Lethal Effects of High Temperatures on Brown Marmorated Stink Bug Adults before and after Overwintering


Department of Agronomy Food Natural Resources Animals and Environment, University of Padova, 35020 Legnaro, Padova, Italy
*
Author to whom correspondence should be addressed.
Insects 2019, 10(10), 355; https://doi.org/10.3390/insects10100355
Submission received: 29 August 2019
/
Revised: 11 October 2019
/
Accepted: 15 October 2019
/
Published: 18 October 2019
Abstract
:The invasive brown marmorated stink bug, Halyomorpha halys, is causing economic and ecological damage in invaded areas. Its overwintering behavior warrants mitigation practices in warehouses and shipping operations. The aim of this study was to characterize the mortality response curves of H. halys adults to short high-temperature exposure. Here we compared field-collected individuals entering (ENA) and exiting diapause (EXA). EXA adults displayed increased susceptibility to high temperatures compared to ENA individuals. Complete mortality of all tested individuals was obtained after 10 min exposure at 50.0 °C, and after 15 (EXA) or 20 min (ENA) at 47.5 °C. The nutritional status of these insects had no effect on high-temperature tolerance. The mortality curves obtained here may be used for the definition of cost-effective heat treatments aimed at the H. halys control.
1. Introduction
Invasive insect species cause significant economic and ecological damage in newly-invaded areas [1,2], resulting in high economic costs [3,4]. In the recent past, the brown marmorated stink bug, Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) started to spread worldwide from its native range in East Asia. It is an invasive phytophagous pest, already recorded to feed on more than 170 plant species worldwide [5]. In Europe, the first H. halys was recorded outside its native range in the 2000s in Switzerland [6], and it is now widespread throughout many European countries [5,7,8]. In Italy, H. halys rapidly became an agricultural pest, resulting in damage to crops and causing nuisance problems for the human population [9,10,11].
In autumn, adult H. halys field populations invade buildings searching for shelters in which to overwinter [12,13]. As a result, adults can be found in warehouses and in food industries that are present in the area of invasion, with potential risks of the contamination of goods that could be distributed worldwide [14,15]. This aspect plays a key role in the invasion process of this pest. Overwintering aggregations are frequently intercepted in freightliners transporting cargo, and this is likely to represent an important invasion pathway for H. halys worldwide [7,16,17].
In disinfestation programs, insect control in food industries can be achieved by modifying the environmental temperature below or above insects’ critical temperature levels. The use of extreme temperatures can be a viable method in quarantine security in order to avoid the introduction of exotic pests into other countries. Treatments using suboptimal low temperatures (i.e., freezing) are less commonly used due to the high energetic costs, while high temperatures have been extensively used since the last century [18,19,20,21] to kill insect pests of stored products [22,23]. Furthermore, heat treatments have been proposed for the control of other insects infesting commodities and against structural pests, where killing temperatures can be applied in multiple forms, such as hot water, vapor heat, forced hot air, dry heat, or other methodologies [24,25,26].
Acute changes in temperature result in significant metabolic changes in insects. These changes affect many physiological processes, including anaerobiotic metabolism, enzyme activity, altered cellular membranes, behavior, and nervous and endocrine systems. External structures such as the cuticular wax of the insect can also be modified [27]. Above 40.0 °C these effects become more critical [27]. Generally, short exposures up to temperatures of 50.0 °C are lethal for almost all tested insects [28]. Similar high-temperature treatments are used for quarantine treatments of goods [29], where exposures of 50–60 °C for 24 h are required in disinfestation programs [23].
For H. halys, it is known that consistently low temperature is required to kill adults [30,31,32], making cold treatments a less suitable practice in disinfestation programs. Conversely, heat treatments have been proposed to manage H. halys in closed environments by exposing goods possibly containing insects to 50.0 °C for 15 min or more [33]. However, more detailed information is necessary to optimize heat treatments, in order to reduce energetic costs and increase efficacy in H. halys control.
Additionally, treatments aimed at managing “hitchhiking” pest populations within shipped goods may involve H. halys in different physiological states. Halyomorpha halys can overwinter as aggregations of adults in non-feeding and non-reproductive state within hidden and protected sites [34,35]. Such insects typically display gradual springtime emergence and field dispersion patterns. This transition from overwintering to the overwintered state is associated with non-reproductive to reproductive physiological changes [36,37]. When overwintering, the nutritional levels of H. halys populations decline [38] but then display a gradual increase as the winter progresses through the summer [39]. Different physiological and nutritional statuses can have a potential effect on resistance to high temperatures, however this information is not available for H. halys since previous research was performed using laboratory-maintained colonies [33].
The aim of this study was to characterize short-term (2.5 min to 1 h) high-temperature mortality response curves of H. halys adults and to test if the effect of heat exposure changes between adults entering and exiting diapause. Lastly, we tested whether the nutritional status of H. halys influenced their mortality induced by heat exposure.
2. Materials and Methods
2.1. Insects
Both entering diapause (September to October; hereafter ENA) and exiting diapause adults (April to May; hereafter EXA) were hand-collected in Legnaro, Italy (45.344872 N, 11.956208 E) in 2017. ENA adults were insects that developed in the current year (two generations per year are recorded in northern Italy [40]). EXA adults were collected from artificial overwintering units placed in outdoor conditions under shade and covered to be protected from weather events such as rainfall. Artificial overwintering units consisted of five plastic boxes (50 × 35 × 15 cm3; IKEA®, Delft, The Netherlands) containing wooden cages (34 × 19 × 10 cm3) with a 34 × 1 cm2 slit along one side. Cardboard and folded paper (abt. 20 × 20 cm2) were placed inside wooden cages to provide shelter for insects. In fall 2016, 250 H. halys adults were placed in each box. These units were monitored three times per week from November 2016 to May 2017. Adults were considered to be exiting diapause when they were found outside the wooden cages in the plastic boxes. Diapause was confirmed by insect dissection, where no eggs were recorded in ENA or EXA females which were in the “one immature oocyte per ovariole” rank as described by Nielsen et al. [37].
2.2. Lethal High Temperatures with Short Exposure Times
Laboratory experiments were performed using a thermocryostat (LAUDA Alpha, RA 12®, LAUDA DR. R. WOBSER GMBH & CO. KG, Lauda-Königshofen, Germany) for heat treatment. Halyomorpha halys adults were placed singly in glass vials (7-mL volume) and were sealed by a cotton swab to allow gas exchange. The available air volume of vials for each adult was about 5.5 mL. The starting relative humidity was 50(± 2)%, and fluctuated to 56(± 4)% during the experiment. At least 30 adults per temperature-time combination were tested, in replicates of 10 vials. Before treatment, adults were collected from buildings (ENA) or from overwintering units (EXA) and kept for two hours in boxes at room temperature (23 °C). Vials were placed in the thermocryostat and exposed to temperatures ranging from 32.5 to 60.0 °C, with a 2.5 °C step (i.e., 12 temperature levels: 32.5, 35.0, 37.5, 40.0, 42.5, 45.0, 47.5, 50.0, 52.5, 55.0, 57.5, or 60.0 °C, herein referred to as set temperatures), for six different time periods (i.e., 2.5, 5, 10, 15, 30, or 60 min) for both ENA and EXA adults. The following six temperatures were also performed only for the ENA adults: 41.5, 45.5, 46.0, 46.5, 47.0, and 56.5 °C. The temperature within the vials was checked using a thermocouple (RhOS, 4-Channel Digital Thermometer Thermocouple Sensor). Heating rate was 15.0 °C min−1, with heating times ranging from 0.6 to 2.5 min depending on the set temperature. When the set temperature was reached, insects were transferred into vials and then placed in the thermocryostat. Vials were kept in the thermocryostat for the time required (i.e., 2.5, 5, 10, 15, 30 or 60 min) according to the experimental design. Then, the insects were removed from vials, kept in rearing cages (30 × 30 × 30 cm3; BugDorm-1, MegaView Science Co., Ltd., Taiwan) at room temperature and followed for 24 h to assess the mortality. Control ENA and EXA insects were subjected to room temperature (maintained at 23 °C during all the procedures).
