
The Importance of SNPs at miRNA Binding Sites as Biomarkers of Gastric and Colorectal Cancers: A Systematic Review

 ,
,  ,
, 

Abstract
1. Introduction
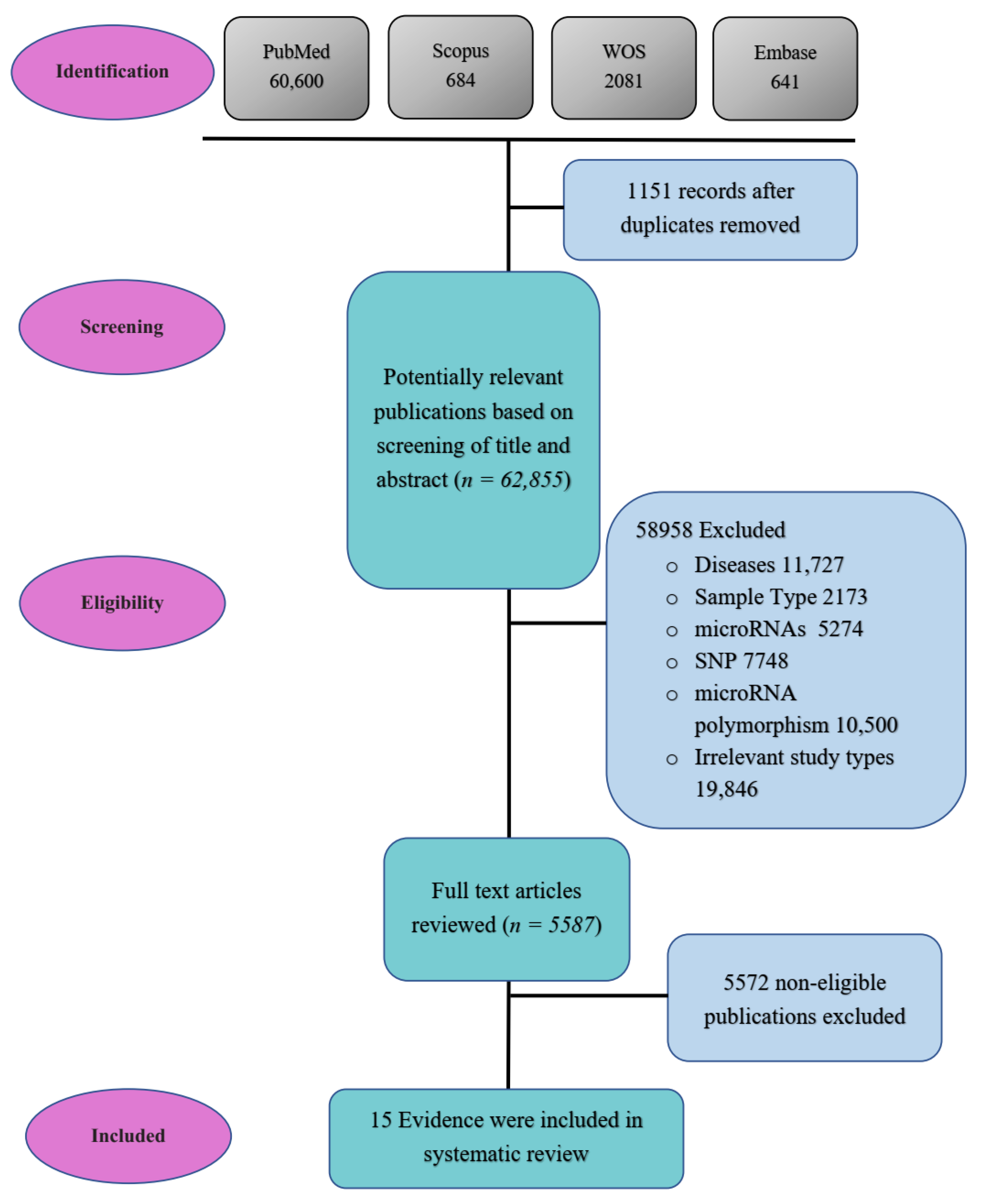
2. Materials and Methods
2.1. Search Strategy
2.2. Study Selection
2.2.1. Inclusion Criteria
2.2.2. Exclusion Criteria
2.2.3. Data Extraction
2.3. Quality Assessment
3. Results
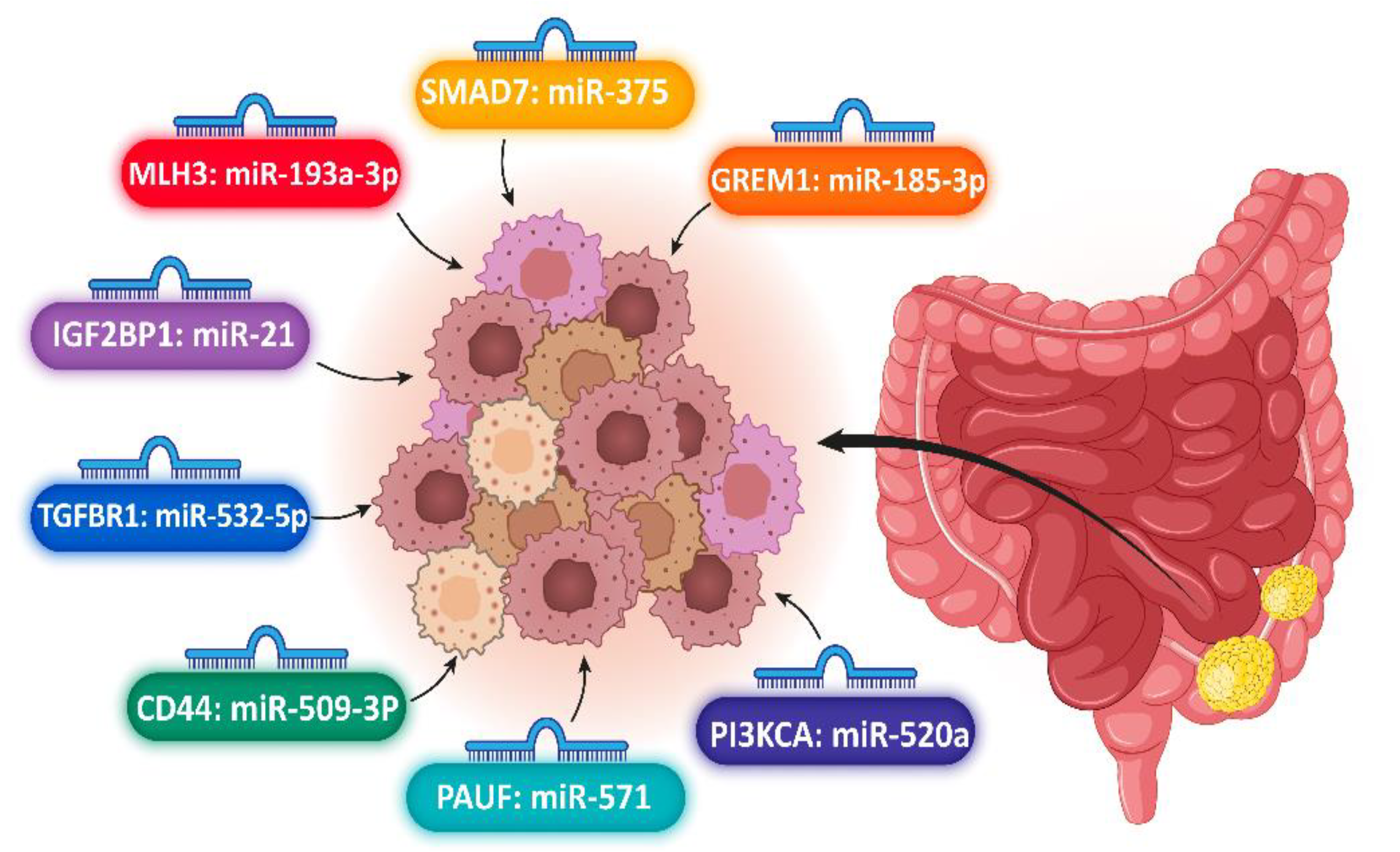
3.1. Colorectal Cancer
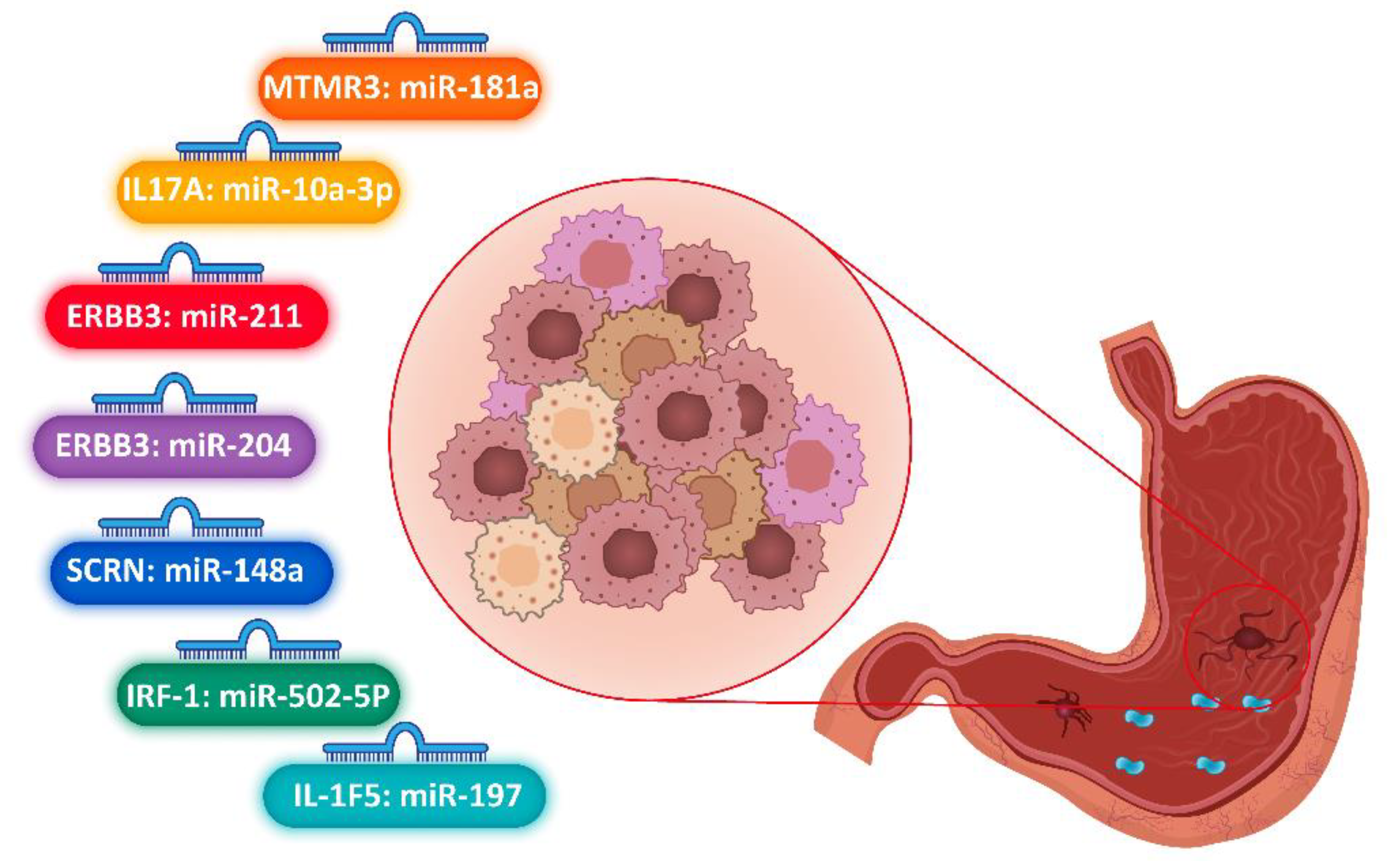
3.2. Gastric Cancer
3.3. Assessment of the Risk of Bias and Quality
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Health Estimates 2019: Deaths by Cause, Age, Sex, by Country and by Region, 2000–2019; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Wu, X.-M.; Yang, H.-G.; Zheng, B.-A.; Cao, H.-F.; Hu, Z.-M.; Wu, W.-D. Functional genetic variations at the microRNA binding-site in the CD44 gene are associated with risk of colorectal cancer in Chinese populations. PLoS ONE 2015, 10, e0127557. [Google Scholar] [CrossRef]
- Lichtenstein, P.; Holm, N.V.; Verkasalo, P.K.; Iliadou, A.; Kaprio, J.; Koskenvuo, M.; Pukkala, E.; Skytthe, A.; Hemminki, K. Environmental and heritable factors in the causation of cancer—Analyses of cohorts of twins from Sweden, Denmark, and Finland. N. Engl. J. Med. 2000, 343, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Rawla, P.; Barsouk, A. Epidemiology of gastric cancer: Global trends, risk factors and prevention. Prz. Gastroenterol. 2019, 14, 26. [Google Scholar] [CrossRef]
- Rawla, P.; Sunkara, T.; Barsouk, A. Epidemiology of colorectal cancer: Incidence, mortality, survival, and risk factors. Prz. Gastroenterol. 2019, 14, 89. [Google Scholar] [CrossRef]
- Huxley, R.R.; Ansary-Moghaddam, A.; Clifton, P.; Czernichow, S.; Parr, C.L.; Woodward, M. The impact of dietary and lifestyle risk factors on risk of colorectal cancer: A quantitative overview of the epidemiological evidence. Int. J. Cancer 2009, 125, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Arends, M.J. Pathways of colorectal carcinogenesis. Appl. Immunohistochem. Mol. Morphol. 