
Frequency of CYP2C9 Promoter Variable Number Tandem Repeat Polymorphism in a Spanish Population: Linkage Disequilibrium with CYP2C9*3 Allele

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Determination of CYP2C9 pVNTR
2.3. CYP2C9*2 and *3 Allele Analysis
2.4. Statistical Analysis
3. Results
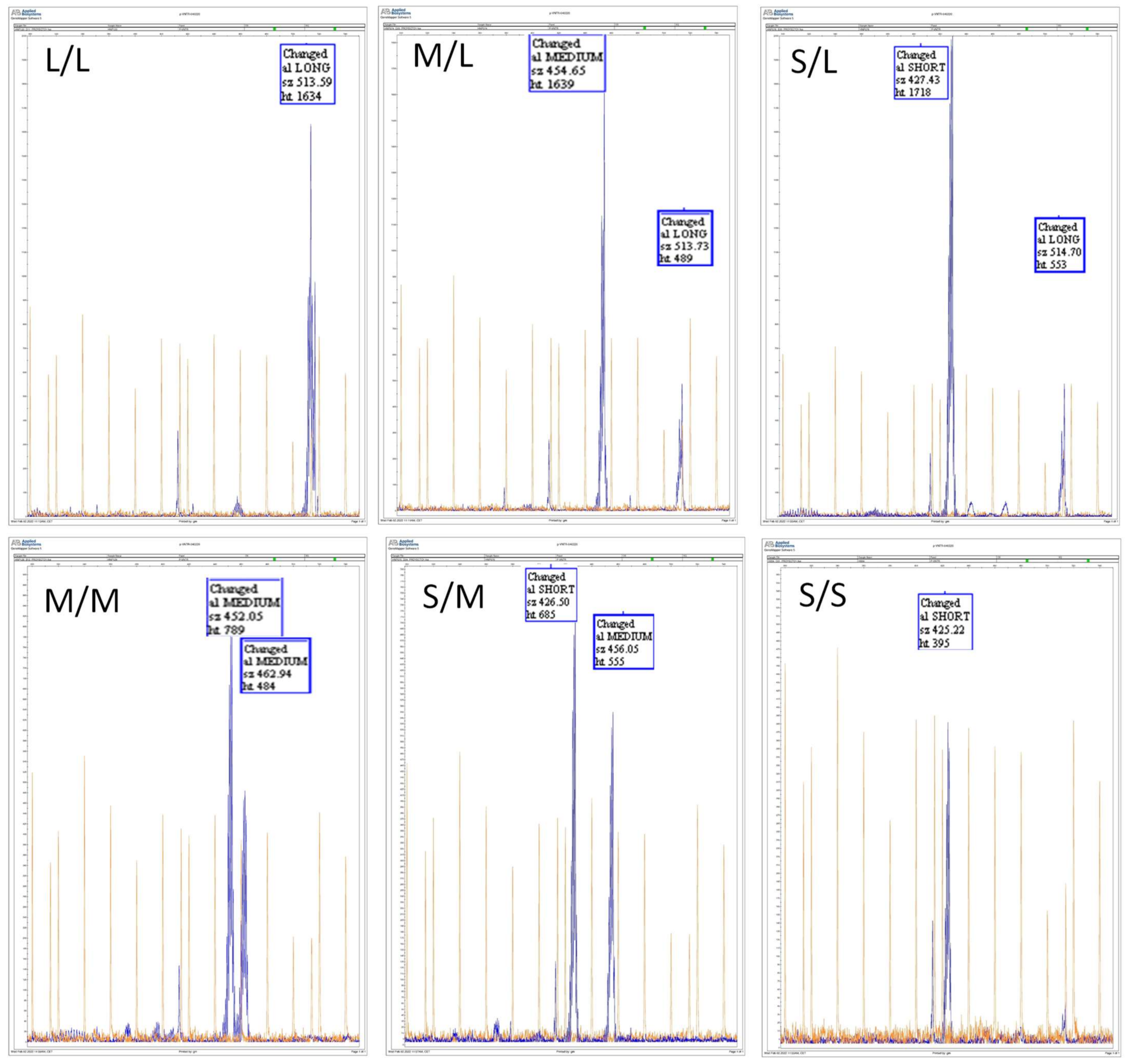
3.1. Determination of the Presence and Frequency of CYP2C9 pVNTR in a Spanish Population
3.2. Comparison of the Frequency of CYP2C9 pVNTR between Different Populations
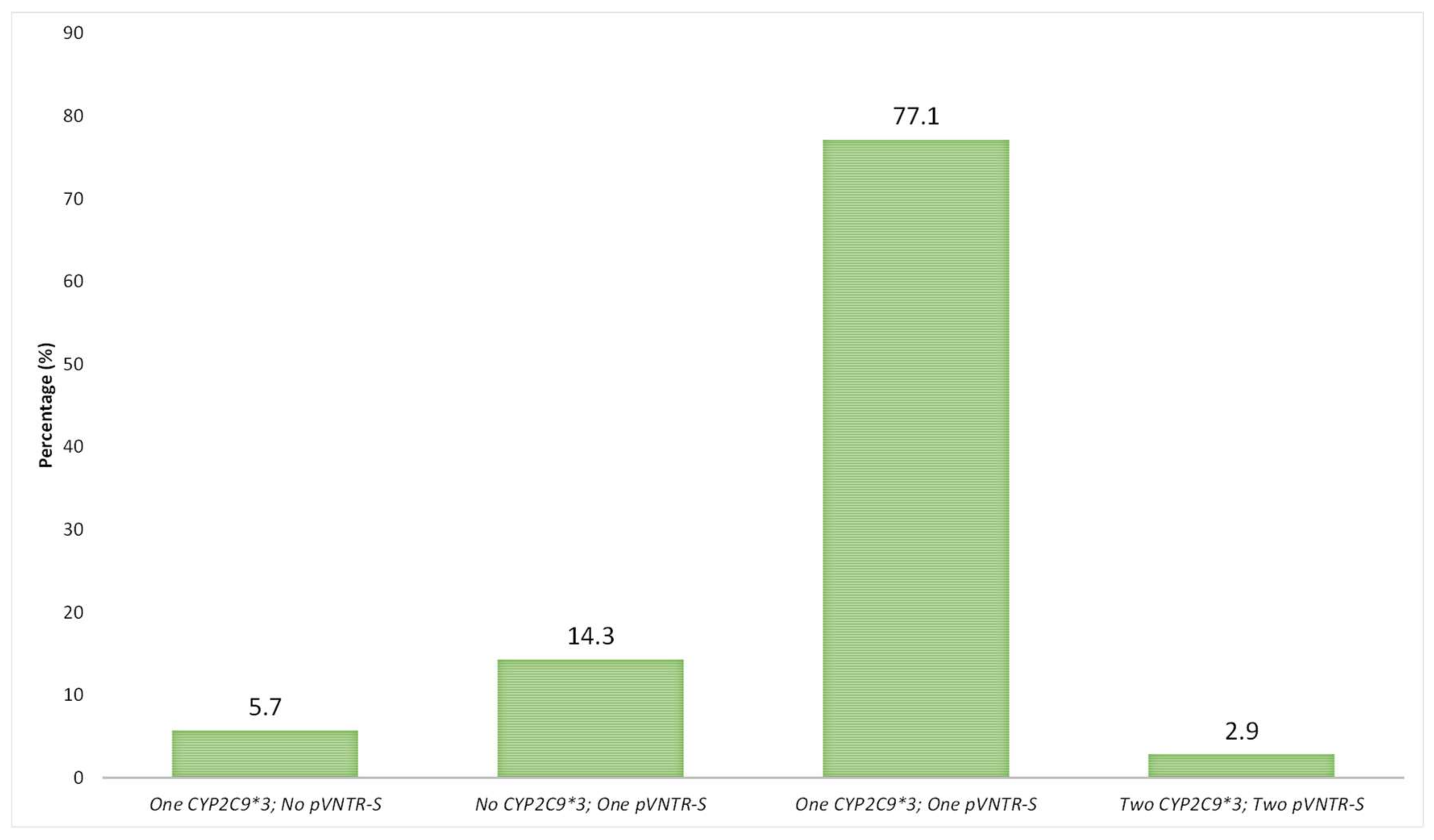
3.3. Analysis of LD between CYP2C9 pVNTR-S and CYP2C9*3 Alleles in a Spanish Population
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, C.R.; Goldstein, J.A.; Pieper, J.A. Cytochrome P450 2C9 polymorphisms: A comprehensive review of the in-vitro and human data. Pharmacogenetics 2002, 12, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Daly, A.K.; Rettie, A.E.; Fowler, D.M.; Miners, J.O. Pharmacogenomics of CYP2C9: Functional and Clinical Considerations. J. Pers. Med. 2017, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, J.A.; de Morais, S.M. Biochemistry and molecular biologyofthe human CYP2C subfamily. Pharmacogenetics 1994, 4, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Soars, M.G.; Gelboin, H.V.; Krausz, K.W.; Riley, R.J. A comparison of relative abundance; activity factor and inhibitory monoclonal antibody approaches in the characterization of human CYP enzymology. Br. J. Clin. Pharmacol. 2003, 55, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Rettie, A.E.; Jones, J.P. Clinical and toxicological relevance of CYP2C9: Drug-druginteractions and pharmacogenetics. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 477–494. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.F.; Zhou, Z.W.; Huang, M. Polymorphisms of human cytochrome P450 2C9 and the functional relevance. Toxicology 2010, 278, 165–188. [Google Scholar] [CrossRef]
- Wang, B.; Wang, J.; Huang, S.Q.; Su, H.H.; Zhou, S.F. Genetic polymorphism of the human cytochrome P450 2C9 gene and its clinical significance. Curr. Drug Metab. 2009, 10, 781–834. [Google Scholar] [CrossRef]