2.3. Nutrient Index and H. halys Mortality after High-Temperature Exposures
Prior to heat exposure, we evaluated the weight and nutritional level of each individual calculating the nutrient index (NI) as described by Funayama [41] and Skillman et al. (i.e., weight of insect (mg)/prothorax width (mm)3 [38,39]). The nutrient index is a non-destructive and easy-to-use method that was used as an indicator of the nutrient status of H. halys [41]. We tested if the weight or level of the nutrient index had an effect on adult mortality after heat exposure. Another set of 240 ENA and EXA females and males were exposed to two of the temperatures reported before for 15, 30, or 60 min using the procedures described above. After heat treatment, insects were processed following the same procedure described above.
2.4. Data Analysis
Data on the effect of lethal high temperatures were analyzed with a probit regression using the PROBIT procedure of SAS® (ver. 9.4) [42] and interpolating the observed data to mortality curves. Lethal temperatures for 50% (LT50) and 99% (LT99) mortality of adults, for any exposure time, were determined. We ran probit regression on data obtained with 2.5, 15, 30, and 60 min exposure times. Comparisons between the two groups of adults (i.e., EXA and ENA) for the lethal temperature levels (i.e., LT50 and LT99) were done by using the lethal dose ratios method (α = 0.05), which is based on the 95% confidence limits of lethal temperature levels, depending on the intercepts and the slopes of the probit lines and considering the variance-covariance matrices as described by Robertson et al. [43]. The method is used for comparisons of point estimates in different insect mortality curves obtained from treatment exposures [43].
The time causing mortality (MT) was studied through the probit regression analysis for 47.5 and 50.0 °C using all exposure times. The probit analysis was conducted as stated before. In this case, the exposure time was considered as the independent variable and the mortality as the dependent one. Comparisons between the two groups of adults (i.e., EXA and ENA) for the lethal time levels MT50 and MT99 were done using the lethal dose ratios method (α = 0.05) [43].
Finally, data on nutrient index and insect weight were analyzed separately using a general linear mixed model, with the MIXED procedure of SAS® (ver. 9.4) [42] with an F-test (α = 0.05) followed by a Tukey–Kramer test (α = 0.05) to determine if there were differences in nutrient index or weight in dead and alive insects after exposure to 42.5 or 45.0 °C for 15, 30, or 60 min. In this analysis, we considered the status of the insect (i.e., dead or alive), sex, state of diapause (i.e., EXA or ENA), temperature (i.e., 42.5 or 45.0 °C), and their interactions as sources of variation. Data on mortality after 15, 30, or 60 min were analyzed separately. Data were checked for model assumptions prior to the analysis, and untransformed data were used.
3. Results
3.1. Lethal High Temperatures
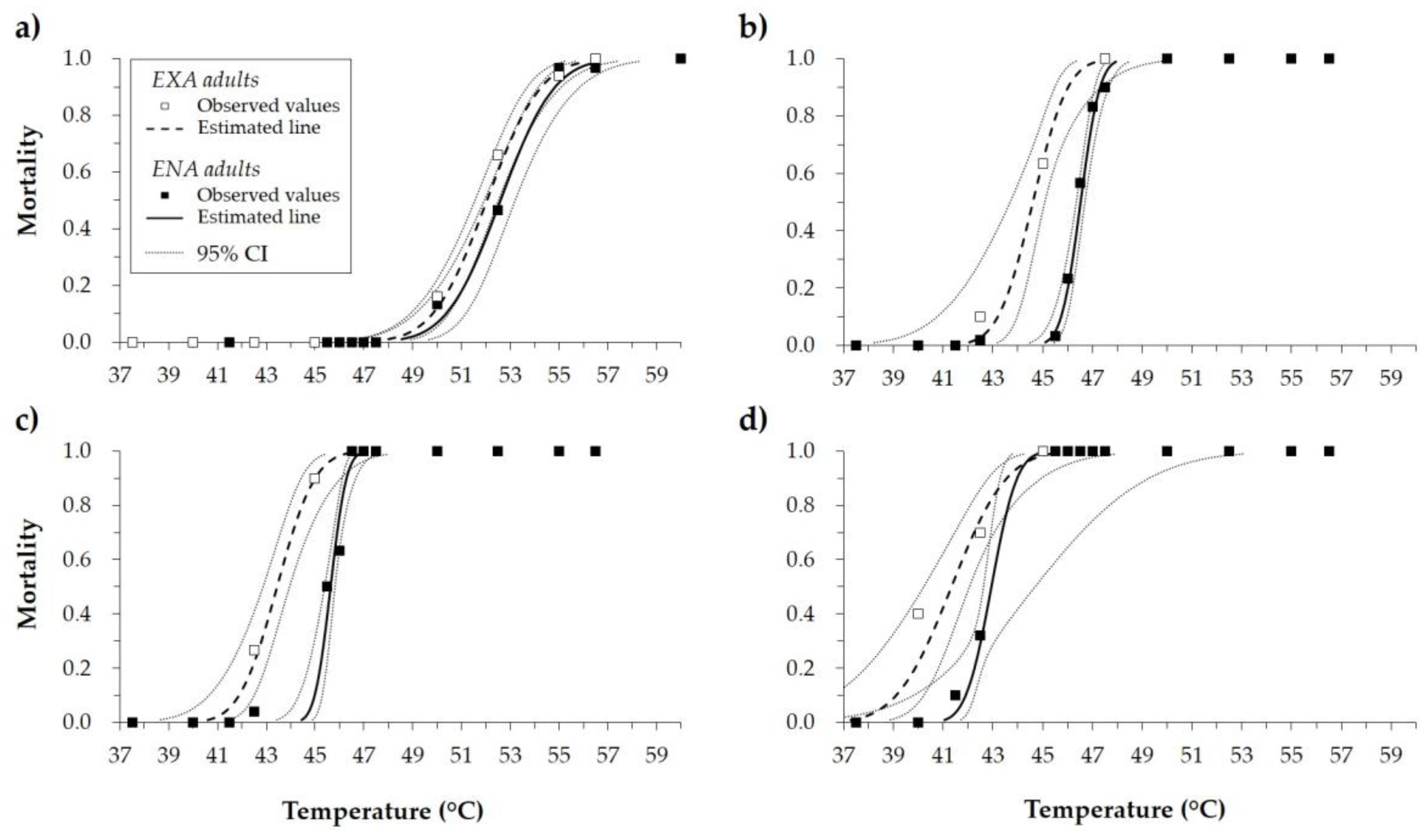
An increase in insect mortality was directly correlated with an increase in temperature. Shorter exposure times required higher temperatures to result in comparable H. halys mortality levels. With 2.5 min exposure, the minimum temperature to induce H. halys mortality was 50.0 °C for both EXA and ENA adults (Figure 1a). With exposure times of 15 and 30 min, the minimum temperature inducing mortality was 42.5 °C (Figure 1b,c), while after 60 min of exposure the minimum temperature to cause mortality was 40.0 °C or 41.5 °C for EXA and ENA adults, respectively (Figure 1d).