2013, 21, 97–102. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Center, M.M.; Jemal, A.; Smith, R.A.; Ward, E. Worldwide variations in colorectal cancer. CA Cancer J. Clin. 2009, 59, 366–378. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, H.; Zou, L.; Ke, J.; Zhang, Y.; Zhu, Y.; Yang, Y.; Gong, Y.; Tian, J.; Zou, D. A functional variant in GREM1 confers risk for colorectal cancer by disrupting a hsa-miR-185-3p binding site. Oncotarget 2017, 8, 61318. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Sood, P.; Krek, A.; Zavolan, M.; Macino, G.; Rajewsky, N. Cell-type-specific signatures of microRNAs on target mRNA expression. Proc. Natl. Acad. Sci. USA 2006, 103, 2746–2751. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a big role in gene regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Annese, T.; Tamma, R.; De Giorgis, M.; Ribatti, D. microRNAs biogenesis, functions and role in tumor angiogenesis. Front. Oncol. 2020, 10, 2610. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.S.; Jiang, W.W.; Smith, I.; Poeta, L.M.; Begum, S.; Glazer, C.; Shan, S.; Westra, W.; Sidransky, D.; Califano, J.A. MicroRNA alterations in head and neck squamous cell carcinoma. Int. J. Cancer 2008, 123, 2791–2797. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Mi, S.; Zhang, W.; Dolan, M.E. Comprehensive analysis of the impact of SNPs and CNVs on human microRNAs and their regulatory genes. RNA Biol. 2009, 6, 412–425. [Google Scholar] [CrossRef]
- Mishra, P.J.; Humeniuk, R.; Mishra, P.J.; Longo-Sorbello, G.S.; Banerjee, D.; Bertino, J.R. A miR-24 microRNA binding-site polymorphism in dihydrofolate reductase gene leads to methotrexate resistance. Proc. Natl. Acad. Sci. USA 2007, 104, 13513–13518. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Jiang, Z.; Chen, Q.; Qin, R.; Fang, Y.; Li, H. A functional variant at miR-520a binding site in PIK3CA alters susceptibility to colorectal cancer in a Chinese Han population. BioMed Res. Int. 2015, 2015, 373252. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.G.; Chae, Y.S.; Lee, S.J.; Kang, B.W.; Park, J.Y.; Lee, E.-J.; Jeon, H.-S.; Park, J.S.; Choi, G.S. Genetic variation in microRNA-binding site and prognosis of patients with colorectal cancer. J. Cancer Res. Clin. Oncol. 2015, 141, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Li, S.; Jin, J.; He, L.; Xu, K.; Zhu, L.; Du, M.; Liu, Y.; Chu, H.; Zhang, Z. Genetic variant in miR-21 binding sites is associated with colorectal cancer risk. J. Cell. Mol. Med. 2019, 23, 2012–2019. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-M.; Lin, C.-C.; Chen, W.-S.; Jiang, J.-K.; Yang, S.-H.; Chang, S.-C.; Ho, C.-L.; Yang, C.-C.; Huang, S.-C.; Chao, Y. Insulin-Like Growth Factor 2 mRNA-Binding Protein 1 (IGF2BP1) Is a Prognostic Biomarker and Associated with Chemotherapy Responsiveness in Colorectal Cancer. Int. J. Mol. Sci. 2021, 22, 6940. [Google Scholar] [CrossRef] [PubMed]
- Peppone, L.J.; Reid, M.E.; Moysich, K.B.; Morrow, G.R.; Jean-Pierre, P.; Mohile, S.G.; Darling, T.V.; Hyland, A. The effect of secondhand smoke exposure on the association between active cigarette smoking and colorectal cancer. Cancer Causes Control 2010, 21, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Gu, D.; Li, S.; Du, M.; Tang, C.; Chu, H.; Tong, N.; Zhang, Z.; Wang, M.; Chen, J. A genetic variant located in the miR-532-5p-binding site of TGFBR1 is associated with the colorectal cancer risk. J. Gastroenterol. 2019, 54, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zheng, X.; Li, X.; Sun, D.; Xue, P.; Zhang, G.; Xiao, M.; Cai, Y.; Jin, C.; Yang, J. The polymorphisms of mi RNA-binding site in MLH 3 and ERCC 1 were linked to the risk of colorectal cancer in a case–control study. Cancer Med. 2018, 7, 1264–1274. [Google Scholar] [CrossRef] [PubMed]
