
Gut–Lung Axis in Focus: Deciphering the Impact of Gut Microbiota on Pulmonary Arterial Hypertension


 , and
, and {kind=link}
Abstract
:1. Introduction
2. Changes in the Gut Microbiota Composition in PAH
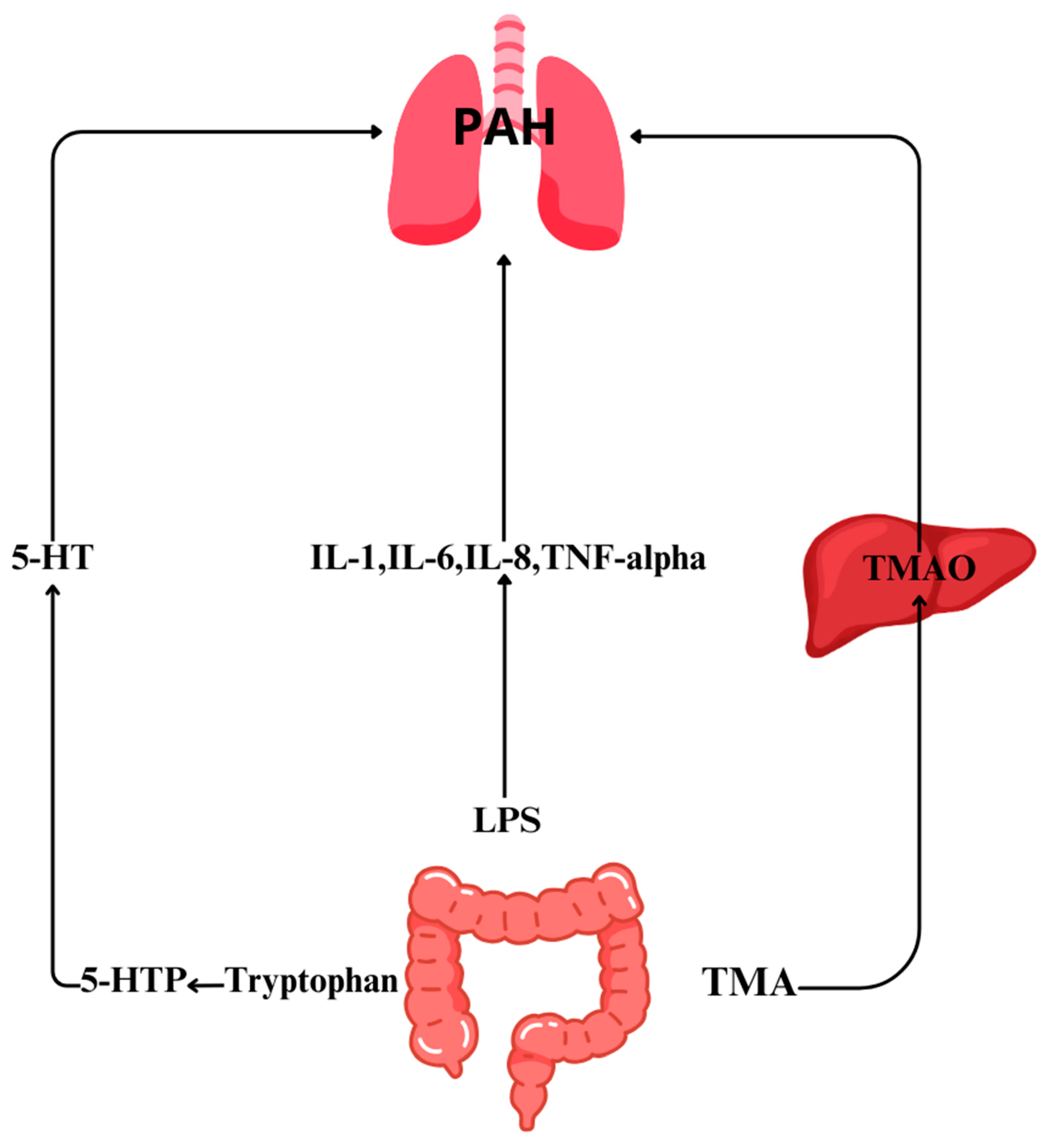
3. Role of Gut Microbiome Metabolites in PAH
3.1. Trimethylamine N-Oxide (TMAO)
3.2. Short-Chain Fatty Acids
3.3. Serotonin
3.4. Lipopolysaccharides
4. Role of Other Diseases and Intestinal Dysbiosis in PAH
4.1. Systemic Lupus Erythematosus
4.2. Infection with Human Immunodeficiency Virus (HIV)
4.3. Schistosomiasis in PAH Development
5. Perspectives on Gut Microbiota in PAH Treatment
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Martin-Gallausiaux, C.; Marinelli, L.; Blottière, H.M.; Larraufie, P.; Lapaque, N. SCFA: Mechanisms and Functional Importance in the Gut. Proc. Nutr. Soc. 2021, 80, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Callejo, M.; Barberá, J.A.; Duarte, J.; Perez-Vizcaino, F. Impact of Nutrition on Pulmonary Arterial Hypertension. Nutrients 2020, 12, 169. [Google Scholar] [CrossRef] [PubMed]
- Paone, P.; Cani, P.D. Mucus Barrier, Mucins and Gut Microbiota: The Expected Slimy Partners? Gut 2020, 69, 2232–2243. [Google Scholar] [CrossRef] [PubMed]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [PubMed]
- Braune, A.; Blaut, M. Bacterial species involved in the conversion of dietary flavonoids in the human gut. Gut Microbes 2016, 7, 216–234. [Google Scholar] [CrossRef] [PubMed]
- Beaugerie, L.; Petit, J.C. Antibiotic-associated diarrhoea. Best. Pract. Res. Clin. Gastroenterol. 2004, 18, 337–352. [Google Scholar] [CrossRef] [PubMed]
- Faderl, M.; Noti, M.; Corazza, N.; Mueller, C. Keeping bugs in check: The mucus layer as a critical component in maintaining intestinal homeostasis. IUBMB Life 2015, 67, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Quigley, E.M. Gut bacteria in health and disease. Gastroenterol. Hepatol. 2013, 9, 560–569. [Google Scholar]
- Shen, S.; Wong, C.H. Bugging inflammation: Role of the gut microbiota. Clin. Transl. Immunol. 2016, 5, e72. [Google Scholar] [CrossRef]
- Nagpal, R.; Yadav, H. Bacterial translocation from the gut to the distant organs: An overview. Ann. Nutr. Metab. 2017, 71 (Suppl. S1), 11–16. [Google Scholar] [CrossRef]
- Takiishi, T.; Fenero, C.I.M.; Camara, N.O.S. Intestinal barrier and gut microbiota: Shaping our immune responses throughout life. Tissue Barriers 2017, 5, e1373208. [Google Scholar] [CrossRef] [PubMed]