- Westphal, C.; Konkel, A.; Schunck, W.H. CYP-eicosanoids—A new link between omega-3 fatty acids and cardiac disease? Prostaglandins Other Lipid Mediat. 2011, 96, 99–108. [Google Scholar] [CrossRef]
- Spector, A.A.; Kim, H.Y. Cytochrome P450 epoxygenase pathway of polyunsaturated fatty acid metabolism. Biochim. Biophys. Acta 2015, 1851, 356–365. [Google Scholar] [CrossRef] [Green Version]
- The Pharmacogene Variation (PharmVar) Consortium. Available online: https://www.pharmvar.org/gene/CYP2C9 (accessed on 18 February 2022).
- Yasar, U.; Forslund-Bergengren, C.; Tybring, G.; Dorado, P.; Llerena, A.; Sjöqvist, F.; Eliasson, E.; Dahl, M.L. Pharmacokinetics of losartan and its metabolite E-3174 in relation to the CYP2C9 genotype. Clin. Pharmacol. Ther. 2002, 71, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Dorado, P.; Berecz, R.; Norberto, M.J.; Yasar, U.; Dahl, M.L.; LLerena, A. CYP2C9 genotypes and diclofenac metabolism in Spanish healthy volunteers. Eur. J. Clin. Pharmacol. 2003, 59, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Dorado, P.; Beltrán, L.J.; Machín, E.; Peñas-Lledó, E.M.; Terán, E.; Llerena, A. Losartan hydroxylation phenotype in an Ecuadorian population: Influence of CYP2C9 genetic polymorphism, habits and gender. Pharmacogenomics 2012, 13, 1711–1717. [Google Scholar] [CrossRef] [PubMed]
- Kirchheiner, J.; Brockmöller, J. Clinical consequences of cytochrome P450 2C9 polymorphisms. Clin. Pharmacol. Ther. 2005, 77, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Vázquez, A.; Dorado, P.; Fricke-Galindo, I.; Jung-Cook, H.; Monroy-Jaramillo, N.; Martínez-Juárez, I.E.; Familiar-López, I.; Peñas-Lledó, E.; LLerena, A.; López-López, M. CYP2C9; CYP2C19; ABCB1 genetic polymorphisms and phenytoin plasma concentrations in Mexican-Mestizo patients with epilepsy. Pharmacogenom. J. 2016, 16, 286–292. [Google Scholar] [CrossRef]
- Ninomiya, H.; Mamiya, K.; Matsuo, S.; Ieiri, I.; Higuchi, S.; Tashiro, N. Genetic polymorphism of the CYP2C subfamily and excessive serum phenytoin concentration with central nervous system intoxication. Ther. Drug Monit. 2000, 22, 230–232. [Google Scholar] [CrossRef]