Considering all the exposure times tested, LT50 ranged from 41.3 to 52.6 °C and LT99 from 44.8 to 56.7 °C. Mortality curves were different between EXA and ENA adults. At the same exposure time, LTs were lower for EXA adults, which were more susceptible to high temperatures than ENA adults (Table 1).
3.2. Exposure Times at High Temperatures
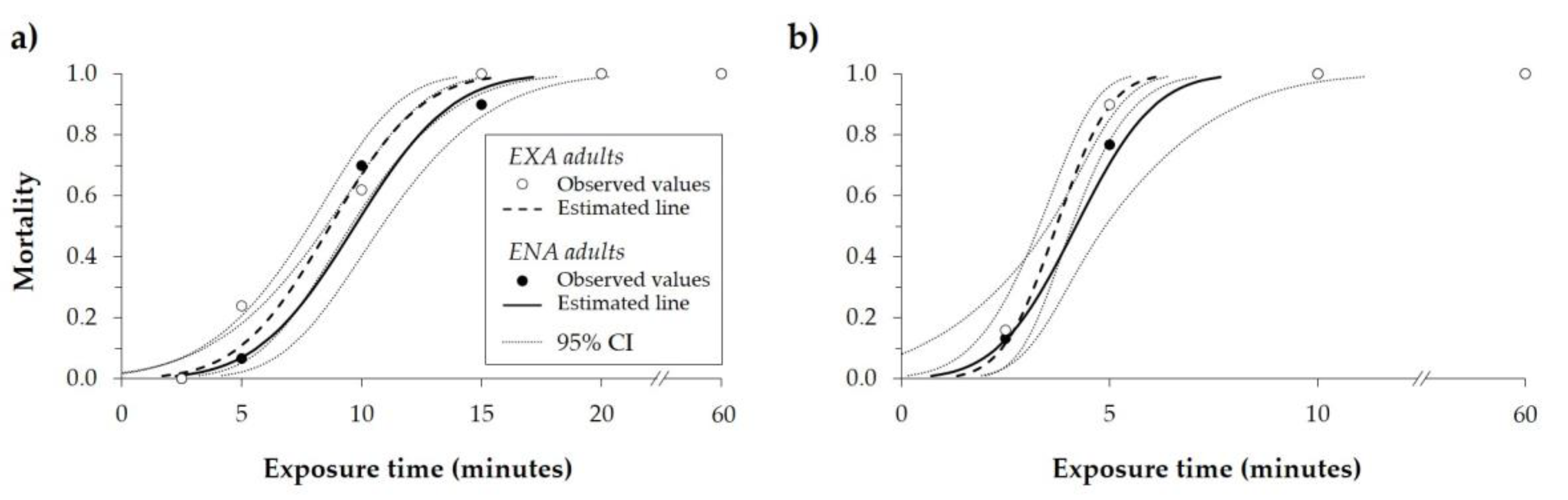
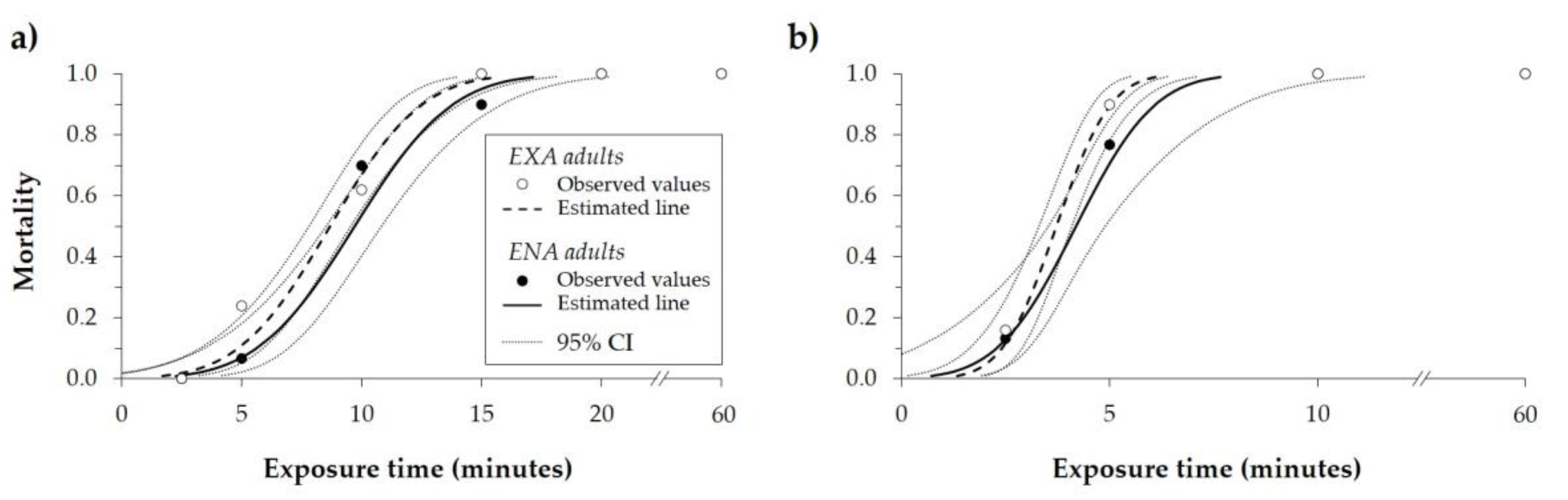
The lowest time inducing H. halys mortality was 2.5 min when adults were exposed to 50.0 °C, and 5 min when exposed to 47.5 °C. The mortality rate was 100% after 10 min of exposure at 50.0 °C, and after 15 (EXA) or 20 min (ENA) at 47.5 °C. No survival was recorded for adults exposed to these two temperatures for 20 min or more (Figure 2). At 47.5 °C, the time required to kill 50% of the adults (MT50) was 8.67 and 9.69 min, while the MT99 was 15.65 and 17.14 min for EXA and ENA, respectively (Table 2). At 50 °C, MT50 was 3.72 and 4.19 min, while the MT99 was 6.10 and 7.67 min for EXA and ENA, respectively (Table 2). For the exposure to the same temperature, the lethal dose ratios method did not show differences between the EXA and ENA curves for the MT50, while MT99 was higher for ENA than EXA at both tested temperatures (Table 2).
3.3. Weight and Nutrient Index
The mortality rate of H. halys after exposure to 42.5 and 45.0 °C for 15, 30, and 60 min was not different considering insect weight or nutrient index (Table 3 and Table 4). Weight (Table 3) and nutrient index values (Table 4) were higher for ENA (weight: 137.36 ± 36.99 mg; NI: 0.24 ± 0.03) than for EXA (weight: 115.27 ± 26.76 mg; NI: 0.20 ± 0.03) adults, and the weight was higher for females (147.83 ± 32.35 mg) than for males (105.91 ± 20.38 mg; Table 3 and Table 4).