- Hong, W.; Mo, Q.; Wang, L.; Peng, F.; Zhou, Y.; Zou, W.; Sun, R.; Liang, C.; Zheng, M.; Li, H.; et al. Changes in the gut microbiome and metabolome in a rat model of pulmonary arterial hypertension. Bioengineered 2021, 12, 5173–5183. [Google Scholar] [CrossRef] [PubMed]
- Callejo, M.; Mondejar-Parreño, G.; Barreira, B.; Izquierdo-Garcia, J.L.; Morales-Cano, D.; Esquivel-Ruiz, S.; Moreno, L.; Cogolludo, Á; Duarte, J.; Perez-Vizcaino, F. Pulmonary Arterial Hypertension Affects the Rat Gut Microbiome. Sci. Rep. 2018, 8, 9681. [Google Scholar] [CrossRef] [PubMed]
- Sanada, T.J.; Hosomi, K.; Shoji, H.; Park, J.; Naito, A.; Ikubo, Y.; Yanagisawa, A.; Kobayashi, T.; Miwa, H.; Suda, R.; et al. Gut microbiota modification suppresses the development of pulmonary arterial hypertension in an SU5416/hypoxia rat model. Pulm. Circ. 2020, 10, 2045894020929147. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.K.; Oliveira, A.C.; Yang, T.; Kim, S.; Zubcevic, J.; Aquino, V.; Lobaton, G.O.; Goel, R.; Richards, E.M.; Raizada, M.K. Pulmonary arterial hypertension-associated changes in gut pathology and microbiota. ERJ Open Res. 2020, 6, 253. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Yin, H.; Gou, D. Gut Microbiota and Metabolome Changes in Three Pulmonary Hypertension Rat Models. Microorganisms 2023, 11, 472. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Rigatto, K.; Gazzana, M.B.; Knorst, M.M.; Richards, E.M.; Pepine, C.J.; Raizada, M.K. Altered Gut Microbiome Profile in Patients With Pulmonary Arterial Hypertension. Hypertension 2020, 75, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, T.; Lu, W.; Duan, X.; Luo, X.; Liu, S.; Chen, Y.; Li, Y.; Chen, J.; Liao, J.; et al. Altered Airway Microbiota Composition in Patients With Pulmonary Hypertension. Hypertension 2020, 76, 1589–1599. [Google Scholar] [CrossRef]
- Moutsoglou, D.M.; Tatah, J.; Prisco, S.Z.; Prins, K.W.; Staley, C.; Lopez, S.; Blake, M.; Teigen, L.; Kazmirczak, F.; Weir, E.K.; et al. Pulmonary Arterial Hypertension Patients Have a Proinflammatory Gut Microbiome and Altered Circulating Microbial Metabolites. Am. J. Respir. Crit. Care Med. 2023, 207, 740–756. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The Human Microbiome Project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef]