- Shaker, O.G.; Mohammed, S.R.; Mohammed, A.M.; Mahmoud, Z. Impact of micro RNA-375 and its target gene SMAD-7 polymorphism on susceptibility of colorectal cancer. J. Clin. Lab. Anal. 2018, 32, e22215. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liu, Q.; Ren, Y.; Zhang, Y.; Wang, X.; Liu, B. Association analysis of miRNA-related genetic polymorphisms in miR-143/145 and KRAS with colorectal cancer susceptibility and survival. Biosci. Rep. 2021, 41, BSR20204136. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Xu, Y.; Cao, X.; Chen, Y.; Jiang, J.; Wang, K. Associations of Il-1 Family-Related polymorphisms with gastric cancer risk and the role of Mir-197 in Il-1f5 Expression. Medicine 2015, 94, e1982. [Google Scholar] [CrossRef]
- Dong, K.; Xu, Y.; Yang, Q.; Shi, J.; Jiang, J.; Chen, Y.; Song, C.; Wang, K. Associations of functional MicroRNA binding site polymorphisms in IL23/Th17 inflammatory pathway genes with gastric cancer risk. Mediat. Inflamm. 2017, 2017, 6974696. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Nie, Y.; Zhao, J.; Chen, X.; Ye, M.; Li, Y.; Du, Y.; Cao, J.; Shen, B.; Li, Y. Genetic polymorphism at miR-181a binding site contributes to gastric cancer susceptibility. Carcinogenesis 2012, 33, 2377–2383. [Google Scholar] [CrossRef]
- Shi, Y.; Chen, X.; Xi, B.; Yu, X.; Ouyang, J.; Han, C.; Qin, Y.; Wu, D.; Shen, H. SNP rs3202538 in 3’UTR region of ErbB3 regulated by miR-204 and miR-211 promote gastric cancer development in Chinese population. Cancer Cell Int. 2017, 17, 81. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Yang, H.; Shen, L.; Wang, J.; Pu, W.; Chen, Z.; Shen, X.; Fu, J.; Zhuang, Z. Rs56288038 (C/G) in 3’UTR of IRF-1 regulated by MiR-502-5p promotes gastric cancer development. Cell. Physiol. Biochem. 2016, 40, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Zhu, H.; Zhang, D.; Chu, H.; Wu, D.; Kang, M.; Wang, M.; Gong, W.; Zhou, J.; Zhang, Z. A genetic variant of miR-148a binding site in the SCRN1 3’-UTR is associated with susceptibility and prognosis of gastric cancer. Sci. Rep. 2014, 4, 7080. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, Q.-X.; Zhu, Y.-Q.; Zhang, H.; Xiao, J. Altered MiRNA expression in gastric cancer: A systematic review and meta-analysis. Cell. Physiol. Biochem. 