Is the Brood Pattern within a Honey Bee Colony a Reliable Indicator of Queen Quality?


Abstract
1. Introduction
2. Materials and Methods
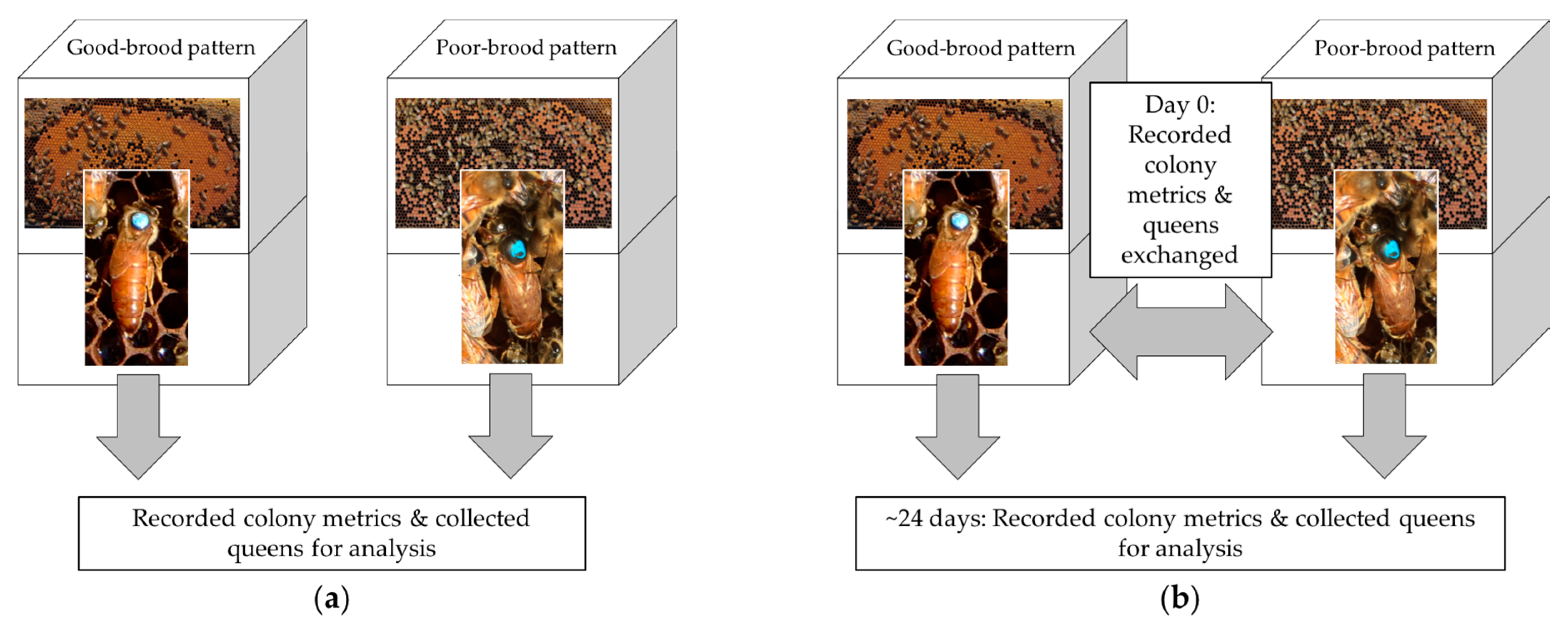
2.1. Colony Selection
2.2. Queen Mating Quality and Morphometric Measurements
2.3. Colony Measurements
2.4. Molecular Analysis
2.5. Statistical Analysis
3. Results
3.1. Brood Pattern Classifications
3.2. Measures Associated with Queens
3.3. Measures Associated with Colony Environment
3.3.1. Adult Bee Pathogens
3.3.2. Brood Pathogens
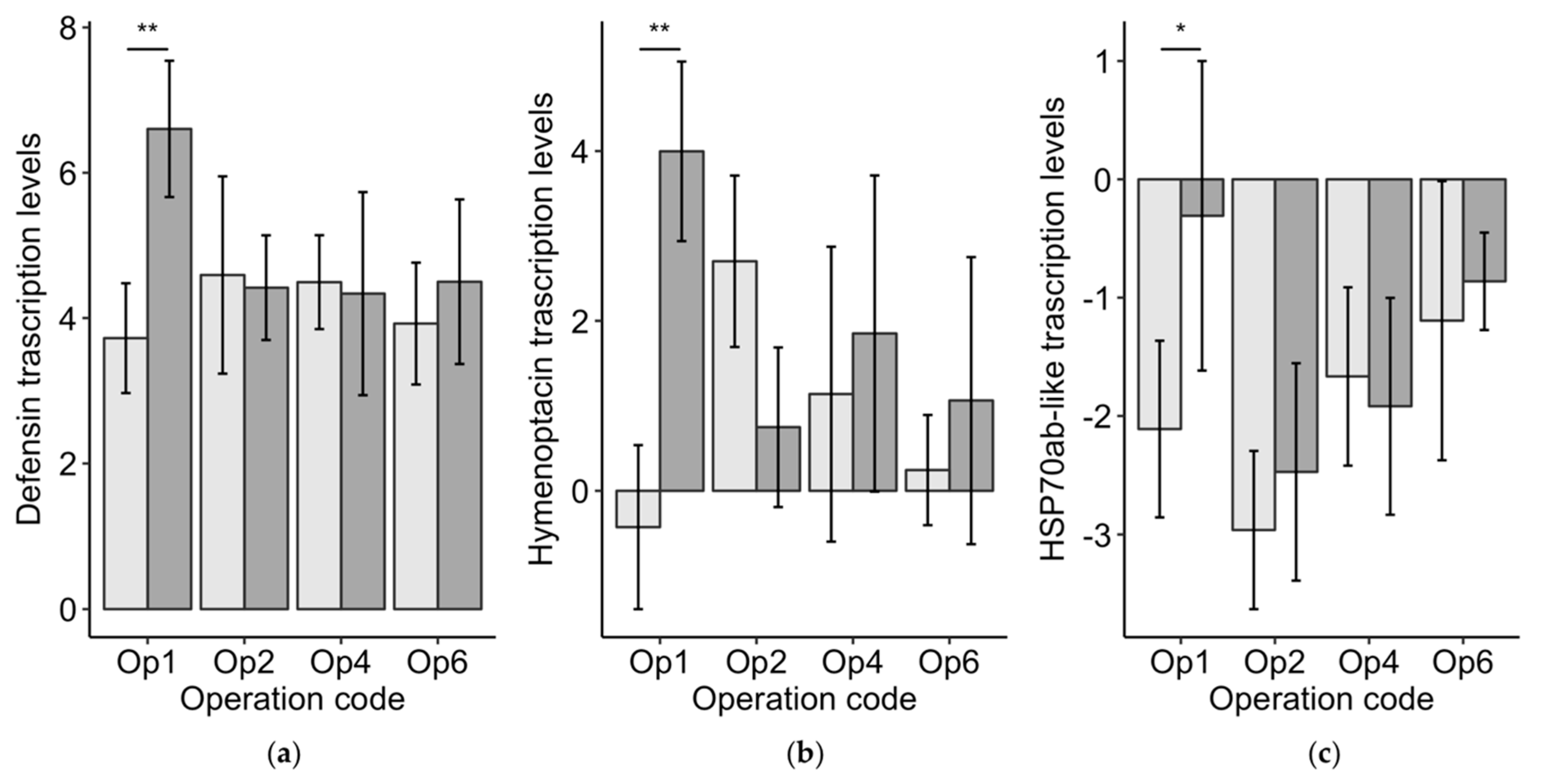
3.3.3. Worker Bee Vitellogenin, Immune Genes, and Heat Shock Protein
3.3.4. Colony Pesticide Levels
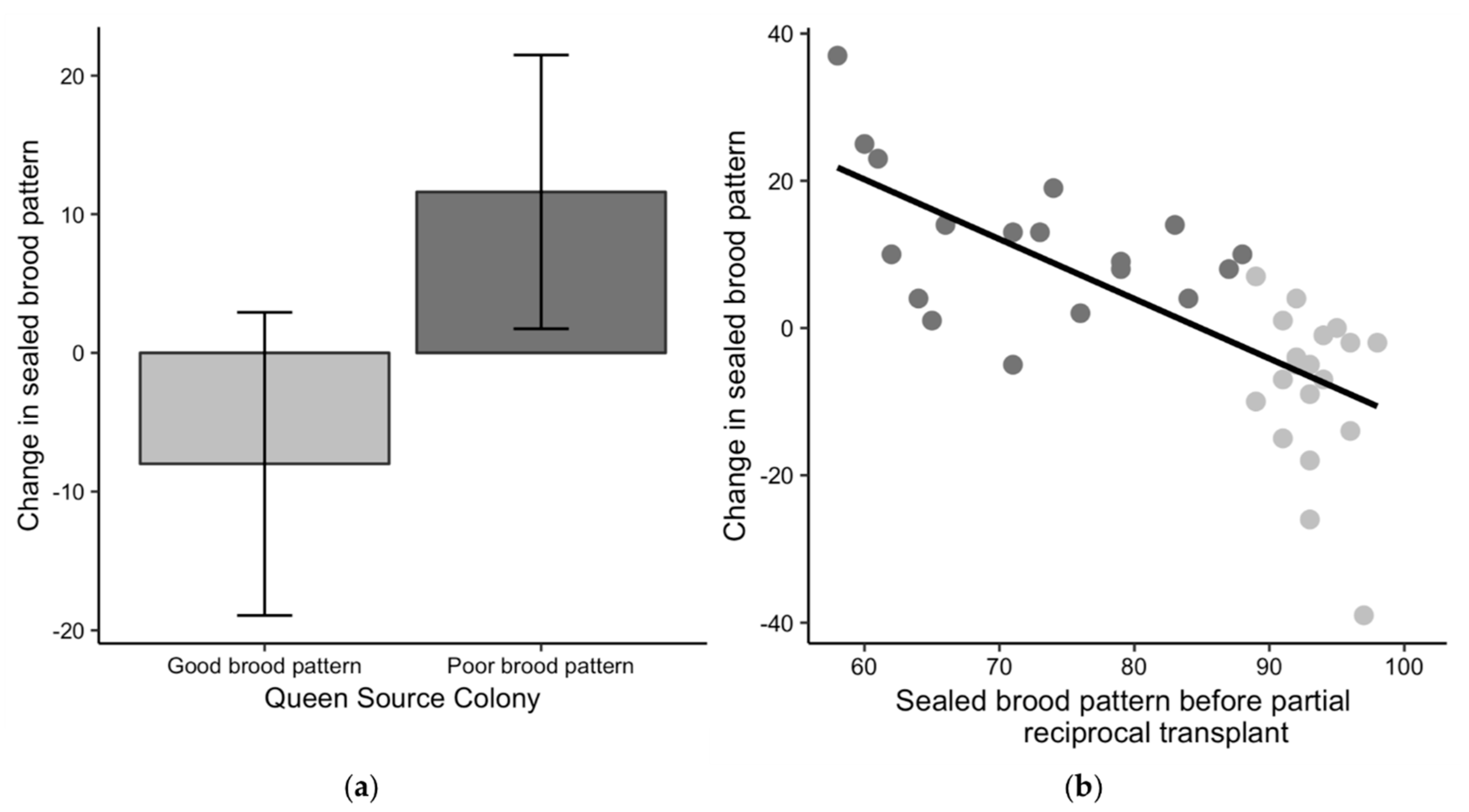
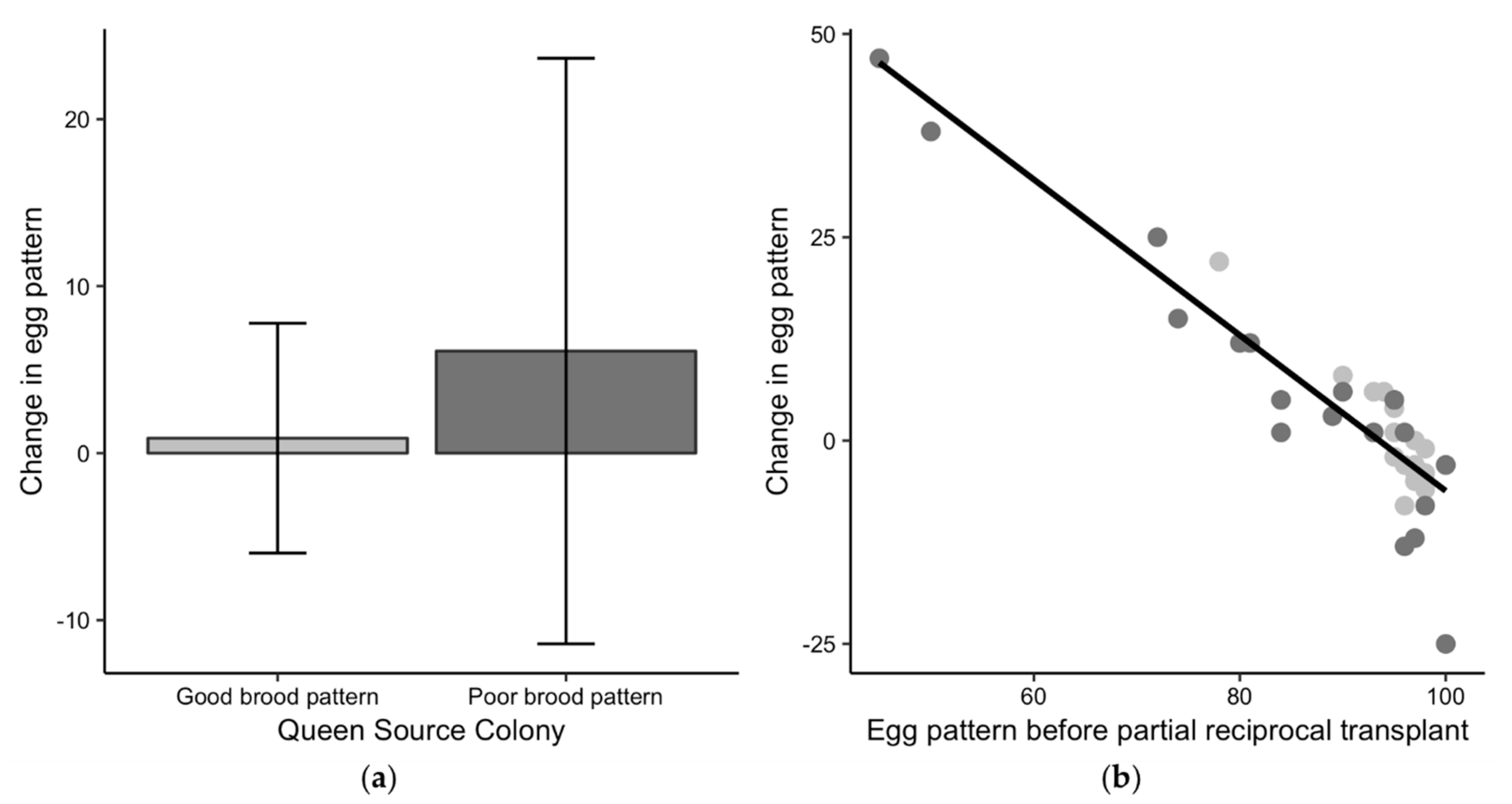
3.4. Brood Pattern Change
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, K.V.; Steinhauer, N.; Rennich, K.; Wilson, M.E.; Tarpy, D.R.; Caron, D.M.; Rose, R.; Delaplane, K.S.; Baylis, K.; Lengerich, E.J.; et al. A national survey of managed honey bee 2013–2014 annual colony losses in the USA. Apidologie 2015, 46, 292–305. [Google Scholar] [CrossRef]
- Kulhanek, K.; Steinhauer, N.; Rennich, K.; Caron, D.M.; Sagili, R.R.; Pettis, J.S.; Ellis, J.D.; Wilson, M.E.; Wilkes, J.T.; Tarpy, D.R.; et al. A national survey of managed honey bee 2015–2016 annual colony losses in the USA. J. Apic. Res. 2017, 56, 328–340. [Google Scholar] [CrossRef]
- Szabo, T.I. Length of Life of Queens in Honey Bee Colonies. Am. Bee J. 1993, 133, 723–724. [Google Scholar]
- Baer, B.; Collins, J.; Maalaps, K.; den Boer, S.P.A. Sperm use economy of honeybee (Apis mellifera) queens. Ecol. Evol. 2016, 6, 2877–2885. [Google Scholar] [CrossRef] [PubMed]