4. Discussion
The mortality parameters from different time and temperature combinations provide significant new insights into effective heat treatments aimed at H. halys control. These data deliver scenarios where high levels of H. halys mortality were obtained using relatively limited energetic costs. Such heat treatments can be effectively used as part of quarantine procedures for H. halys contamination. Both ENA and EXA adults are likely H. halys life stages that could be effectively targeted by such heat treatments. These insects are in a non-reproductive state [36,37], as confirmed by the absence of eggs in ENA and EXA females in our study. ENA adults enter into overwintering sites in autumn, while EXA adults emerge from these sites in spring [40]. In general, these H. halys adults do not have to cope with high-temperature stress because such high temperatures do not normally occur in autumn or spring. On the other hand, adults developed during the warmer period of the year (which move to overwintering sites at the end of the summer [12,13]) may have some forms of tolerance to high temperatures. This topic fell outside the scope of this study and should be investigated in future.
Halyomorpha halys adults start to emerge from overwintering sites when temperatures exceed 10.0 °C, and flight activity strongly increases at temperatures above 15.0 °C [44,45,46]. In shipping cargo, temperature up to 30 °C could stimulate the activity of overwintering H. halys, but these insects will die due to the lack of available food [17]. Aigner and Kuhar [33] found that a minimum temperature of 45.0 °C for 15 min or 35.0 °C for 4 h were required to kill H. halys adults in the laboratory, but a precise minimum threshold could not be derived from their data. Additionally, they used H. halys from an artificial mass rearing colony and did not assess potential effects of adult diapause state. In the present manuscript we observed that mortality of H. halys started at 40.0 °C and 41.5 °C for 60 min for EXA and ENA adults, respectively. Using data from probit regression we determined a minimum threshold for H. halys mortality that was 41.1 °C for ENA adults and 37.3 °C for EXA adults considering a 60 min exposure, while higher thresholds were obtained with 2.5 min of exposure (i.e., 48.6 °C and 48.0 °C for ENA and EXA adults, respectively).
Notably, our results showed that EXA adults were more sensitive to high temperatures than ENA ones, possibly due to the physiological status of the insect, as adults entering diapause have a reduction of nutritional levels and a different energetic fitness than adults exiting diapause [38,39]. EXA adults had a lower weight and nutrient index than ENA adults. However, the lipid, glycogen, and sugar content do not seem to be related to the nutrient index of H. halys adults [38]. In our study, the state of diapause well explained the mortality levels showed by H. halys adults, while the nutrient index or the weight did not, confirming that the adult’s state of diapause more than their nutrient index should be considered during the optimization of heat treatment against H. halys.
Furthermore, all the tested H. halys adults died with exposures to at least 48.0 °C for 15 min, or 45.5 °C for 1 h. In the USA, H. halys high-temperature mortality showed no adult survival with the exposure to 50.0 °C for at least 15 min, or to 45.0 °C for 1 h or more [33]. Here we found that the time required to kill 99% of the adults was ~17 or ~7.5 min for 47.5 or 50 °C, respectively (considering ENA adults as the most conservative case), highlighting the temperature–time combinations that may be considered for practical uses for heat treatment at quarantine facilities. Differences on mortality detected between studies may be related to the use of laboratory-reared insects (as in Aigner and Kuhar [33]) versus field-collected ones (as in this study), but also by genetic features of the tested insects [47] or the methodology performed [48,49,50].
The results of the present study, and in particular data on minimum thresholds for H. halys mortality obtained with 1-h exposure, can also be of importance in forecasting the geographical distribution of this invasive pest. Previous published data on H. halys development also showed that European populations failed to develop under controlled conditions at 35.0 °C or more [51], implying possible restrictions in the distribution area of the species and influencing its phenology in areas where unsuitable climate conditions and heatwaves occur [52]. Heatwaves, which are extreme short-term climatic events defined as prolonged periods of excessive heat, are globally increasing in frequency [53,54]. Typically, during heatwaves in several parts of the word, maximum daily temperatures can exceed 35–40.0 °C for several days [55,56,57]. This temperature range poses risks for the survival of H. halys in some of these areas (e.g., southern Europe and Australia) where heatwaves can be associated with temperatures above 35.0 °C for hours. Here we found that temperatures higher than 37.3 °C for 1 h could reduce the survival of H. halys adults. This type of information can be used to update current models on the dynamics and geographic distribution of this pest [33,58,59,60] accounting for the effect of high environmental temperatures on mortality. Data provided here are not complete for understanding the impact of high environmental temperature on H. halys, but may represent a starting point for future research that should investigate the effect of high temperatures on young stages and simulating longer exposure time.
5. Conclusions
The results obtained here underline the ability of H. halys to tolerate high temperatures. These data provide important parameters that can be used as heat shock treatments for H. halys control in quarantine methods for the disinfestation of goods and in fresh food export industries. Heat treatments with short-time exposure should be targeted on the H. halys physiological status, which seems to be related to the overwintering state. The definition of cost-effective heat treatments aimed at the H. halys control may also be performed following the mortality curves reported here.
Author Contributions
Conceptualization, C.D. and A.P.; methodology, D.S., C.D. and A.P.; formal analysis, D.S. and A.P.; investigation, D.S.; writing—original draft preparation, D.S. and A.P.; writing—review and editing, D.S., C.D. and A.P.
Funding
This research was partially supported by Bottega s.p.a. (Godega di S. Urbano, Italy).
Acknowledgments
The authors thank Vaughn Walton (Oregon State University, USA) for comments on an early version of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kenis, M.; Auger-Rozenberg, M.-A.; Roques, A.; Timms, L.; Péré, C.; Cock, M.J.W.; Settele, J.; Augustin, S.; Lopez-Vaamonde, C. Ecological effects of invasive alien insects. Biol. Invasions 2009, 11, 21–45. [Google Scholar] [CrossRef]
- Kenis, M.; Branco, M. Impact of alien terrestrial arthropods in Europe. Chapter 5. BioRisk 2010, 4, 51–71. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, D.; Zuniga, R.; Morrison, D. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Kenis, M.; Roques, A.; Santini, A.; Liebhold, A.M. Impact of non-native invertebrates and pathogens on market forest tree resources. In Impact of Biological Invasions on Ecosystem Services; Vilà, M., Hulme, P.E., Eds.; Invading Nature—Springer Series in Invasion Ecology; Springer: Cham, Switzerland, 2017; Volume 12, pp. 103–117. ISBN 9783319451213. [Google Scholar]
- Leskey, T.C.; Nielsen, A.L. Impact of the invasive brown marmorated stink bug in North America and Europe: History, biology, ecology, and management. Annu. Rev. Entomol. 2018, 63, 599–618. [Google Scholar] [CrossRef]
- Wermelinger, B.; Wyniger, D.; Forster, B. First records of an invasive bug in Europe: Halyomorpha halys Stål (Heteroptera: Pentatomidae), a new pest on woody ornamentals and fruit trees? Mitteilungen Schweiz. Entomol. Ges. 2008, 81, 1–8. [Google Scholar]