2015, 35, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Gholami, M.; Larijani, B.; Sharifi, F.; Hasani-Ranjbar, S.; Taslimi, R.; Bastami, M.; Atlasi, R.; Amoli, M.M. MicroRNA-binding site polymorphisms and risk of colorectal cancer: A systematic review and meta-analysis. Cancer Med. 2019, 8, 7477–7499. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.V.; Galbraith, N.J.; Yang, D.; Burton, J.F.; Walker, S.P.; Galandiuk, S. Blood-based microRNAs as biomarkers for the diagnosis of colorectal cancer: A systematic review and meta-analysis. Br. J. Cancer 2017, 116, 762–774. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Population | Region | Variables | Age | Gender Control | Gender Patient | Stage | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ID | Patient | Control | Country | miRNAs | SNP | Gene | Patient | Control | Male | Female | Male | Female | Low | intermediate | high |
| Xie, 2018 | 1147 | 1203 | China | miR-21 | rs6504593 | IGF2BP1 | 60.0 ± 12.6 | 59.9 ± 14.3 | 698 | 505 | 702 | 445 | 85 | 880 | 182 |
| Gu, 2018 | 1147 | 1203 | China | miR-532-5p | rs1590 | TGFBR1 | 60.0 ± 12.6 | 59.9 ± 14.3 | 698 | 505 | 702 | 445 | 85 | 880 | 182 |
| Zhang, 2017 | 200 | 200 | China | miR-193a-3p | rs10862, rs3212986 | MLH3, ERCC1 | 62.18 ± 12.637 | 61.59 ± 13.153 | 89 | 89 | 111 | 111 | ND | ND | ND |
| Shaker, 2016 | 86 | 36 | Egypt | mir-375 | rs4939827 | SMAD-7 | 50.4 ± 12.4 | 46.8 ± 8.7 | 19 | 17 | 59 | 19 | ND | ND | ND |
| Li, 2017 | 1841 | 1837 | China | hsa-miR-185-3p | rs12915554 | GREM1 | 40 ± 24.8 | 40 ± 26.1 | 1026 | 811 | 1025 | 816 | 1841 | ND | ND |
| Wang, 2021 | 507 | 497 | China | miR-143, miR-145 | rs74693964 | KRAS | 62.55 ± 11.88 | 62.75 ±11.99 | 288 | 207 | 329 | 178 | ND | ND | ND |
| Ding, 2015 | 386 | 394 | China | miR-520a | rs141178472 | PIK3CA | 60.1 ± 12.3 | 60.7 ± 12.9 | 229 | 165 | 216 | 170 | 38 | 308 | 40 |
| Kim, 2015 | 831 | - | South Korean | miR-571 | rs12373, rs3757417T | PAUF | 63 ± 20.3 | ND | ND | ND | 55.3 ± 2.4 | 43.2 ± 1.3 | 150 | 665 | 16 |
| Wu, 2015 | 946 | 989 | China | hsa-mir-509-3p | rs13347, rs10836347 rs11821102G/A | CD44 | 55 ± 24.6 | 58 ± 22.7 | 535 | 454 | 519 | 427 | 84 | 606 | 256 |