- Karoor, V.; Strassheim, D.; Sullivan, T.; Verin, A.; Umapathy, N.S.; Dempsey, E.C.; Frank, D.N.; Stenmark, K.R.; Gerasimovskaya, E. The Short-Chain Fatty Acid Butyrate Attenuates Pulmonary Vascular Remodeling and Inflammation in Hypoxia-Induced Pulmonary Hypertension. Int. J. Mol. Sci. 2021, 22, 9916. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, B.; Li, X.; Xue, L.; Liu, B.; Liang, Y.; Zhao, Z.; Luo, Q.; Liu, Z.; Zeng, Q.; et al. Higher circulating Trimethy lamine N-oxide levels are associated with worse severity and prognosis in pulmonary hypertension: A cohort study. Respir. Res. 2022, 14, 344. [Google Scholar] [CrossRef] [PubMed]
- MacLean, M.M.R. The serotonin hypothesis in pulmonary hypertension revisited: Targets for novel therapies (2017 Grover Conference Series). Pulm. Circ. 2018, 8, 2045894018759125. [Google Scholar] [CrossRef] [PubMed]
- Cho, C.E.; Taesuwan, S.; Malysheva, O.V.; Bender, E.; Tulchinsky, N.F.; Yan, J.; Sutter, J.L.; Caudill, M.A. Trimethylamine-N-oxide (TMAO) response to animal source foods varies among healthy young men and is influenced by their gut microbiota composition: A randomized controlled trial. Mol. Nutr. Food Res. 2017, 61, 1600324. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Ivashkin, V.T.; Kashukh, Y.A. Impact of L-carnitine and phosphatidylcholine containing products on the proatherogenic metabolite TMAO production and gut microbiome changes in patients with coronary artery disease. Vopr. Pitan. 2019, 88, 25–33. [Google Scholar] [PubMed]
- Tan, Y.; Sheng, Z.; Zhou, P.; Liu, C.; Zhao, H.; Song, L.; Li, J.; Zhou, J.; Chen, Y.; Wang, L.; et al. Plasma trimethylamine N-oxide as a novel biomarker for plaque rupture in patients with ST-seg ment-elevation myocardial infarction. Circ. Cardiovasc. Interv. 2019, 12, e007281. [Google Scholar] [CrossRef]
- Papandreou, C.; Bulló, M.; Hernández-Alonso, P.; Ruiz-Canela, M.; Li, J.; Guasch-Ferré, M.; Toledo, E.; Clish, C.; Corella, D.; Estruch, R.; et al. Choline Metabolism and Risk of Atrial Fibrillation and Heart Failure in the PREDIMED Study. Clin. Chem. 2021, 67, 288–297. [Google Scholar] [CrossRef]
- Ge, X.; Zheng, L.; Zhuang, R.; Yu, P.; Xu, Z.; Liu, G.; Xi, X.; Zhou, X.; Fan, H. The Gut Microbial Metabolite Trimethylamine N-Oxide and Hypertension Risk: A Systematic Review and Dose-Response Meta-analysis. Adv. Nutr. 2020, 11, 66–76. [Google Scholar] [CrossRef]
