
Integration of Genomic Risk Scores to Improve the Prediction of Childhood Asthma Diagnosis

 , ,
, ,  and
and
Abstract
1. Introduction
2. Results
2.1. Polygenic Risk Score
2.2. Methylation Risk Scores
2.2.1. Newborn MRS
2.2.2. Childhood MRS
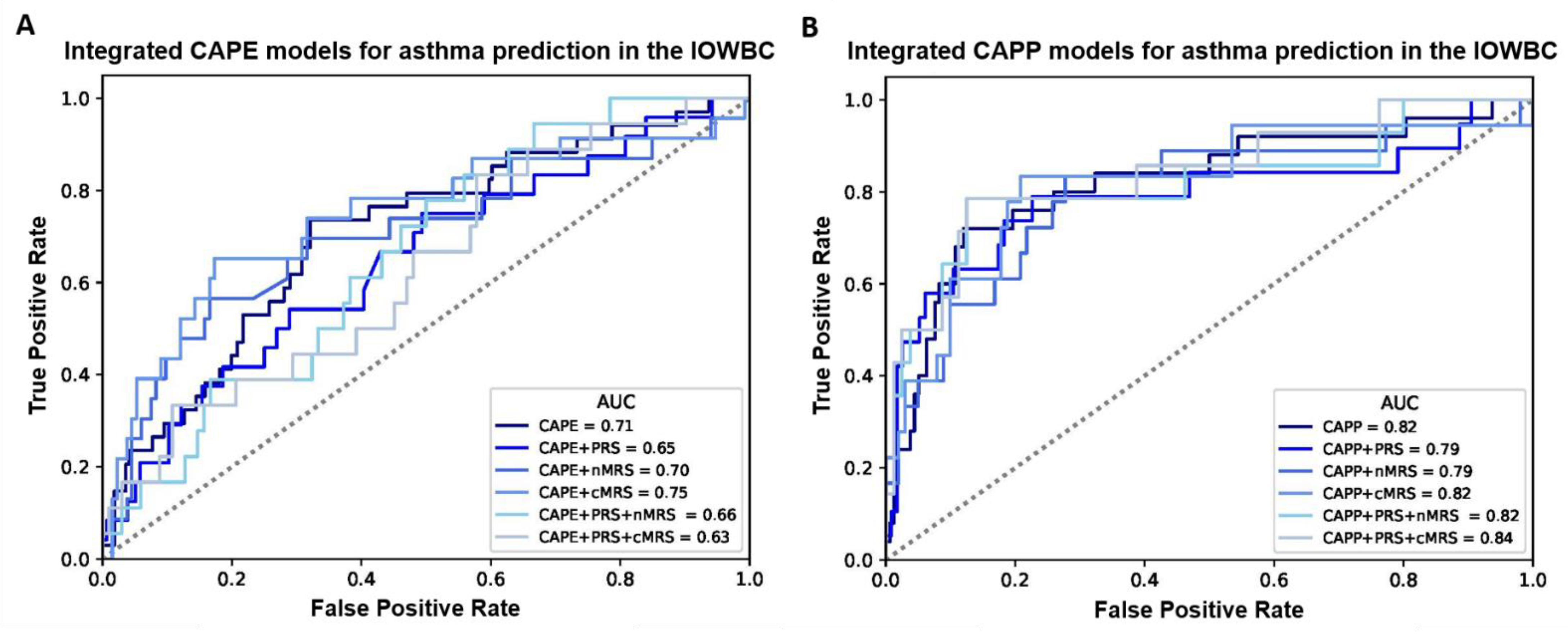
2.3. Integration of Genomic Biomarkers to the CAPE and CAPP Models
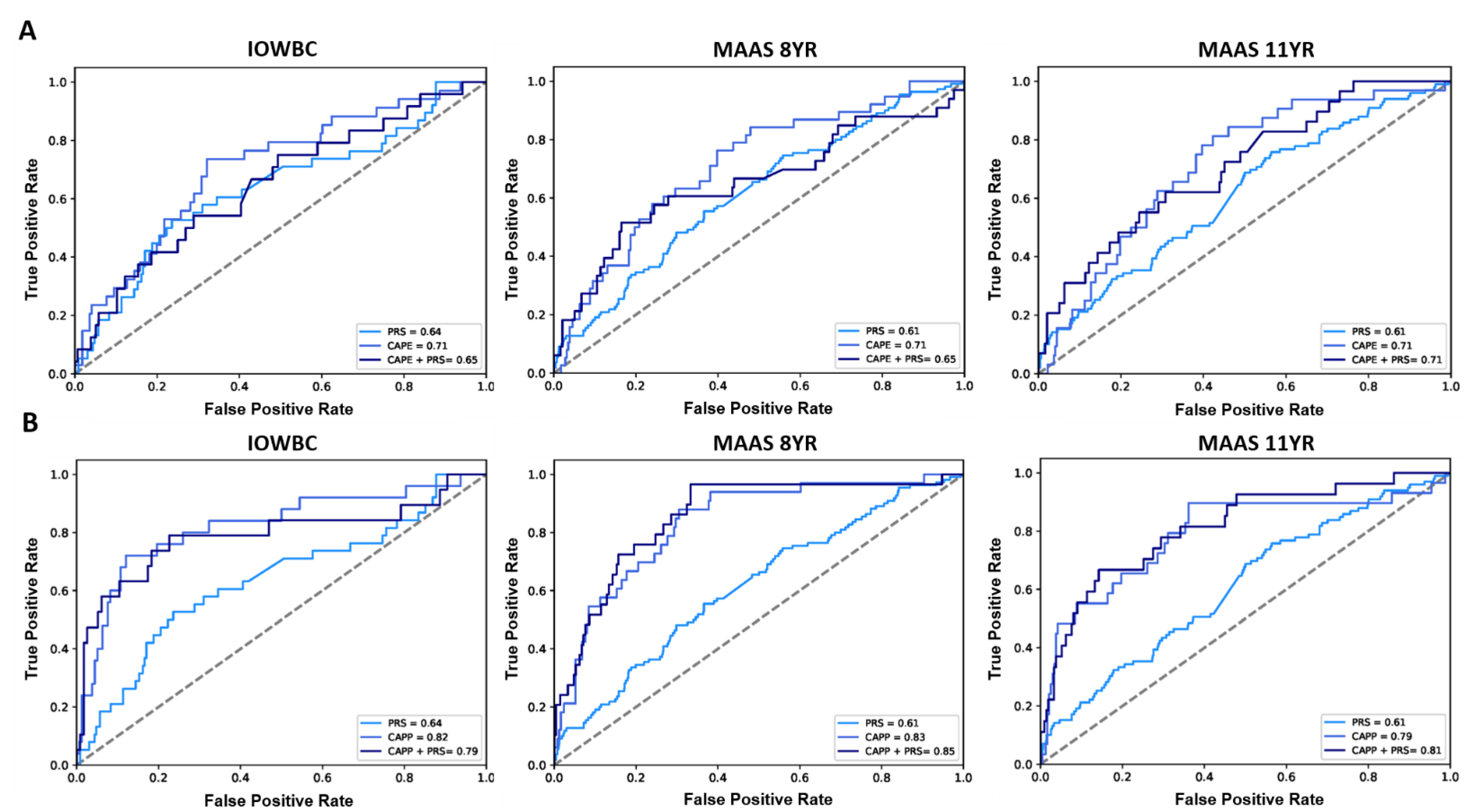
2.4. External Validation
3. Discussion
3.1. Comparison with Existing Studies
3.2. Selection of Genomic Markers
3.2.1. SNPs Included in PRS
3.2.2. CpGs Included in MRS
3.3. Methods for Calculating Genomic Risk Scores
3.4. Integration of Genomic Markers
3.5. Strengths and Limitations
4. Materials and Methods
4.1. Developmental Study Population
4.2. Prediction Outcome
4.3. Construction of the PRS
4.4. Construction of the MRS
4.5. Integration of the Genomic Biomarkers with the CAPE and CAPP Models
4.6. Evaluation of Model Performance
4.7. External Validation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martinez, F.D.; Wright, A.L.; Taussig, L.M.; Holberg, C.J.; Halonen, M.; Morgan, W.J. Asthma and Wheezing in the First Six Years of Life. N. Engl. J. Med. 1995, 332, 133–138. [Google Scholar] [CrossRef]
- Ullmann, N.; Mirra, V.; Di Marco, A.; Pavone, M.; Porcaro, F.; Negro, V.; Onofri, A.; Cutrera, R. Asthma: Differential Diagnosis and Comorbidities. Front. Pediatr. 2018, 6, 276. [Google Scholar] [CrossRef] [PubMed]
- Kothalawala, D.M.; Kadalayil, L.; Weiss, V.B.N.; Kyyaly, M.A.; Arshad, S.H.; Holloway, J.W.; Rezwan, F.I. Prediction models for childhood asthma: A systematic review. Allergy Immunol. 2020, 31, 616–627. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.; Hall, G.L.; Broadhurst, D.; Smith, A.; Schultz, A.; Foong, R.E. Does machine learning have a role in the prediction of asthma in children? Paediatr. Respir. Rev. 2021. [Google Scholar] [CrossRef]