- Koeniger, G.; Koeniger, N.; Ellis, J.D.; Connor, L. Mating Biology of Honey Bees (Apis mellifera); Wicwas Press LLC: Kalamazoo, MI, USA, 2014; ISBN 978-1-878075-38-3. [Google Scholar]
- Tarpy, D.R.; Keller, J.J.; Caren, J.R.; Delaney, D.A. Assessing the Mating ‘Health’ of Commercial Honey Bee Queens. J. Econ. Entomol. 2012, 105, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Lodesani, M.; Balduzzi, D.; Galli, A. A study on spermatozoa viability over time in honey bee (Apis mellifera ligustica) queen spermathecae. J. Apic. Res. 2004, 43, 27–28. [Google Scholar] [CrossRef]
- Tarpy, D.R.; Olivarez, R. Measuring sperm viability over time in honey bee queens to determine patterns in stored-sperm and queen longevity. J. Apic. Res. 2014, 53, 493–495. [Google Scholar] [CrossRef]
- Collins, A.M. Relationship between semen quality and performance of instrumentally inseminated honey bee queens. Apidologie 2000, 31, 421–429. [Google Scholar] [CrossRef]
- Collins, A.M.; Williams, V.; Evans, J.D. Sperm storage and antioxidative enzyme expression in the honey bee, Apis mellifera. Insect Mol. Biol. 2004, 13, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Delaney, D.A.; Keller, J.J.; Caren, J.R.; Tarpy, D.R. The physical, insemination, and reproductive quality of honey bee queens (Apis mellifera L.). Apidologie 2011, 42, 1–13. [Google Scholar] [CrossRef]
- Amiri, E.; Strand, M.; Rueppell, O.; Tarpy, D. Queen Quality and the Impact of Honey Bee Diseases on Queen Health: Potential for Interactions between Two Major Threats to Colony Health. Insects 2017, 8, 48. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, L.; Ravallec, M.; Tournaire, M.; Cousserans, F.; Bergoin, M.; Dainat, B.; de Miranda, J.R. Viruses Associated with Ovarian Degeneration in Apis mellifera L. Queens. PLoS ONE 2011, 6, e16217. [Google Scholar] [CrossRef] [PubMed]