- Zhu, W.; Gregory, J.; Org, E.; Buffa, J.; Gupta, N.; Wang, Z.; Li, L.; Fu, X.; Wu, Y.; Mehrabian, M. Gut microbial metabolite TMAO enhances platelet hyperreactivity and thrombosis risk. Cell 2016, 165, 111–124. [Google Scholar] [CrossRef]
- Li, T.; Chen, Y.; Gu, C.; Li, X. Elevated circulating trimethylamine N-oxide levels contribute to endothelial dysfunction in aged rats through vascular inflammation and oxidative stress. Front. Physiol. 2017, 8, 350. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zeng, Q.; Gao, J.; Yang, B.; Zhou, J.; Li, K.; Li, L.; Wang, A.; Li, X.; Liu, Z.; et al. High-circulating gut microbiota-dependent metabolite trimethylamine N-oxide is associated with poor prognosis in pulmonary arterial hypertension. Eur. Heart. J. Open. 2022, 2, oeac021. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The role of short-chain fatty acids in health and disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar] [PubMed]
- Macfarlane, S.; Macfarlane, G.T. Regulation of short-chain fatty acid production. Proc. Nutr. Soc. 2003, 62, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Topping, D.L.; Clifton, P.M. Short-chain fatty acids and human colonic function: Roles of resistant starch and nonstarch polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Lee, G.; Son, H.; Koh, H.; Kim, E.S.; Unno, T.; Shin, J.H. Butyrate producers, “The Sentinel of Gut”: Their intestinal significance with and beyond butyrate, and prospective use as microbial therapeutics. Front. Microbiol. 2023, 13, 1103836. [Google Scholar] [CrossRef]
- Ye, Z.; Zhang, N.; Wu, C.; Zhang, X.; Wang, Q.; Huang, X.; Du, L.; Cao, Q.; Tang, J.; Zhou, C.; et al. A metagenomic study of the gut microbiome in Behcet’s disease. Microbiome 2018, 6, 135. [Google Scholar] [CrossRef]
- Cait, A.; Hughes, M.R. Microbiome-driven allergic lung inflammation is ameliorated by short chain fatty acids. Mucosal Immunol. 2018, 11, 785–796. [Google Scholar] [CrossRef]
- Pulgarin, A.; Methe, B.; Morris, A.; Al Ghouleh, I. Abstract 12755, Role of Micriobiome-Derived Short-Chain Fatty Acid Butyrate in Pulmonary Hypertension. Circulation 2022, 146, A12755. [Google Scholar] [CrossRef]
- American Heart Association Journals. Available online: https://www.ahajournals.org/doi/abs/10.1161/circ.136.suppl_1.20620 (accessed on 8 November 2023).