- Owora, A.H.; Tepper, R.S.; Ramsey, C.D.; Becker, A.B.; Genuneit, J. Decision tree-based rules outperform risk scores for childhood asthma prognosis. Pediatr. Allergy Immunol. 2021, 32, 1464–1473. [Google Scholar] [CrossRef]
- Kothalawala, D.M.; Murray, C.S.; Simpson, A.; Custovic, A.; Tapper, W.J.; Arshad, S.H.; Holloway, J.W.; Rezwan, F.I. Development of childhood asthma prediction models using machine learning approaches. Clin. Transl. Allergy 2021, 11, e12076. [Google Scholar] [CrossRef]
- Carr, T.F.; Kraft, M. Use of biomarkers to identify phenotypes and endotypes of severe asthma. Ann. Allergy Asthma Immunol. 2018, 121, 414–420. [Google Scholar] [CrossRef]
- Stephenson, L. Monoclonal Antibody Therapy for Asthma. Clin. Pulm. Med. 2017, 24, 250–257. [Google Scholar] [CrossRef]
- Kuruvilla, M.E.; Lee, F.E.; Lee, G.B. Understanding Asthma Phenotypes, Endotypes, and Mechanisms of Disease. Clin. Rev. Allergy Immunol. 2019, 56, 219–233. [Google Scholar] [CrossRef]
- Ivanova, O.; Richards, L.B.; Vijverberg, S.J.; Neerincx, A.H.; Sinha, A.; Sterk, P.J.; Maitland-van der Zee, A.H. What did we learn from multiple omics studies in asthma? Allergy 2019, 74, 2129–2145. [Google Scholar] [CrossRef]
- Ullemar, V.; Magnusson, P.K.; Lundholm, C.; Zettergren, A.; Melen, E.; Lichtenstein, P.; Almqvist, C. Heritability and confirmation of genetic association studies for childhood asthma in twins. Allergy 2016, 71, 230–238. [Google Scholar] [CrossRef]
- Thomsen, S.F.; Van Der Sluis, S.; Kyvik, K.O.; Skytthe, A.; Backer, V. Estimates of asthma heritability in a large twin sample. Clin. Exp. Allergy 2010, 40, 1054–1061. [Google Scholar] [CrossRef]
- Bracken, M.B.; Belanger, K.; Cookson, W.O.; Triche, E.; Christiani, D.C.; Leaderer, B.P. Genetic and Perinatal Risk Factors for Asthma Onset and Severity: A Review and Theoretical Analysis. Epidemiol. Rev. 2002, 24, 176–189. [Google Scholar] [CrossRef][Green Version]