- Brandolese, R.; Scordo, M.G.; Spina, E.; Gusella, M.; Padrini, R. Severe phenytoin intoxication in a subject homozygous for CYP2C9*3. Clin. Pharmacol. Ther. 2001, 70, 391–394. [Google Scholar] [CrossRef]
- Kidd, R.S.; Curry, T.B.; Gallagher, S.; Edeki, T.; Blaisdell, J.; Goldstein, J.A. Identification of a null allele of CYP2C9 in an African-American exhibiting toxicity to phenytoin. Pharmacogenetics 2001, 11, 803–808. [Google Scholar] [CrossRef]
- Dorado, P.; López-Torres, E.; Peñas-Lledó, E.M.; Martínez-Antón, J.; Llerena, A. Neurological toxicity after phenytoin infusion in a pediatric patient with epilepsy: Influence of CYP2C9; CYP2C19 and ABCB1 genetic polymorphisms. Pharmacogenom. J. 2013, 13, 359–361. [Google Scholar] [CrossRef]
- Aithal, G.P.; Day, C.P.; Kesteven, P.J.; Daly, A.K. Association of polymorphisms in the cytochrome P450 CYP2C9 with warfarin dose requirement and risk of bleeding complications. Lancet 1999, 353, 717–719. [Google Scholar] [CrossRef]
- Margaglione, M.; Colaizzo, D.; D’Andrea, G.; Brancaccio, V.; Ciampa, A.; Grandone, E.; Di Minno, G. Genetic modulation of oral anticoagulation with warfarin. Thromb. Haemost. 2000, 84, 775–778. [Google Scholar]
- Taube, J.; Halsall, D.; Baglin, T. Influence of cytochrome P-450 CYP2C9 polymorphisms on warfarin sensitivity and risk of over-anticoagulation in patients on long-term treatment. Blood 2000, 96, 1816–1819. [Google Scholar] [CrossRef] [PubMed]
- Verstuyft, C.; Morin, S.; Robert, A.; Loriot, M.A.; Beaune, P.; Jaillon, P.; Becquemont, L. Early acenocoumarol overanticoagulation among cytochrome P450 2C9 poor metabolizers. Pharmacogenetics 2001, 11, 735–737. [Google Scholar] [CrossRef] [PubMed]
- Dreisbach, A.W.; Smith, S.V.; Kyle, P.B.; Ramaiah, M.; Amenuke, M.; Garrett, M.R.; Lirette, S.T.; Griswold, M.E.; Roman, R.J. Urinary CYP eicosanoid excretion correlates with glomerular filtration in African-Americans with chronic kidney disease. Prostaglandins Other Lipid Mediat. 2014, 113–115, 45–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakhtiari, M.; Park, J.; Ding, Y.C.; Shleizer-Burko, S.; Neuhausen, S.L.; Halldórsson, B.V.; Stefánsson, K.; Gymrek, M.; Bafna, V. Variable number tandem repeats mediate the expression of proximal genes. Nat. Commun. 2021, 12, 2075. [Google Scholar] [CrossRef]