- Schippa, S.; Conte, M.P. Dysbiotic events in gut microbiota: Impact on human health. Nutrients 2014, 6, 5786–5805. [Google Scholar] [CrossRef]
- Siegel, G.J.; Agranoff, B.W. Basic Neurochemistry: Molecular, Cellular and Medical Aspects, 6th ed.; Lippincott-Raven: Philadelphia, PA, USA, 1999; p. 227. [Google Scholar]
- Papamatheakis, D.G.; Vemulakonda, S.; Blood, Q.; Goyal, R.; Rubalcava, M.; Vrancken, K.; Bennett, A.; Dawson, A.; Osman, N.J.; Blood, A.B.; et al. Preservation of serotonin-mediated contractility in adult sheep pulmo-nary arteries following long-term high-altitude hypoxia. High Alt. Med. Biol. 2011, 12, 253–264. [Google Scholar]
- MacLean, M.R.; Sweeney, G.; Baird, M.; McCulloch, K.M.; Houslay, M.; Morecroft, I. 5-Hydroxytryptamine receptors mediating vasoconstriction in pulmonary arteries from control and pulmonary hypertensive rats. Br. J. Pharmacol. 1996, 119, 917–930. [Google Scholar] [CrossRef] [PubMed]
- MacLean, M.R.; Clayton, R.A.; Hillis, S.W.; McIntyre, P.D.; Peacock, A.J.; Templeton, A.G. 5-HT1-receptor-mediated vasoconstriction in bovine isolated pulmonary arteries: Influences of vascular endothelium and tone. Pulm. Pharmacol. 1994, 7, 65–72. [Google Scholar] [CrossRef] [PubMed]
- MacLean, M.R. Pulmonary hypertension, anorexigens and 5-HT: Pharmacological synergism in action? Trends Pharmacol. Sci. 1999, 20, 490–495. [Google Scholar]
- Baranowska-Kuczko, M.; Kozłowska, H.; Schlicker, E.; Göthert, M.; MacLean, M.R.; Kozłowski, M.; Kloza, M.; Sadowska, O.; Malinowska, B. Reduction of the serotonin 5-HT1B and 5-HT2A receptor-mediated contraction of human pulmonary artery by the combined 5-HT1B receptor antagonist and serotonin transporter inhibitor LY393558. Pharmacol. Rep. 2020, 72, 756–762. [Google Scholar] [CrossRef] [PubMed]
- Entrez Gene: TPH1 Tryptophan Hydroxylase 1 (Tryptophan 5-Monooxygenase). Available online: https://www.ncbi.nlm.nih.gov/gene?Db=gene&Cmd=ShowDetailView&TermToSearch=7166 (accessed on 8 November 2023).
- Aiello, R.J.; Bourassa, P.A.; Zhang, Q.; Dubins, J.; Goldberg, D.R.; De Lombaert, S.; Humbert, M.; Guignabert, C.; Cavasin, M.A.; McKinsey, T.A.; et al. Tryptophan hydroxylase 1 Inhibition Impacts Pulmonary Vascular Remodeling in Two Rat Models of Pulmonary Hypertension. J. Pharmacol. Exp. Ther. 2017, 360, 267–279. [Google Scholar] [CrossRef]
- Biochemistry, Lipopolysaccharide. Available online: https://www.ncbi.nlm.nih.gov/books/NBK554414/ (accessed on 7 November 2023).
- Lawrence, T. The Nuclear Factor NF-κB Pathway in Inflammation. Csh. Perspect. Biol. 2009, 1, a1651. [Google Scholar]
- Soon, E.; Crosby, A.; Southwood, M.; Yang, P.; Tajsic, T.; Toshner, M.; Appleby, S.; Shanahan, C.M.; Bloch, K.D.; Pepke-Zaba, J.; et al. Bone Morphogenetic Protein Receptor Type II Deficiency and Increased Inflammatory Cytokine Production. A Gateway to Pulmonary Arterial Hypertension. Am. J. Respir. Crit. Care Med. 2015, 192, 859–872. [Google Scholar] [CrossRef]
- Galle, J.; Quaschning, T.; Seibold, S.; Wanner, C. Endothelial dysfunction and inflammation: What is the link? Kidney Int. 2003, 63 (Suppl. S84), 45–49. [Google Scholar] [CrossRef]
- Senthong, V.; Wang, Z.; Fan, Y.; Wu, Y.; Hazen, S.L.; Tang, W.W. Trimethylamine N-Oxide and Mortality Risk in Patients With Peripheral Artery Disease. J. Am. Heart Assoc. 2016, 5, e004237. [Google Scholar] [CrossRef]