- Haye, T.; Gariepy, T.; Hoelmer, K.; Rossi, J.-P.; Streito, J.-C.; Tassus, X.; Desneux, N. Range expansion of the invasive brown marmorated stinkbug, Halyomorpha halys: An increasing threat to field, fruit and vegetable crops worldwide. J. Pest Sci. 2015, 88, 665–673. [Google Scholar] [CrossRef]
- Vétek, G.; Károlyi, B.; Mészáros, Á.; Horváth, D.; Korányi, D. The invasive brown marmorated stink bug (Halyomorpha halys) is now widespread in Hungary. Entomol. Gen. 2018, 38, 3–14. [Google Scholar] [CrossRef]
- Maistrello, L.; Dioli, P.; Bariselli, M. Trovata una cimice esotica dannosa per i frutteti. Agricoltura 2013, 6, 67–68. [Google Scholar]
- Maistrello, L.; Dioli, P.; Bariselli, M.; Mazzoli, G.L.; Giacalone-Forini, I. Citizen science and early detection of invasive species: Phenology of first occurrences of Halyomorpha halys in Southern Europe. Biol. Invasions 2016, 18, 3109–3116. [Google Scholar] [CrossRef]
- Bariselli, M.; Bugiana, R.; Maistrello, L. Distribution and damage caused by Halyomorpha halys in Italy. EPPO Bull. 2016, 46, 332–334. [Google Scholar] [CrossRef]
- Inkley, D.B. Characteristics of home invasion by the brown marmorated stink bug (Hemiptera: Pentatomidae). J. Entomol. Sci. 2012, 47, 125–130. [Google Scholar] [CrossRef]
- Cambridge, J.; Payenski, A.; Hamilton, G.C. The distribution of overwintering brown marmorated stink bugs (Hemiptera: Penatatomidae) in college dormitories. Fla. Entomol. 2015, 98, 1257–1259. [Google Scholar] [CrossRef]
- Nicoli Aldini, R.; Fraschini, A. Gestione della difesa antiparassitaria in un’industria conserviera del nord Italia. In Programma e Riassunti, Proceedings of the X Simposio—La Difesa Antiparassitaria nelle Industrie Alimentari e la Protezione degli Alimenti, Piacenza, Italy, 20–22 September 2017; Accademia Nazionale Italiana di Entomologia: Piacenza, Italy, 2017; p. 47. [Google Scholar]
- Acebes-Doria, A.L.; Morrison, W.R.; Short, B.D.; Rice, K.B.; Bush, H.G.; Kuhar, T.P.; Duthie, C.; Leskey, T.C. Monitoring and biosurveillance tools for the brown marmorated stink bug, Halyomorpha halys (Stål) (Hemiptera: Pentatomidae). Insects 2018, 9, 82. [Google Scholar] [CrossRef] [PubMed]
- Nixon, L.J.; Morrison, W.R.; Rice, K.B.; Brockerhoff, E.G.; Leskey, T.C.; Guzman, F.; Khrimian, A.; Goldson, S.; Rostás, M. Identification of volatiles released by diapausing brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae). PLoS ONE 2018, 13, e0191223. [Google Scholar] [CrossRef] [PubMed]
- Nixon, L.J.; Tabb, A.; Morrison, W.R.; Rice, K.B.; Brockerhoff, E.G.; Leskey, T.C.; van Koten, C.; Goldson, S.; Rostás, M. Volatile release, mobility, and mortality of diapausing Halyomorpha halys during simulated shipping movements and temperature changes. J. Pest Sci. 2019, 92, 633–641. [Google Scholar] [CrossRef]
- Burges, H.D.; Burrell, N.J. Cooling bulk grain in the British climate to control storage insects and to improve keeping quality. J. Sci. Food Agric. 1964, 15, 32–50. [Google Scholar] [CrossRef]
- Chapman, R.N. Insects infesting stored food products. Minn. Bull. 1932, 198, 1–83. [Google Scholar]
- Dean, G.A. Heat as a means of controlling mill insects. J. Econ. Entomol. 1911, 4, 142–161. [Google Scholar] [CrossRef]
- Dean, G.A. Further data on heat as a means of controlling mill insects. J. Econ. Entomol. 1913, 6, 40–55. [Google Scholar] [CrossRef]
- Fields, P.G.; White, N.D.G. Alternatives to methyl bromide treatments for stored-product and quarantine insects. Annu. Rev. Entomol. 2002, 47, 331–359. [Google Scholar] [CrossRef]
- Phillips, T.W.; Throne, J.E. Biorational approaches to managing stored-product insects. Annu. Rev. Entomol. 2010, 55, 375–397. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.D.; Johnson, J.A.; Winter, D.A. History and use of heat in pest control: A review. Int. J. Pest Manag. 2011, 57, 267–289. [Google Scholar] [CrossRef]
- Kells, S.A.; Goblirsch, M.J. Temperature and time requirements for controlling bed bugs (Cimex lectularius) under commercial heat treatment conditions. Insects 2011, 2, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Nayak, M.K.; Collins, P.J.; Throne, J.E.; Wang, J.-J. Biology and management of Psocids infesting stored products. Annu. Rev. Entomol. 2014, 59, 279–297. [Google Scholar] [CrossRef] [PubMed]
- Neven, L.G. Physiological responses of insects to heat. Postharvest Biol. Technol. 2000, 21, 103–111. [Google Scholar] [CrossRef]
- Hammond, D. Heat Treatment for Insect Control: Developments and Applications; Woodhead Publishing Series in Food Science, Technology and Nutrition: Number 241; Woodhead Publishing: Amsterdam, The Netherlands, 2015; pp. 1–120. ISBN 9780857097767. [Google Scholar]
- Mangan, R.L.; Hallman, G.J. Temperature treatments for quarantine security: New approaches for fresh commodities. In Temperature Sensitivity in Insects and Application in Integrated Pest Management; Hallman, G.J., Denlinger, D.L., Eds.; Westview Press: Boulder, CO, USA, 1998; pp. 201–236. ISBN 0813389909. [Google Scholar]
- Cira, T.M.; Venette, R.C.; Aigner, J.; Kuhar, T.; Mullins, D.E.; Gabbert, S.E.; Hutchison, W.D. Cold tolerance of Halyomorpha halys (Hemiptera: Pentatomidae) across geographic and temporal scales. Environ. Entomol. 2016, 45, 484–491. [Google Scholar] [CrossRef]
- Cira, T.M.; Koch, R.L.; Burkness, E.C.; Hutchison, W.D.; Venette, R.C. Effects of diapause on Halyomorpha halys (Hemiptera: Pentatomidae) cold tolerance. Environ. Entomol. 2018, 47, 997–1004. [Google Scholar] [CrossRef]
- Lowenstein, D.M.; Walton, V.M. Halyomorpha halys (Hemiptera: Pentatomidae) winter survival, feeding activity, and reproduction rates based on episodic cold shock and winter temperature regimes. J. Econ. Entomol. 2018, 111, 1210–1218. [Google Scholar] [CrossRef]
- Aigner, J.D.; Kuhar, T.P. Lethal high temperature extremes of the brown marmorated stink bug (Hemiptera: Pentatomidae) and efficacy of commercial heat treatments for control in expert shipping cargo. J. Agric. Urban Entomol. 2016, 32, 1–6. [Google Scholar] [CrossRef]
- Watanabe, M.; Arakawa, R.; Shinagawa, Y.; Okazawa, T. Overwintering flight of brown-marmorated stink bug, Halyomorpha mista to the buildings. Jpn. J. Sanit. Zool. 1994, 45, 25–31. [Google Scholar] [CrossRef]
- Lee, D.-H.; Cullum, J.P.; Anderson, J.L.; Daugherty, J.L.; Beckett, L.M.; Leskey, T.C. Characterization of overwintering sites of the invasive brown marmorated stink bug in natural landscapes using human surveyors and detector canines. PLoS ONE 2014, 9, e91575. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, A.L.; Chen, S.; Fleischer, S.J. Coupling developmental physiology, photoperiod, and temperature to model phenology and dynamics of an invasive heteropteran, Halyomorpha halys. Front. Physiol. 2016, 7, 165. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, A.L.; Fleischer, S.; Hamilton, G.C.; Hancock, T.; Krawczyk, G.; Lee, J.C.; Ogburn, E.; Pote, J.M.; Raudenbush, A.; Rucker, A.; et al. Phenology of brown marmorated stink bug described using female reproductive development. Ecol. Evol. 2017, 7, 6680–6690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skillman, V.P.; Wiman, N.G.; Lee, J.C. Nutrient declines in overwintering Halyomorpha halys populations. Entomol. Exp. Appl. 2018, 166, 778–789. [Google Scholar] [CrossRef]
- Skillman, V.P.; Wiman, N.G.; Lee, J.C. Monitoring nutrient status of Brown Marmorated Stink Bug adults and nymphs on Summer Holly. Insects 2018, 9, 120. [Google Scholar] [CrossRef] [PubMed]
- Costi, E.; Haye, T.; Maistrello, L. Biological parameters of the invasive brown marmorated stink bug, Halyomorpha halys, in southern Europe. J. Pest Sci. 2017, 90, 1059–1067. [Google Scholar] [CrossRef]
- Funayama, K. Importance of apple fruits as food for the brown-marmorated stink bug, Halyomorpha halys (Stål) (Heteroptera: Pentatomidae). Appl. Entomol. Zool. 2004, 39, 617–623. [Google Scholar] [CrossRef]
- SAS Institute. PROC User’s Manual, 6th ed.; SAS Institute: Cary, NC, USA, 2016; Available online: http://support.sas.com/documentation/cdl/en/indbug/68442/PDF/default/indbug.pdf (accessed on 26 August 2019).
