. Smells like queen since the cretaceous. Science 2014, 343, 254–255. [Google Scholar] [CrossRef] [PubMed]
- Amsalem, E.; Orlova, M.; Grozinger, C.M. A conserved class of queen pheromones? Re-evaluating the evidence in bumblebees (Bombus impatiens). Proc. R. Soc. B Biol. Sci. 2015, 282, 20151800. [Google Scholar] [CrossRef] [PubMed]
- Amsalem, E.; Padilla, M.; Schreiber, P.M.; Altman, N.S.; Hefetz, A.; Grozinger, C.M. Do bumble bee, Bombus impatiens, queens signal their reproductive and mating status to their workers? J. Chem. Ecol. 2017, 43, 563–572. [Google Scholar] [CrossRef]
- Oi, C.A.; Oliveira, R.C.; van Zweden, J.S.; Mateus, S.; Millar, J.G.; Nascimento, F.S.; Wenseleers, T. Do primitively eusocial wasps use queen pheromones to regulate reproduction? A case study of the paper wasp Polistes satan. Front. Ecol. Evol. 2019, 7, 199. [Google Scholar] [CrossRef]
- Padilla, M.; Amsalem, E.; Altman, N.; Hefetz, A.; Grozinger, C.M. Chemical communication is not sufficient to explain reproductive inhibition in the bumblebee Bombus impatiens. R. Soc. Open Sci. 2016, 3, 160576. [Google Scholar] [CrossRef]
- Pardi, L. Dominance order in Polistes wasps. Physiol. Zool. 1948, 21, 1–13. [Google Scholar] [CrossRef]
- Starkey, J.; Brown, A.; Amsalem, E. The road to sociality: Brood regulation of worker reproduction in the simple eusocial bee Bombus impatiens. Anim. Behav. 2019, 154, 57–65. [Google Scholar] [CrossRef]
- Buckle, G.R. Queen-worker behavior and nestmate interactions in young colonies of Lasioglossum zephyrum. Insect Soc. 1982, 29, 125–137. [Google Scholar] [CrossRef]
- Brothers, D.J.; Michener, C.D. Interactions in colonies of primitively social bees. J. Comp. Physiol. 1974, 90, 129–168. [Google Scholar] [CrossRef]
- Groh, C.; Rössler, W. Caste-specific postembryonic development of primary and secondary olfactory centers in the female honeybee brain. Arthropod Struct. Dev. 2008, 37, 459–468. [Google Scholar] [CrossRef]
- Oliveira, R.C.; Oi, C.A.; do Nascimento, M.M.C.; Vollet-Neto, A.; Alves, D.A.; Campos, M.C.; Nascimento, F.; Wenseleers, T. The origin and evolution of queen and fertility signals in Corbiculate bees. BMC Evol. Biol. 2015, 15, 254. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavior | Description | Mode of Interaction |
|---|---|---|
| backing | fast movement backwards apart from the encountering bee | avoidance/submission |
| moving backwards | slow movement backwards apart from the encountering bee | avoidance |
| nudging | fast movement forwards, bee brings its face into contact with face of encountering bee | dominance |
| passing | both encountering bees pass and move on in opposite directions | cooperation |
| withdraw | 180 degree turn away from encountering bee | avoidance |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steitz, I.; Brandt, K.; Biefel, F.; Minat, Ä.; Ayasse, M. Queen Recognition Signals in Two Primitively Eusocial Halictid Bees: Evolutionary Conservation and Caste-Specific Perception. Insects 2019, 10, 416. https://doi.org/10.3390/insects10120416
Steitz I, Brandt K, Biefel F, Minat Ä, Ayasse M. Queen Recognition Signals in Two Primitively Eusocial Halictid Bees: Evolutionary Conservation and Caste-Specific Perception. Insects. 2019; 10(12):416. https://doi.org/10.3390/insects10120416
Chicago/Turabian StyleSteitz, Iris, Katharina Brandt, Felix Biefel, Ädem Minat, and Manfred Ayasse. 2019. "Queen Recognition Signals in Two Primitively Eusocial Halictid Bees: Evolutionary Conservation and Caste-Specific Perception" Insects 10, no. 12: 416. https://doi.org/10.3390/insects10120416
APA StyleSteitz, I., Brandt, K., Biefel, F., Minat, Ä., & Ayasse, M. (2019). Queen Recognition Signals in Two Primitively Eusocial Halictid Bees: Evolutionary Conservation and Caste-Specific Perception. Insects, 10(12), 416. https://doi.org/10.3390/insects10120416