- Schupack, D.A.; Mars, R.A.T.; Voelker, D.H.; Abeykoon, J.P.; Kashyap, P.C. The Promise of the Gut Microbiome as Part of Indi-vidualized Treatment Strategies. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 7–25. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Miyagawa, K.; Chen, S.-Y.; Tamosiuniene, R.; Wang, L.; Sharpe, O.; Samayoa, E.; Harada, D.; Moonen, J.-R.A.; Cao, A.; et al. Upregulation of Human Endogenous Retrovirus-K Is Linked to Immunity and Inflammation in Pulmonary Arterial Hypertension. Circulation 2017, 136, 1920–1935. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.M.; Edwards, M.R.; Mu, Q.; Yu, Y.; Vieson, M.D.; Reilly, C.M.; Ahmed, S.A.; Bankole, A.A. Gut Microbiota in Human Systemic Lupus Eryth-matosus and a Mouse Model of Lupus. Appl. Environ. Microbiol. 2018, 84, e02288-17. [Google Scholar] [CrossRef] [PubMed]
- Almodovar, S.; Cicalini, S.; Petrosillo, N.; Flores, S.C. Pulmonary hypertension associated with HIV infection: Pulmonary vascular disease: The global perspective. Chest 2010, 137 (Suppl. S6), 6S–12S. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, G.; Tincati, C.; Silvestri, G. Microbial Translocation in the Pathogenesis of HIV Infection and AIDS. Clin. Microbiol. Rev. 2013, 26, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Gori, A.; Tincati, C.; Rizzardini, G.; Torti, C.; Quirino, T.; Haarman, M.; Ben Amor, K.; van Schaik, J.; Vriesema, A.; Knol, J.; et al. Early impairment of gut function and gut flora supporting a role for alteration of gastrointestinal mucosa in human immunodeficiency virus pathogenesis. J. Clin. Microbiol. 2008, 46, 757–758. [Google Scholar] [CrossRef]
- Sibomana, J.P.; Campeche, A.; Carvalho-Filho, R.J.; Correa, R.A.; Duani, H.; Guimaraes, V.P.; Hilton, J.F.; Kassa, B.; Kumar, R.; Lee, M.H.; et al. Schistosomiasis Pulmonary Arterial Hypertension. Front. Immunol. 2020, 11, 608883. [Google Scholar] [CrossRef]
- Kumar, R.; Lee, M.H.; Kassa, B.; Fonseca Balladares, D.C.; Mickael, C.; Sanders, L.; Andruska, A.; Kumar, M.; Spiekerkoetter, E.; Bandeira, A.; et al. Repetitive schistosoma exposure causes perivascular lung fibrosis and persistent pulmonary hypertension. Clin. Sci. 2023, 137, 617–631. [Google Scholar] [CrossRef]
- Jenkins, T.P.; Peachey, L.E.; Ajami, N.J.; MacDonald, A.S.; Hsieh, M.H.; Brindley, P.J.; Cantacessi, C.; Rinaldi, G. Schistosoma mansoniinfection is associated with quantitative and qualitative modifications of the mammalian intestinal microbiota. Sci. Rep. 2018, 8, 12072. [Google Scholar] [CrossRef]
- Floudas, A.; Aviello, G.; Schwartz, C.; Jeffery, I.B.; O’Toole, P.W.; Fallon, P.G. Schistosoma mansoni Worm Infection Regulates the Intestinal Microbiota and Susceptibility to Colitis. Infect. Immun. 2019, 87, 10-1128. [Google Scholar] [CrossRef]
- Bisanz, J.E.; Upadhyay, V.; Turnbaugh, J.A.; Ly, K.; Turnbaugh, P.J. Meta-Analysis Reveals Reproducible Gut Microbiome Alterations in Response to a High-Fat Diet. Cell Host Microbe. 2019, 26, 265–272.e4. [Google Scholar] [CrossRef]