- Wang, D.; Sun, X.; Gong, Y.; Gawronski, B.E.; Langaee, T.Y.; Shahin, M.H.; Khalifa, S.I.; Johnson, J.A. CYP2C9 promoter variable number tandem repeat polymorphism regulates mRNA expression in human livers. Drug Metab. Dispos. 2012, 40, 884–891. [Google Scholar] [CrossRef] [Green Version]
- Al-Eitan, L.N.; Almasri, A.Y.; Al-Habahbeh, S.O. Impact of a variable number tandem repeat in the CYP2C9 promoter on warfarin sensitivity and responsiveness in Jordanians with cardiovascular disease. Pharmgenom. Pers. Med. 2019, 21, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Theken, K.N.; Lee, C.R.; Gong, L.; Caudle, K.E.; Formea, C.M.; Gaedigk, A.; Klein, T.E.; Agúndez, J.A.G.; Grosser, T. Clinical Pharmacogenetics Implementation Consortium Guideline (CPIC) for CYP2C9 and Nonsteroidal Anti-Inflammatory Drugs. Clin. Pharmacol. Ther. 2020, 108, 191–200. [Google Scholar] [CrossRef]
- Johnson, J.A.; Caudle, K.E.; Gong, L.; Whirl-Carrillo, M.; Stein, C.M.; Scott, S.A.; Lee, M.T.; Gage, B.F.; Kimmel, S.E.; Perera, M.A.; et al. Clinical Pharmacogenetics Implementation Consortium (CPIC) Guideline for Pharmacogenetics-Guided Warfarin Dosing: 2017 Update. Clin. Pharmacol. Ther. 2017, 102, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Karnes, J.H.; Rettie, A.E.; Somogyi, A.A.; Huddart, R.; Fohner, A.E.; Formea, C.M.; Ta Michael Lee, M.; Llerena, A.; Whirl-Carrillo, M.; Klein, T.E.; et al. Clinical Pharmacogenetics Implementation Consortium (CPIC) Guideline for CYP2C9 and HLA-B Genotypes and Phenytoin Dosing: 2020 Update. Clin. Pharmacol. Ther. 2021, 109, 302–309. [Google Scholar] [CrossRef]
- Cooper-DeHoff, R.M.; Niemi, M.; Ramsey, L.B.; Luzum, J.A.; Tarkiainen, E.K.; Straka, R.J.; Gong, L.; Tuteja, S.; Wilke, R.A.; Wadelius, M.; et al. The Clinical Pharmacogenetics Implementation Consortium (CPIC) guideline for SLCO1B1; ABCG2; and CYP2C9 and statin-associated musculoskeletal symptoms. Clin. Pharmacol. Ther. 2022, 111, 1007–1021. [Google Scholar] [CrossRef]
- PharmGKB. CYP2C9 Frequency Table. Available online: https://api.pharmgkb.org/v1/download/file/attachment/CYP2C9_frequency_table.xlsx (accessed on 18 April 2022).