- Alaux, C.; Folschweiller, M.; McDonnell, C.; Beslay, D.; Cousin, M.; Dussaubat, C.; Brunet, J.-L.; Conte, Y.L. Pathological effects of the microsporidium Nosema ceranae on honey bee queen physiology (Apis mellifera). J. Invertebr. Pathol. 2011, 106, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Chaimanee, V.; Chantawannakul, P.; Chen, Y.; Evans, J.D.; Pettis, J.S. Effects of host age on susceptibility to infection and immune gene expression in honey bee queens (Apis mellifera) inoculated with Nosema ceranae. Apidologie 2014, 45, 451–463. [Google Scholar] [CrossRef]
- Haarmann, T.; Spivak, M.; Weaver, D.; Weaver, B.; Glenn, T. Effects of Fluvalinate and Coumaphos on Queen Honey Bees (Hymenoptera: Apidae) in Two Commercial Queen Rearing Operations. J. Econ. Entomol. 2002, 95, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Pettis, J.S.; Collins, A.M.; Wilbanks, R.; Feldlaufer, M.F. Effects of coumaphos on queen rearing in the honey bee, Apis mellifera. Apidologie 2004, 35, 605–610. [Google Scholar] [CrossRef]
- Burley, L.M.; Fell, R.D.; Saacke, R.G. Survival of Honey Bee (Hymenoptera: Apidae) Spermatozoa Incubated at Room Temperature from Drones Exposed to Miticides. J. Econ. Entomol. 2008, 101, 1081–1087. [Google Scholar] [CrossRef] [PubMed]
- Chaimanee, V.; Evans, J.D.; Chen, Y.; Jackson, C.; Pettis, J.S. Sperm viability and gene expression in honey bee queens (Apis mellifera) following exposure to the neonicotinoid insecticide imidacloprid and the organophosphate acaricide coumaphos. J. Insect Physiol. 2016, 89, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rangel, J.; Tarpy, D.R. In-Hive Miticides and their Effect on Queen Supersedure and Colony Growth in the Honey Bee (Apis mellifera). J. Environ. Anal. Toxicol. 2016, 6. [Google Scholar] [CrossRef]