- Robertson, J.L.; Savin, N.E.; Preisler, H.K.; Russell, R.M. Bioassays with Arthropods, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2007; pp. 1–224. ISBN 9781420004045. [Google Scholar]
- Bergh, J.C.; Morrison III, W.R.; Joseph, S.V.; Leskey, T.C. Characterizing spring emergence of adult Halyomorpha halys using experimental overwintering shelters and commercial pheromone traps. Entomol. Exp. Appl. 2017, 162, 336–345. [Google Scholar] [CrossRef]
- Lee, D.-H.; Leskey, T.C. Flight behavior of foraging and overwintering brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae). Bull. Entomol. Res. 2015, 105, 566–573. [Google Scholar] [CrossRef]
- Lee, D.-H.; Short, B.D.; Joseph, S.V.; Bergh, J.C.; Leskey, T.C. Review of the biology, ecology, and management of Halyomorpha halys (Hemiptera: Pentatomidae) in China, Japan, and the Republic of Korea. Environ. Entomol. 2013, 42, 627–641. [Google Scholar] [CrossRef]
- Loeschcke, V.; Krebs, R.A.; Barker, J.S.F. Genetic variation for resistance and acclimation to high temperature stress in Drosophila buzzatii. Biol. J. Linn. Soc. 1994, 52, 83–92. [Google Scholar] [CrossRef]
- Chown, S.L.; Jumbam, K.R.; Sørensen, J.G.; Terblanche, J.S. Phenotypic variance, plasticity and heritability estimates of critical thermal limits depend on methodological context. Funct. Ecol. 2009, 23, 133–140. [Google Scholar] [CrossRef]
- Overgaard, J.; Kristensen, T.N.; Sørensen, J.G. Validity of thermal ramping assays used to assess thermal tolerance in arthropods. PLoS ONE 2012, 7, e32758. [Google Scholar] [CrossRef] [PubMed]
- Terblanche, J.S.; Hoffmann, A.A.; Mitchell, K.A.; Rako, L.; le Roux, P.C.; Chown, S.L. Ecologically relevant measures of tolerance to potentially lethal temperatures. J. Exp. Biol. 2011, 214, 3713–3725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haye, T.; Abdallah, S.; Gariepy, T.; Wyniger, D. Phenology, life table analysis and temperature requirements of the invasive brown marmorated stink bug, Halyomorpha halys, in Europe. J. Pest Sci. 2014, 87, 407–418. [Google Scholar] [CrossRef]
- Ingels, C.A.; Daane, K.M. Phenology of brown marmorated stink bug in a California urban landscape. J. Econ. Entomol. 2018, 111, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Perkins, S.E.; Alexander, L.V. On the measurement of heat waves. J. Clim. 2013, 156, 4500–4517. [Google Scholar] [CrossRef]
- Perkins, S.E.; Alexander, L.V.; Nairn, J.R. Increasing frequency, intensity and duration of observed global heatwaves and warm spells. Geophys. Res. Lett. 2012, 39, L20714. [Google Scholar] [CrossRef]
- WMO Statement on the State of the Global Climate in 2018. Available online: https://library.wmo.int/doc_num.php?explnum_id=5789 (accessed on 26 August 2019).
- European Heatwave Sets New Temperature Records. Available online: https://public.wmo.int/en/media/news/european-heatwave-sets-new-temperature-records (accessed on 26 August 2019).
- 2019 Starts with Extreme, High-Impact Weather. Available online: https://public.wmo.int/en/media/news/2019-starts-extreme-high-impact-weather (accessed on 26 August 2019).
- Kriticos, D.J.; Kean, J.M.; Phillips, C.B.; Senay, S.D.; Acosta, H.; Haye, T. The potential global distribution of the brown marmorated stink bug, Halyomorpha halys, a critical threat to plant biosecurity. J. Pest Sci. 2017, 90, 1033–1043. [Google Scholar] [CrossRef]
- Zhu, G.; Bu, W.; Gao, Y.; Liu, G. Potential geographic distribution of brown marmorated stink bug invasion (Halyomorpha halys). PLoS ONE 2012, 7, e31246. [Google Scholar] [CrossRef]
- Zobel, E.S.; Hooks, C.R.R.; Dively, G.P. Seasonal abundance, host suitability, and feeding injury of the brown marmorated stink bug, Halyomorpha halys (Heteroptera: Pentatomidae), in selected vegetables. J. Econ. Entomol. 2016, 109, 1289–1302. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mortality rate (observed values, estimates of the curves, and 95% confidence interval (CI)) of Halyomorpha halys adults (exiting diapause (EXA) and entering diapause (ENA)) after exposure to high temperatures for (a) 2.5, (b) 15, (c) 30, and (d) 60 min.
Figure 1.
Mortality rate (observed values, estimates of the curves, and 95% confidence interval (CI)) of Halyomorpha halys adults (exiting diapause (EXA) and entering diapause (ENA)) after exposure to high temperatures for (a) 2.5, (b) 15, (c) 30, and (d) 60 min.