{kind=link}
{kind=link}
| Nephrology Subjects N = 126 | University Subjects N = 83 | Total Subjects N = 209 | ||||
|---|---|---|---|---|---|---|
| CYP2C9 pVNTR | n | Freq. | n | Freq. | n | Freq. |
| L/L | 3 | 0.024 | 1 | 0.012 | 4 | 0.019 |
| M/L | 19 | 0.151 | 13 | 0.157 | 32 | 0.153 |
| S/L | 2 | 0.016 | 1 | 0.012 | 3 | 0.014 |
| M/M | 83 | 0.659 | 57 | 0.687 | 140 | 0.670 |
| S/M | 19 | 0.151 | 10 | 0.120 | 29 | 0.139 |
| S/S | 0 | 0.000 | 1 | 0.012 | 1 | 0.005 |
| L | 27 | 0.107 | 16 | 0.096 | 43 | 0.103 |
| M | 204 | 0.810 | 137 | 0.825 | 341 | 0.816 |
| S | 21 | 0.083 | 13 | 0.078 | 34 | 0.081 |
| CYP2C9 | n | Freq. | n | Freq. | n | Freq. |
| *1/*1 | 81 | 0.643 | 48 | 0.578 | 129 | 0.617 |
| *1/*2 | 21 | 0.167 | 23 | 0.277 | 44 | 0.211 |
| *1/*3 | 17 | 0.135 | 8 | 0.096 | 25 | 0.120 |
| *2/*2 | 5 | 0.040 | 1 | 0.012 | 6 | 0.029 |
| *2/*3 | 2 | 0.016 | 2 | 0.024 | 4 | 0.019 |
| *3/*3 | 0 | 0.000 | 1 | 0.012 | 1 | 0.005 |
| *1 | 200 | 0.794 | 127 | 0.765 | 327 | 0.782 |
| *2 | 33 | 0.131 | 27 | 0.163 | 60 | 0.144 |
| *3 | 19 | 0.075 | 12 | 0.072 | 31 | 0.074 |
| Population | n | pVNTR-S | pVNTR-M | pVNTR-L | * LD r2 | Ref. |
|---|---|---|---|---|---|---|
| Jordanians | 205 | 0.295 | 0.627 | 0.078 | n.e. | [27] |
| Egyptians | 207 | 0.115 | 0.785 | 0.100 | 0.59 | [26] |
| White Americans | 804 | 0.058 | 0.789 | 0.152 | 0.75 | [26] |
| African Americans | 120 | 0.051 | 0.883 | 0.065 | 0.53 | [26] |
| Spaniards | 209 | 0.081 | 0.816 | 0.103 | 0.88 | Present study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dorado, P.; Santos-Díaz, G.; Gutiérrez-Martín, Y.; Suárez-Santisteban, M.Á. Frequency of CYP2C9 Promoter Variable Number Tandem Repeat Polymorphism in a Spanish Population: Linkage Disequilibrium with CYP2C9*3 Allele. J. Pers. Med. 2022, 12, 782. https://doi.org/10.3390/jpm12050782
Dorado P, Santos-Díaz G, Gutiérrez-Martín Y, Suárez-Santisteban MÁ. Frequency of CYP2C9 Promoter Variable Number Tandem Repeat Polymorphism in a Spanish Population: Linkage Disequilibrium with CYP2C9*3 Allele. Journal of Personalized Medicine. 2022; 12(5):782. https://doi.org/10.3390/jpm12050782
Chicago/Turabian StyleDorado, Pedro, Gracia Santos-Díaz, Yolanda Gutiérrez-Martín, and Miguel Ángel Suárez-Santisteban. 2022. "Frequency of CYP2C9 Promoter Variable Number Tandem Repeat Polymorphism in a Spanish Population: Linkage Disequilibrium with CYP2C9*3 Allele" Journal of Personalized Medicine 12, no. 5: 782. https://doi.org/10.3390/jpm12050782
APA StyleDorado, P., Santos-Díaz, G., Gutiérrez-Martín, Y., & Suárez-Santisteban, M. Á. (2022). Frequency of CYP2C9 Promoter Variable Number Tandem Repeat Polymorphism in a Spanish Population: Linkage Disequilibrium with CYP2C9*3 Allele. Journal of Personalized Medicine, 12(5), 782. https://doi.org/10.3390/jpm12050782