- Sandrock, C.; Tanadini, M.; Tanadini, L.G.; Fauser-Misslin, A.; Potts, S.G.; Neumann, P. Impact of Chronic Neonicotinoid Exposure on Honeybee Colony Performance and Queen Supersedure. PLoS ONE 2014, 9, e103592. [Google Scholar] [CrossRef] [PubMed]
- Tsvetkov, N.; Samson-Robert, O.; Sood, K.; Patel, H.S.; Malena, D.A.; Gajiwala, P.H.; Maciukiewicz, P.; Fournier, V.; Zayed, A. Chronic exposure to neonicotinoids reduces honey bee health near corn crops. Science 2017, 356, 1395–1397. [Google Scholar] [CrossRef] [PubMed]
- Traynor, K.S.; Pettis, J.S.; Tarpy, D.R.; Mullin, C.A.; Frazier, J.L.; Frazier, M.; vanEngelsdorp, D. In-hive Pesticide Exposome: Assessing risks to migratory honey bees from in-hive pesticide contamination in the Eastern United States. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Dussaubat, C.; Maisonnasse, A.; Crauser, D.; Tchamitchian, S.; Bonnet, M.; Cousin, M.; Kretzschmar, A.; Brunet, J.-L.; Le Conte, Y. Combined neonicotinoid pesticide and parasite stress alter honeybee queens’ physiology and survival. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- vanEngelsdorp, D.; Tarpy, D.R.; Lengerich, E.J.; Pettis, J.S. Idiopathic brood disease syndrome and queen events as precursors of colony mortality in migratory beekeeping operations in the eastern United States. Prev. Vet. Med. 2013, 108, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.Y.; Anelli, C.M.; Sheppard, W.S. Sub-Lethal Effects of Pesticide Residues in Brood Comb on Worker Honey Bee (Apis mellifera) Development and Longevity. PLoS ONE 2011, 6, e14720. [Google Scholar] [CrossRef] [PubMed]