- Buniello, A.; MacArthur, J.A.L.; Cerezo, M.; Harris, L.W.; Hayhurst, J.; Malangone, C.; McMahon, A.; Morales, J.; Mountjoy, E.; Sollis, E.; et al. The NHGRI-EBI GWAS Catalog of published genome-wide association studies, targeted arrays and summary statistics 2019. Nucleic Acids Res. 2019, 47, D1005–D1012. [Google Scholar] [CrossRef]
- Meyers, D.A.; Bleecker, E.R.; Holloway, J.W.; Holgate, S.T. Asthma genetics and personalised medicine. Lancet Respir. Med. 2014, 2, 405–415. [Google Scholar] [CrossRef]
- Reese, S.E.; Xu, C.J.; den Dekker, H.T.; Lee, M.K.; Sikdar, S.; Ruiz-Arenas, C.; Merid, S.K.; Rezwan, F.I.; Page, C.M.; Ullemar, V.; et al. Epigenome-wide meta-analysis of DNA methylation and childhood asthma. J. Allergy Clin. Immunol. 2019, 143, 2062–2074. [Google Scholar] [CrossRef]
- Edris, A.; den Dekker, H.T.; Melén, E.; Lahousse, L. Epigenome-wide association studies in asthma: A systematic review. Clin. Exp. Allergy 2019, 49, 953–968. [Google Scholar] [CrossRef]
- Qi, C.; Xu, C.-J.; Koppelman, G.H. The role of epigenetics in the development of childhood asthma. Expert Rev. Clin. Immunol. 2019, 15, 1287–1302. [Google Scholar] [CrossRef] [PubMed]
- Belsky, D.W.; Sears, M.R.; Hancox, R.J.; Harrington, H.; Houts, R.; Moffitt, T.E.; Sugden, K.; Williams, B.; Poulton, R.; Caspi, A. Polygenic risk and the development and course of asthma: An analysis of data from a four-decade longitudinal study. Lancet Respir. Med. 2013, 1, 453–461. [Google Scholar] [CrossRef][Green Version]
- Arshad, S.H.; Holloway, J.W.; Karmaus, W.; Zhang, H.; Ewart, S.; Mansfield, L.; Matthews, S.; Hodgekiss, C.; Roberts, G.; Kurukulaaratchy, R. Cohort Profile: The Isle of Wight Whole Population Birth Cohort (IOWBC). Int. J. Epidemiol. 2018, 47, 1043–1044i. [Google Scholar] [CrossRef] [PubMed]
- El-Husseini, Z.W.; Gosens, R.; Dekker, F.; Koppelman, G.H. The genetics of asthma and the promise of genomics-guided drug target discovery. Lancet Respir. Med. 2020, 8, 1045–1056. [Google Scholar] [CrossRef]