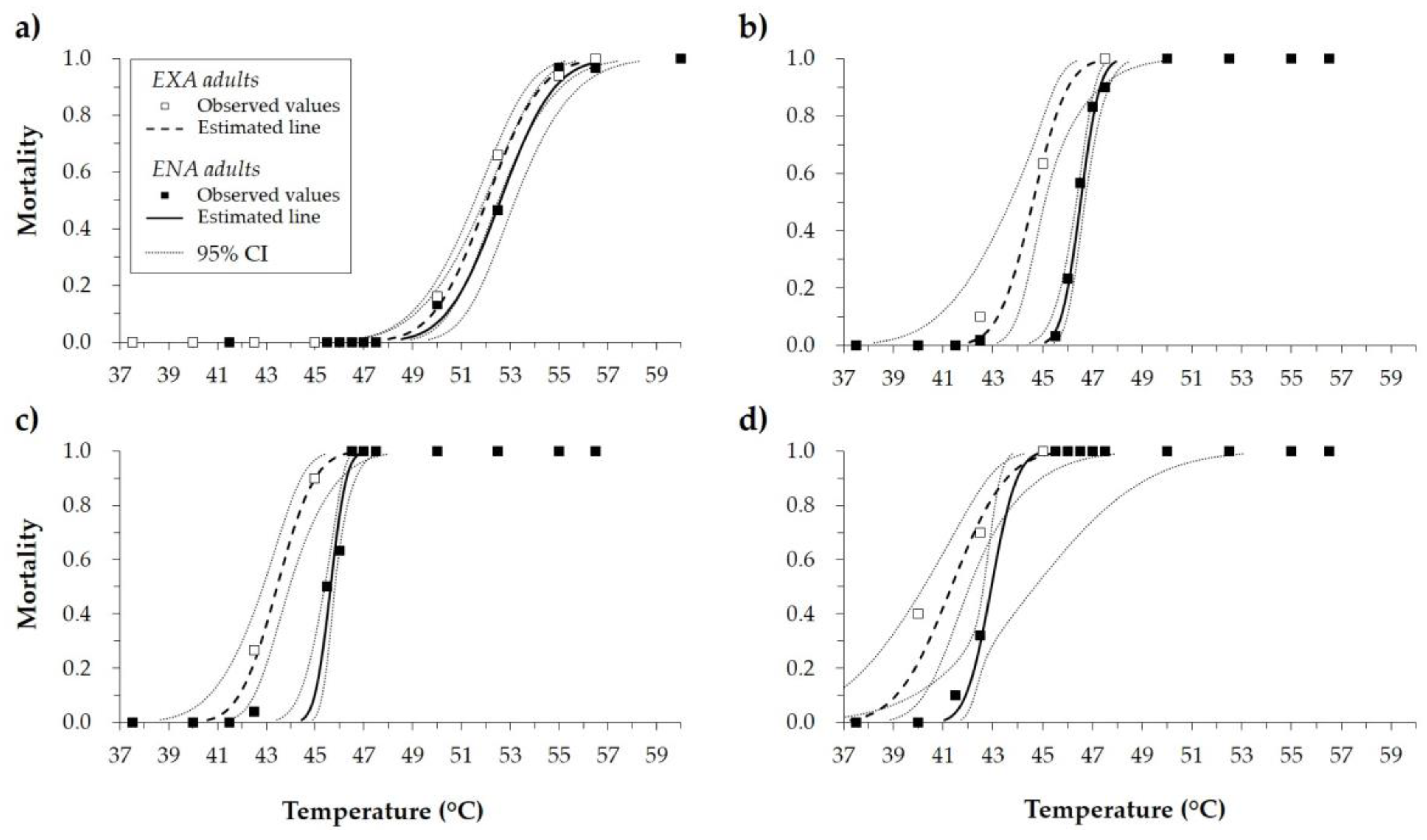
Figure 2.
Mortality rate (observed values, estimates of the curves, and 95% confidence interval (CI)) of Halyomorpha halys adults (EXA and ENA) calculated after different exposure times at the constant temperature of (a) 47.5 °C and (b) 50.0 °C.
Figure 2.
Mortality rate (observed values, estimates of the curves, and 95% confidence interval (CI)) of Halyomorpha halys adults (EXA and ENA) calculated after different exposure times at the constant temperature of (a) 47.5 °C and (b) 50.0 °C.

{kind=link}
{kind=link}
Table 1.
Lethal high temperatures (LT) 50 and 99 with 95% confidence interval (CI) for Halyomorpha halys adults of the two investigated states (i.e., ENA and EXA), with probit regression parameters.
Table 1.
Lethal high temperatures (LT) 50 and 99 with 95% confidence interval (CI) for Halyomorpha halys adults of the two investigated states (i.e., ENA and EXA), with probit regression parameters.
| Adults’ State | Exposure Time (Minutes) | n | LT50 (°C) * | 95% CILT50 (°C) | LT99 (°C) * | 95% CILT99 (°C) | Intercept | SEIntercept | Slope | SESlope | Χ2 ** | df | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lower | Upper | Lower | Upper | |||||||||||||
| EXA | 2.5 | 310 | 52.04 | b | 51.54 | 52.50 | 56.07 | b | 55.22 | 57.39 | −30.01 | 3.70 | 0.58 | 0.07 | 32.94 | 29 |
| ENA | 2.5 | 400 | 52.61 | a | 51.94 | 53.20 | 56.66 | a | 55.68 | 58.30 | −73.15 | 4.51 | 1.55 | 0.09 | 26.16 | 35 |
| EXA | 15.0 | 310 | 44.60 | b | 43.69 | 45.09 | 47.17 | b | 46.33 | 49.81 | −40.47 | 11.49 | 0.91 | 0.25 | 8.77 | 29 |
| ENA | 15.0 | 400 | 46.53 | a | 46.36 | 46.69 | 47.95 | a | 47.62 | 48.51 | −76.34 | 10.92 | 1.64 | 0.23 | 24.66 | 35 |
| EXA | 30.0 | 310 | 43.44 | b | 42.86 | 43.96 | 46.29 | b | 45.44 | 48.01 | −35.44 | 6.81 | 0.82 | 0.16 | 10.97 | 29 |
| ENA | 30.0 | 400 | 45.64 | a | 45.38 | 45.81 | 46.83 | a | 46.52 | 47.53 | −89.54 | 19.04 | 1.96 | 0.41 | 14.17 | 35 |
| EXA | 60.0 | 310 | 41.34 | b | 40.23 | 42.05 | 45.39 | a | 44.26 | 47.94 | −23.74 | 5.31 | 0.57 | 0.13 | 11.01 | 29 |
| ENA | 60.0 | 400 | 42.95 | a | 42.62 | 44.70 | 44.83 | b | 43.78 | 53.05 | −53.09 | 20.83 | 1.24 | 0.49 | 4.61 | 35 |
* Within a column, LT values for each exposure time pair with the same letter are not significantly different (p > 0.05) according to the lethal dose ratios method [43]. ** All Χ2 values fit the model at p > 0.05.
Table 2.
Time causing mortality (MT) 50 and 99 with 95% confidence interval (CI) for Halyomorpha halys adults exposed to high temperatures, with probit regression parameters.
Table 2.
Time causing mortality (MT) 50 and 99 with 95% confidence interval (CI) for Halyomorpha halys adults exposed to high temperatures, with probit regression parameters.
| Adults’ State | Temperature (°C) | n | MT50 (Minutes) * | 95% CIMT50 (Minutes) | MT99 (Minutes) * | 95% CIMT99 (Minutes) | Intercept | SEIntercept | Slope | SESlope | Χ2 ** | df | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lower | Upper | Lower | Upper | |||||||||||||
| EXA | 47.5 | 400 | 8.67 | a | 7.80 | 9.54 | 15.65 | b | 14.06 | 18.13 | −2.89 | 0.38 | 0.33 | 0.04 | 33.28 | 43 |
| ENA | 47.5 | 310 | 9.69 | a | 8.53 | 10.83 | 17.14 | a | 15.22 | 20.39 | −3.03 | 0.48 | 0.31 | 0.05 | 22.14 | 28 |
| EXA | 50.0 | 400 | 3.72 | a | 3.31 | 4.09 | 6.10 | b | 5.50 | 7.12 | −3.64 | 0.61 | 0.98 | 0.15 | 9.05 | 34 |
| ENA | 50.0 | 310 | 4.19 | a | 3.53 | 4.90 | 7.67 | a | 6.40 | 11.18 | −2.80 | 0.72 | 0.67 | 0.17 | 2.43 | 28 |
* Within a column, MT values for each exposure temperature pair (i.e., 47.5 or 50.0 °C) with the same letter are not significantly different (p > 0.05) according to the lethal dose ratios method [43]. ** All Χ2 values fit the model at p > 0.05.