- Brodschneider, R.; Crailsheim, K. Nutrition and health in honey bees. Apidologie 2010, 41, 278–294. [Google Scholar] [CrossRef]
- Spivak, M.; Reuter, G.S. Performance of hygienic honey bee colonies in a commercial apiary. Apidologie 1998, 29, 291–302. [Google Scholar] [CrossRef]
- Harbo, J.R.; Harris, J.W. Responses to Varroa by honey bees with different levels of Varroa Sensitive Hygiene. J. Apic. Res. 2009, 48, 156–161. [Google Scholar] [CrossRef]
- Guzmán-Novoa, E.; Page, R.E. Selective Breeding of Honey Bees (Hymenoptera: Apidae) in Africanized Areas. J. Econ. Entomol. 1999, 92, 521–525. [Google Scholar] [CrossRef]
- Delaplane, K.S.; van der Steen, J.; Guzman-Novoa, E. Standard methods for estimating strength parameters of Apis mellifera colonies. J. Apic. Res. 2013, 52, 1–12. [Google Scholar] [CrossRef]
- Pettis, J.S.; Rice, N.; Joselow, K.; vanEngelsdorp, D.; Chaimanee, V. Colony Failure Linked to Low Sperm Viability in Honey Bee (Apis mellifera) Queens and an Exploration of Potential Causative Factors. PLoS ONE 2016, 11, e0147220. [Google Scholar] [CrossRef]
- Simone-Finstrom, M.; Tarpy, D.R. Honey Bee Queens Do Not Count Mates to Assess their Mating Success. J. Insect Behav. 2018, 31, 200–209. [Google Scholar] [CrossRef]
- Genersch, E.; von der Ohe, W.; Kaatz, H.; Schroeder, A.; Otten, C.; Büchler, R.; Berg, S.; Ritter, W.; Mühlen, W.; Gisder, S.; et al. The German bee monitoring project: A long term study to understand periodically high winter losses of honey bee colonies. Apidologie 2010, 41, 332–352. [Google Scholar] [CrossRef]