| Population | Region | Variables | Age | Gender Control | Gender Patient | Stage | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ID | Patient | Control | Country | miRNAs | SNP | Gene | Patient | Control | Male | Female | Male | Female | Low | intermediate | high |
| Dong, 2017 | 500 | 500 | China | hsa-miR-10a-3p | rs3748067 | IL17A | 57.93 ± 11.88 | 57.27 ± 12.15 | 124 | 376 | 124 | 376 | ND | ND | ND |
| Lin, 2012 | 500 | 502 | China | miR-181a | rs12537 | MTMR3 | 60 ± 13 | 60 ± 15 | 319 | 183 | 314 | 186 | ND | ND | ND |
| Shi, 2017 | 851 | 799 | China | miR-204 miR-211 | rs3202538 | ErbB3 | ≤50 | >50 | 319 | 480 | 318 | 533 | 214 | 294 | 343 |
| Wang, 2016 | 819 | 765 | China | MiR-502-5p | Rs56288038 | IRF-1 | ≤50 | >50 | 319 | 501 | 318 | 480 | 214 | 294 | 311 |
| Chen, 2015 | 500 | 500 | China | miR-197 | rs2472188 | IL1-F5 | 58.03 ± 11.89 | 57.24 ± 12.15 | 376 | 376 | 124 | 124 | ND | ND | ND |
| Song, 2014 | 753 | 949 | China | miR-148a | rs6976789, rs2235749 | SCRN1 | 65 ± 19 | 64 ± 18 | 628 | 321 | 512 | 241 | 222 | 431 | 12 |
| Study ID | Selection | Comparability | Exposure | Score | Quality Assessment | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Xie, 2018 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 8 | 5 |
| Gu, 2018 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | 5 |
| Zhang, 2017 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 7 | 4 |
| Shaker, 2016 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | 5 |
| Li, 2017 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 7 | 4 |
| Wang, 2021 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 7 | 4 |
| Wu, 2015 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 9 | 5 |
| Kim, 2015 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 6 | 3 |
| Ding, 2015 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 1 | 8 | 5 |
| Shi, 2017 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 8 | 5 |
| Dong, 2017 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 7 | 4 |