- Singh, R.K.; Chang, H.-W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.-K.; Ivanova, E.A.; Orekhov, A.N. Gut microbiome: A possible common therapeutic target for treatment of atherosclerosis and cancer. Semin. Cancer Biol. 2021, 70, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, T.; Rampelli, S.; Jeffery, I.B.; Santoro, A.; Neto, M.C.; Capri, M.; Giampieri, E.; Jennings, A.; Candela, M.; Turroni, S.; et al. Mediterranean diet intervention alters the gut microbiome in older people reducing frailty and improving health status: The NU-AGE 1-year dietary intervention across five European countries. Gut 2020, 69, 1218–1228. [Google Scholar] [CrossRef] [PubMed]
- Zhan, K.; Gong, X.; Chen, Y.; Jiang, M.; Yang, T.; Zhao, G. Short-Chain Fatty Acids Regulate The immune Responses Via G Protein Coupled Receptor 41 in Bovine Rumen Epithelial Cells. Front. Immunol. 2019, 10, 2042. [Google Scholar] [CrossRef]
- Vinke, P.; Bowen, T.S.; Boekschoten, M.V.; Witkamp, R.F.; Adams, V.; van Norren, K. Anti-inflammatory nutrition with high protein attenuates cardiac and skeletal muscle alterations in a pulmonary arterial hypertension model. Sci. Rep. 2019, 9, 10160. [Google Scholar] [CrossRef]
- Robles-Vera, I.; Callejo, M.; Ramos, R.; Duarte, J.; Perez-Vizcaino, F. Impact of Vitamin D Deficit on the Rat Gut Microbiome. Nutrients. 2019, 11, 2564. [Google Scholar] [CrossRef]
- Yilmaz, B.; Li, H. Gut Microbiota and Iron: The Crucial Actors in Health and Disease. Pharmaceuticals 2018, 11, 98. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Q.; Ma, W.; Tian, F.; Shen, H.; Zhou, M. A combination of quercetin and resveratrol reduces obesity in high-fat diet-fed rats by modulation of gut microbiota. Food Funct. 2017, 8, 4644–4656. [Google Scholar] [CrossRef]
- Hemarajata, P.; Versalovic, J. Effects of probiotics on gut microbiota: Mechanisms of intestinal immunomodulation and neuromodulation. Ther. Adv. Gastroenterol. 2013, 6, 39–51. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, L.; Yang, Z.; Chen, F.; Zhang, Y. Effects of Probiotics on Ghrelin and Lungs in Children with Acute Lung Injury: A Double-Blind Randomized, Controlled Trial. Pediatr. Pulmonol. 2018, 53, 197–203. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov. Available online: https://classic.clinicaltrials.gov/ct2/show/NCT04884971 (accessed on 8 November 2023).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suswał, K.; Tomaszewski, M.; Romaniuk, A.; Świechowska-Starek, P.; Zygmunt, W.; Styczeń, A.; Romaniuk-Suswał, M. Gut–Lung Axis in Focus: Deciphering the Impact of Gut Microbiota on Pulmonary Arterial Hypertension. J. Pers. Med. 2024, 14, 8. https://doi.org/10.3390/jpm14010008
Suswał K, Tomaszewski M, Romaniuk A, Świechowska-Starek P, Zygmunt W, Styczeń A, Romaniuk-Suswał M. Gut–Lung Axis in Focus: Deciphering the Impact of Gut Microbiota on Pulmonary Arterial Hypertension. Journal of Personalized Medicine. 2024; 14(1):8. https://doi.org/10.3390/jpm14010008
Chicago/Turabian StyleSuswał, Konrad, Michał Tomaszewski, Aleksandra Romaniuk, Paulina Świechowska-Starek, Wojciech Zygmunt, Agnieszka Styczeń, and Małgorzata Romaniuk-Suswał. 2024. "Gut–Lung Axis in Focus: Deciphering the Impact of Gut Microbiota on Pulmonary Arterial Hypertension" Journal of Personalized Medicine 14, no. 1: 8. https://doi.org/10.3390/jpm14010008
APA StyleSuswał, K., Tomaszewski, M., Romaniuk, A., Świechowska-Starek, P., Zygmunt, W., Styczeń, A., & Romaniuk-Suswał, M. (2024). Gut–Lung Axis in Focus: Deciphering the Impact of Gut Microbiota on Pulmonary Arterial Hypertension. Journal of Personalized Medicine, 14(1), 8. https://doi.org/10.3390/jpm14010008