- Shimanuki, H.; Calderone, N.W.; Knox, D.A. Parasitic mite syndrome: The symptoms. Am. Bee J. 1994, 134, 117–119. [Google Scholar]
- vanEngelsdorp, D.; Evans, J.D.; Donovall, L.; Mullin, C.; Frazier, M.; Frazier, J.; Tarpy, D.R.; Hayes, J.; Pettis, J.S. “Entombed Pollen”: A new condition in honey bee colonies associated with increased risk of colony mortality. J. Invertebr. Pathol. 2009, 101, 147–149. [Google Scholar] [CrossRef] [PubMed]
- De Jong, D.; De Andrea Roma, D.; Gonçalves, L.S. A comparative analysis of shaking solutions for the detection of Varroa jacobsoni on adult honeybees. Apidologie 1982, 13, 297–306. [Google Scholar] [CrossRef]
- Cantwell, G.E. Standard methods for counting nosema spores. Am. Bee J. 1970, 110, 222–223. [Google Scholar]
- Traynor, K.S.; Rennich, K.; Forsgren, E.; Rose, R.; Pettis, J.; Kunkel, G.; Madella, S.; Evans, J.; Lopez, D.; vanEngelsdorp, D. Multiyear survey targeting disease incidence in US honey bees. Apidologie 2016, 47, 325–347. [Google Scholar] [CrossRef]
- Mullin, C.A.; Frazier, M.; Frazier, J.L.; Ashcraft, S.; Simonds, R.; vanEngelsdorp, D.; Pettis, J.S. High Levels of Miticides and Agrochemicals in North American Apiaries: Implications for Honey Bee Health. PLoS ONE 2010, 5, e9754. [Google Scholar] [CrossRef] [PubMed]
- Stoner, K.A.; Eitzer, B.D. Using a Hazard Quotient to Evaluate Pesticide Residues Detected in Pollen Trapped from Honey Bees (Apis mellifera) in Connecticut. PLoS ONE 2013, 8, e77550. [Google Scholar] [CrossRef] [PubMed]
- US EPA Ecotox Database. Available online: http://cfpub.epa.gov/ecotox/ (accessed on 26 May 2018).
- Hertfordshire Pesticide Properties Database. Available online: http://sitem.herts.ac.uk/aeru/ppdb/en/ index.htm (accessed on 26 May 2018).