| Wang, 2016 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 7 | 4 |
| Chen, 2015 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 7 | 4 |
| Peng Song, 2014 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 8 | 5 | |
| Yong Lin, 2012 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 7 | 4 |
| Study ID | Genes | SNPs | miRNAs | Expression miRNAs Pattern in Colorectal Cancer | Expression miRNAs Pattern in Gastric Cancer | Biomarker Categories | Significant Scores |
|---|---|---|---|---|---|---|---|
| Ding, 2015 | PIK3CA | rs141178472-T allele | miR-520a | ↑ | ND | Prognosis | 95% CI: 1.716 (1.084–2.716), p = 0.022 |
| Kim, 2015 | PAUF | rs12373-A allele | miR-571 | ↑ | ND | Prognosis | 95% CI: 1.59 (1.21–2.08), p = 0.0008 |
| Wu, 2015 | CD44 | rs13347-CT and TT alleles | miR-509-3p | ↓ | ND | Prognosis | 95% CI: 1.79 (1.50–2.17), p = 0.004 |
| Xie, 2018 | IGF2BP1 | rs1049109 T allele and rs6504593 | miR-21 | ↓ | ND | Diagnosis | 95% CI: 1.23 (1.07–1.41), p = 0.004 and 95% CI: 1.19 (1.04–1.36), p = 0.011 |
| Gu, 2018 | TGFBR1 | rs1590 GT and GG alleles | miR532-5p | ↓ | ND | Prognosis | 95% CI: 0.82 (0.68–0.97) p = 0.024 |
| Zhang, 2017 | MLH3 | rs108621 CC and TC alleles | miR-193a-3p | ↑ | ND | Prognosis | 95% CI: 6.237 (1.298–29.966), p = 0.013 And 95% CI: 2.079 (1.171–3.691), p = 0.014 |
| Zhang, 2017 | ERCC1 | rs3212986 AA alleles | - | ND | ND | Prognosis | 95% CI: 4.043 (1.261–12.968), p = 0.021 |
| Shaker, 2016 | SMAD7 | rs4939827 T allele | miR-375 | ↓ | ND | Diagnosis | p = 0.01 |
| Li, 2017 | GREM1 | rs12915554 C allele | miR-185-3p | ↓ | ND | Prognosis | 95%CI: 1.43 (1.04–1.95), p = 0.026 |
| Wang, 2021 | KRAS | rs74693964 C and T alleles | miR-145 | ↓ | ND | Prognosis | 95% CI: 1.901 (0.943–3.835), p = 0.001 |
| Dong, 2017 | IL23R | rs10889677 CC | - | ND | - | Diagnosis | 95% CI: 2.22 (1.27–3.87), p = 0.01 |
| IL17A | rs3748067 T, CT and CT + TT alleles | miR-10a-3p | ND | ↑ | Diagnosis | 95% CI: 0.58 (0.43, 0.77), p = 0.01 | |
| Lin, 2012 | MTMR3 | rs12537 CT and TT alleles | miR-181a | ND | ↑ | Prognosis | 95% CI: 1.72 (1.36–2.16), p = 3.99 × 10−5 |