Table 3.
Effects of Halyomorpha halys weight on tested variables and their interactions. Significant p-values are indicated in bold. The × is used to indicate the interaction between sources of variation.
Table 3.
Effects of Halyomorpha halys weight on tested variables and their interactions. Significant p-values are indicated in bold. The × is used to indicate the interaction between sources of variation.
| Source of Variation | 15 min | 30 min | 60 min | ||||||
|---|---|---|---|---|---|---|---|---|---|
| F Value | df | p-Value | F Value | df | p-Value | F Value | df | p-Value | |
| Status (dead or alive) | 0.10 | 1; 210 | 0.7517 | 0.38 | 1; 210 | 0.5404 | 0.17 | 1; 210 | 0.6850 |
| Temperature (Temp) | 3.47 | 1; 210 | 0.0639 | 2.47 | 1; 210 | 0.1173 | 1.20 | 1; 210 | 0.2739 |
| Status × Temp | 0.50 | 1; 210 | 0.4804 | 0.66 | 1; 210 | 0.4173 | 0.00 | 1; 210 | 0.9954 |
| Sex | 92.77 | 1; 210 | <0.0001 | 54.81 | 1; 210 | <0.0001 | 46.79 | 1; 210 | <0.0001 |
| Status × Sex | 0.31 | 1; 210 | 0.5793 | 1.47 | 1; 210 | 0.2273 | 0.27 | 1; 210 | 0.6033 |
| Sex × Temp | 0.46 | 1; 210 | 0.4968 | 0.27 | 1; 210 | 0.6066 | 0.83 | 1; 210 | 0.3621 |
| Status × Sex × Temp | 0.14 | 1; 210 | 0.7090 | 0.73 | 1; 210 | 0.3950 | 0.18 | 1; 210 | 0.6732 |
| Overwintering state (OS) | 22.12 | 1; 210 | <0.0001 | 13.88 | 1; 210 | 0.0003 | 16.35 | 1; 210 | <0.0001 |
| Status × OS | 0.08 | 1; 210 | 0.7759 | 0.41 | 1; 210 | 0.5202 | 0.30 | 1; 210 | 0.5815 |
| OS × Temp | 0.11 | 1; 210 | 0.7391 | 0.45 | 1; 210 | 0.5037 | 1.29 | 1; 210 | 0.2579 |
| Status × OS × Temp | 0.51 | 1; 210 | 0.4763 | 0.21 | 1; 210 | 0.6442 | 1.86 | 1; 210 | 0.1744 |
| Sex × OS | 0.76 | 1; 210 | 0.3844 | 1.84 | 1; 210 | 0.1765 | 1.37 | 1; 210 | 0.2439 |
| Status × Sex × OS | 0.07 | 1; 210 | 0.7867 | 0.18 | 1; 210 | 0.6746 | 0.62 | 1; 210 | 0.4324 |
| Sex × OS × Temp | 0.93 | 1; 210 | 0.3366 | 0.69 | 1; 210 | 0.4071 | 0.24 | 1; 210 | 0.6227 |
| Status × Sex × OS × Temp | 0.03 | 1; 210 | 0.8633 | 0.27 | 1; 210 | 0.6029 | 0.01 | 1; 210 | 0.9118 |
Table 4.
Effects of Halyomorpha halys nutrient index on tested variables and their interactions. Significant p-values are indicated in bold. The × is used to indicate the interaction between sources of variation.
Table 4.
Effects of Halyomorpha halys nutrient index on tested variables and their interactions. Significant p-values are indicated in bold. The × is used to indicate the interaction between sources of variation.
| Source of Variation | 15 min | 30 min | 60 min | ||||||
|---|---|---|---|---|---|---|---|---|---|
| F Value | df | p-Value | F Value | df | p-Value | F Value | df | p-Value | |
| Status (dead or alive) | 0.14 | 1; 210 | 0.7120 | 0.00 | 1; 210 | 0.9877 | 0.69 | 1; 210 | 0.4066 |
| Temperature (Temp) | 1.94 | 1; 210 | 0.1652 | 2.55 | 1; 210 | 0.1118 | 0.98 | 1; 210 | 0.3223 |
| Status × Temp | 0.16 | 1; 210 | 0.6930 | 0.84 | 1; 210 | 0.3595 | 0.02 | 1; 210 | 0.9024 |
| Sex | 1.34 | 1; 210 | 0.2481 | 2.87 | 1; 210 | 0.0915 | 3.88 | 1; 210 | 0.0501 |
| Status × Sex | 0.03 | 1; 210 | 0.8519 | 1.65 | 1; 210 | 0.1999 | 1.93 | 1; 210 | 0.1657 |
| Sex × Temp | 0.94 | 1; 210 | 0.3343 | 1.42 | 1; 210 | 0.2353 | 0.28 | 1; 210 | 0.6002 |
| Status × Sex × Temp | 0.67 | 1; 210 | 0.4144 | 0.17 | 1; 210 | 0.6766 | 0.13 | 1; 210 | 0.7237 |
| Overwintering state (OS) | 22.5 | 1; 210 | <0.0001 | 17.77 | 1; 210 | <0.0001 | 20.34 | 1; 210 | <0.0001 |
| Status × OS | 0.02 | 1; 210 | 0.8758 | 0.90 | 1; 210 | 0.3431 | 1.03 | 1; 210 | 0.3110 |
| OS × Temp | 0.29 | 1; 210 | 0.5913 | 0.81 | 1; 210 | 0.3701 | 0.49 | 1; 210 | 0.4832 |
| Status × OS × Temp | 0.32 | 1; 210 | 0.5732 | 0.62 | 1; 210 | 0.4313 | 0.19 | 1; 210 | 0.6646 |
| Sex × OS | 0.38 | 1; 210 | 0.5361 | 2.70 | 1; 210 | 0.1016 | 1.80 | 1; 210 | 0.1814 |
| Status × Sex × OS | 0.01 | 1; 210 | 0.9237 | 0.42 | 1; 210 | 0.5154 | 0.28 | 1; 210 | 0.5991 |
| Sex × OS × Temp | 3.88 | 1; 210 | 0.0501 | 1.80 | 1; 210 | 0.1817 | 0.53 | 1; 210 | 0.4692 |
| Status × Sex × OS × Temp | 0.00 | 1; 210 | 0.9530 | 0.00 | 1; 210 | 0.979 | 0.46 | 1; 210 | 0.5000 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Scaccini, D.; Duso, C.; Pozzebon, A. Lethal Effects of High Temperatures on Brown Marmorated Stink Bug Adults before and after Overwintering. Insects 2019, 10, 355. https://doi.org/10.3390/insects10100355
AMA Style
Scaccini D, Duso C, Pozzebon A. Lethal Effects of High Temperatures on Brown Marmorated Stink Bug Adults before and after Overwintering. Insects. 2019; 10(10):355. https://doi.org/10.3390/insects10100355
Chicago/Turabian StyleScaccini, Davide, Carlo Duso, and Alberto Pozzebon. 2019. "Lethal Effects of High Temperatures on Brown Marmorated Stink Bug Adults before and after Overwintering" Insects 10, no. 10: 355. https://doi.org/10.3390/insects10100355
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.