- Sanchez-Bayo, F.; Goka, K. Pesticide Residues and Bees—A Risk Assessment. PLoS ONE 2014, 9, e94482. [Google Scholar] [CrossRef] [PubMed]
- Alburaki, M.; Chen, D.; Skinner, J.; Meikle, W.; Tarpy, D.; Adamczyk, J.; Stewart, S. Honey Bee Survival and Pathogen Prevalence: From the Perspective of Landscape and Exposure to Pesticides. Insects 2018, 9, 65. [Google Scholar] [CrossRef] [PubMed]
- Amdam, G.V.; Omholt, S.W. The Regulatory Anatomy of Honeybee Lifespan. J. Theor. Biol. 2002, 216, 209–228. [Google Scholar] [CrossRef] [PubMed]
- Corona, M.; Velarde, R.A.; Remolina, S.; Moran-Lauter, A.; Wang, Y.; Hughes, K.A.; Robinson, G.E. Vitellogenin, juvenile hormone, insulin signaling, and queen honey bee longevity. Proc. Natl. Acad. Sci. USA 2007, 104, 7128–7133. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.M.; Ihle, K.E.; Fondrk, M.K.; Page, R.E.; Amdam, G.V. The Gene vitellogenin Has Multiple Coordinating Effects on Social Organization. PLoS Biol. 2007, 5, e62. [Google Scholar] [CrossRef] [PubMed]
- Seehuus, S.-C.; Norberg, K.; Gimsa, U.; Krekling, T.; Amdam, G.V. Reproductive protein protects functionally sterile honey bee workers from oxidative stress. Proc. Natl. Acad. Sci. USA 2006, 103, 962–967. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.D.; Pettis, J.S. Colony-level impacts of immune responsiveness in honey bees, Apis mellifera. Evolution 2005, 59, 2270–2274. [Google Scholar] [CrossRef] [PubMed]
- Simone, M.; Evans, J.D.; Spivak, M. Resin collection and social immunity in honey bees. Evolution 2009, 63, 3016–3022. [Google Scholar] [CrossRef] [PubMed]
- Even, N.; Devaud, J.-M.; Barron, A. General Stress Responses in the Honey Bee. Insects 2012, 3, 1271–1298. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. Available online: https://www.R-project.org/ (accessed on 26 May 2018).
- vanEngelsdorp, D.; Lengerich, E.; Spleen, A.; Dainat, B.; Cresswell, J.; Baylis, K.; Nguyen, B.K.; Soroker, V.; Underwood, R.; Human, H.; et al. Standard epidemiological methods to understand and improve Apis mellifera health. J. Apic. Res. 2013, 52, 1–16. [Google Scholar] [CrossRef]
- Haldane, J.B.S. The mean and variance of the moments of chi-squaredwhen used as a test of homogeneity, when expectations are small. Biometrika 1940, 29, 133–134. [Google Scholar]
- Anscombe, F.J. On estimating binomial response relations. Biometrika 1956, 4, 461–464. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Woyke, J. Natural and artificial insemination of queen honeybees. Bee World 1962, 43, 21–22. [Google Scholar] [CrossRef]
- Giacobino, A.; Molineri, A.; Cagnolo, N.B.; Merke, J.; Orellano, E.; Bertozzi, E.; Masciángelo, G.; Pietronave, H.; Pacini, A.; Salto, C.; et al. Risk factors associated with failures of Varroa treatments in honey bee colonies without broodless period. Apidologie 2015, 46, 573–582. [Google Scholar] [CrossRef]
- Tarpy, D.R.; vanEngelsdorp, D.; Pettis, J.S. Genetic diversity affects colony survivorship in commercial honey bee colonies. Naturwissenschaften 2013, 100, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Niño, E.L.; Malka, O.; Hefetz, A.; Tarpy, D.R.; Grozinger, C.M. Chemical Profiles of Two Pheromone Glands Are Differentially Regulated by Distinct Mating Factors in Honey Bee Queens (Apis mellifera L.). PLoS ONE 2013, 8, e78637. [Google Scholar] [CrossRef] [PubMed]
- Kocher, S.D.; Grozinger, C.M. Cooperation, Conflict, and the Evolution of Queen Pheromones. J. Chem. Ecol. 2011, 37, 1263–1275. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Paper (Year) | Brood Pattern | n | Sperm Count, Millions (Range) | Poorly Mated 1 | Sperm Viability (%) | Weight (mg) | Thorax Width (mm) | Head Width (mm) |