| Shi, 2017 | ERBB3 | rs3202538 GT and TT alleles | miR-204 | ND | ↓ | Diagnosis | 95% CI: 1.89 (1.48–2.01), p = 0.0001 |
| Shi, 2017 | ERBB3 | rs3202538 GT and TT alleles | miR-211 | ND | ↓ | Diagnosis | 95% CI: 4.32 (1.34–1.88), p = 0.0001 |
| Wang, 2016 | IRF-1 | rs56288038 C and G alleles | miR-502-5p | ND | ↑ | Diagnosis | 95% CI: 3.96 (1.52– 2.11), p = 0.0001 |
| Chen, 2015 | IL-1F5 | rs2472188 GC and GC + CC alleles | miR-197 | ND | ↑ | Prognosis | 95% CI: 1.51 (1.15,1.99), p = 0.01 And 1.37 (1.06,1.77), p = 0.01 |
| Chen, 2015 | IL-1F5 | rs2515401 C.T. alleles | miR-197 | ND | ↑ | Prognosis | 95% CI: 1.36 (1.04,1.76), p = 0.01 |
| Song, 2014 | SCRN1 | rs6976789 allele | miR-148a | ND | ↓ | Prognosis | 95% CI: 2.47, (1.21–5.05), p = 0.009 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hajibabaie, F.; Abedpoor, N.; Assareh, N.; Tabatabaiefar, M.A.; Shariati, L.; Zarrabi, A. The Importance of SNPs at miRNA Binding Sites as Biomarkers of Gastric and Colorectal Cancers: A Systematic Review. J. Pers. Med. 2022, 12, 456. https://doi.org/10.3390/jpm12030456
Hajibabaie F, Abedpoor N, Assareh N, Tabatabaiefar MA, Shariati L, Zarrabi A. The Importance of SNPs at miRNA Binding Sites as Biomarkers of Gastric and Colorectal Cancers: A Systematic Review. Journal of Personalized Medicine. 2022; 12(3):456. https://doi.org/10.3390/jpm12030456
Chicago/Turabian StyleHajibabaie, Fatemeh, Navid Abedpoor, Nazanin Assareh, Mohammad Amin Tabatabaiefar, Laleh Shariati, and Ali Zarrabi. 2022. "The Importance of SNPs at miRNA Binding Sites as Biomarkers of Gastric and Colorectal Cancers: A Systematic Review" Journal of Personalized Medicine 12, no. 3: 456. https://doi.org/10.3390/jpm12030456
APA StyleHajibabaie, F., Abedpoor, N., Assareh, N., Tabatabaiefar, M. A., Shariati, L., & Zarrabi, A. (2022). The Importance of SNPs at miRNA Binding Sites as Biomarkers of Gastric and Colorectal Cancers: A Systematic Review. Journal of Personalized Medicine, 12(3), 456. https://doi.org/10.3390/jpm12030456