|---|---|---|---|---|---|---|---|---|
| This study (2016) | Good-brood | 17 | 6.74 ± 1.95 (2.55–9.37) | 6% | 83.7 ± 6.3 | 223.9 ± 17.1 | 4.89 ± 0.15 | 3.79 ± 0.14 |
| Poor-brood | 17 | 5.07 ± 2.51 (0.52–8.09) | 24% | 78.3 ± 11.2 | 216.5 ± 23.0 | 4.85 ± 0.18 | 3.8 ± 0.12 | |
| This study (2017) | Good-brood | 19 | 5.69 ± 1.82 (1.39–8.4) | 11% | 78.0 ± 6.4 | 223.6 ± 27.9 | 4.93 ± 0.19 | 3.83 ± 0.10 |
| Poor-brood | 18 | 5.88 ± 1.57 (2.8–8.1) | 6% | 78.3 ± 10.9 2 | 231.8 ± 23.0 | 4.94 ± 0.19 | 3.83 ± 0.09 | |
| Delaney et al. (2011) | NA | 114 | 3.99 ± 1.50 (0.2–9.0) | 18.6% | NA | 184.8 ± 21.7 | 4.35 ± 0.19 | 3.62 ± 0.12 |
| Tarpy et al. (2012) | NA | 61 | 4.37 ± 1.45 | 13.6%, & 1 virgin queen | 83.7 ± 3.3 | 218.7 ± 20.7 | 4.34 ± 0.23 | 3.45 ± 0.23 |
| % of Samples with Positive Detections | ||||
|---|---|---|---|---|
| Sample Type (Year) | Factor | Good-Brood | Poor-Brood | Odds Ratio (95% CI) |
| Queens (2016) | No pathogens detected | 33 | 13 | 0.31 (0.05–1.93) |
| Black Queen Cell Virus | 7 | 0 | 0.31 (0.01–8.29) | |
| Deformed Wing Virus type A | 40 | 60 | 2.25 (0.52–9.70) | |
| Deformed Wing Virus type B | 53 | 73 | 2.41 (0.52–11.1) | |
| Lake Sinai Virus | 13 | 0 | 0.17 (0.01–3.96) | |
| Queens (2017) 1 | No pathogens detected | 79 | 78 | 0.93 (0.2–4.47) |
| Deformed Wing Virus type A | 5 | 11 | 2.25 (0.19–27.22) | |
| Deformed Wing Virus type B | 16 | 17 | 1.07 (0.19–6.13) | |
| Israeli Acute Paralysis Virus | 5 | 0 | 0.33 (0.01–8.73) | |
| Worker bees (2016) | >3 Varroa mites per bee | 6 | 6 | 1.00 (0.06–17.41) |
| >1 million Nosema spores per bee, by microscopy | 12 | 18 | 1.61 (0.23–11.09) | |
| Worker bees (2017) 2 | >3 Varroa mites per bee | 0 | 5 | 3.15 (0.12–81.74) |
| >1 million Nosema spores per bee, by microscopy | 33 | 48 | 1.82 (0.52–6.33) | |
| Acute Bee Paralysis Virus | 10 | 5 | 0.48 (0.04–5.68) | |
| Black Queen Cell Virus | 38 | 19 | 0.38 (0.09–1.55) | |
| Deformed Wing Virus type A | 5 | 14 | 3.33 (0.32–34.99) | |
| Deformed Wing Virus type B | 24 | 43 | 2.40 (0.64–9.03) | |
| Israeli Acute Paralysis Virus | 19 | 5 | 0.21 (0.02–2.09) | |
| Lake Sinai Virus | 100 | 100 | 1.00 (0.02–52.74) | |
| Trypanosomes | 10 | 19 | 2.24 (0.36–13.78) | |
| Nosema spp., by PCR | 90 | 86 | 0.63 (0.09–4.23) | |
| Brood disease (2016) | Chalkbrood | 6 | 0 | 0.31 (0.01–8.27) |
| Brood disease (2017) | Chalkbrood 3 | 52 | 76 | 2.91 (0.78–10.89) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, K.V.; Goblirsch, M.; McDermott, E.; Tarpy, D.R.; Spivak, M. Is the Brood Pattern within a Honey Bee Colony a Reliable Indicator of Queen Quality? Insects 2019, 10, 12. https://doi.org/10.3390/insects10010012
Lee KV, Goblirsch M, McDermott E, Tarpy DR, Spivak M. Is the Brood Pattern within a Honey Bee Colony a Reliable Indicator of Queen Quality? Insects. 2019; 10(1):12. https://doi.org/10.3390/insects10010012
Chicago/Turabian StyleLee, Kathleen V., Michael Goblirsch, Erin McDermott, David R. Tarpy, and Marla Spivak. 2019. "Is the Brood Pattern within a Honey Bee Colony a Reliable Indicator of Queen Quality?" Insects 10, no. 1: 12. https://doi.org/10.3390/insects10010012
APA StyleLee, K. V., Goblirsch, M., McDermott, E., Tarpy, D. R., & Spivak, M. (2019). Is the Brood Pattern within a Honey Bee Colony a Reliable Indicator of Queen Quality? Insects, 10(1), 12. https://doi.org/10.3390/insects10010012