- Ferreira, M.A.R.; Mathur, R.; Vonk, J.M.; Szwajda, A.; Brumpton, B.; Granell, R.; Brew, B.K.; Ullemar, V.; Lu, Y.; Jiang, Y.; et al. Genetic Architectures of Childhood- and Adult-Onset Asthma Are Partly Distinct. Am. J. Hum. Genet. 2019, 104, 665–684. [Google Scholar] [CrossRef]
- Clark, H.; Granell, R.; Curtin, J.A.; Belgrave, D.; Simpson, A.; Murray, C.; Henderson, A.J.; Custovic, A.; Paternoster, L. Differential associations of allergic disease genetic variants with developmental profiles of eczema, wheeze and rhinitis. Clin. Exp. Allergy 2019, 49, 1475–1486. [Google Scholar] [CrossRef]
- Park, J.; Jang, H.; Kim, M.; Hong, J.Y.; Kim, Y.H.; Sohn, M.H.; Park, S.-C.; Won, S.; Kim, K.W. Predicting allergic diseases in children using genome-wide association study (GWAS) data and family history. World Allergy Organ. J. 2021, 14, 100539. [Google Scholar] [CrossRef] [PubMed]
- Simard, M.; Madore, A.-M.; Girard, S.; Waserman, S.; Duan, Q.; Subbarao, P.; Sears, M.R.; Moraes, T.J.; Becker, A.B.; Turvey, S.E.; et al. Polygenic risk score for atopic dermatitis in the Canadian population. J. Allergy Clin. Immunol. 2021, 147, 406–409. [Google Scholar] [CrossRef] [PubMed]
- Sordillo, J.E.; Lutz, S.M.; McGeachie, M.J.; Lasky-Su, J.; Weiss, S.T.; Celedón, J.C.; Wu, A.C. Pharmacogenetic Polygenic Risk Score for Bronchodilator Response in Children and Adolescents with Asthma: Proof-of-Concept. J. Pers. Med. 2021, 11, 319. [Google Scholar] [CrossRef]
- Spycher, B.D.; Henderson, J.; Granell, R.; Evans, D.M.; Smith, G.D.; Timpson, N.J.; Sterne, J.A.C. Genome-wide prediction of childhood asthma and related phenotypes in a longitudinal birth cohort. J. Allergy Clin. Immunol. 2012, 130, 503–509.e7. [Google Scholar] [CrossRef]
- Hüls, A.; Czamara, D. Methodological challenges in constructing DNA methylation risk scores. Epigenetics 2020, 15, 1–11. [Google Scholar] [CrossRef]
- Horvath, S.; Raj, K. DNA methylation-based biomarkers and the epigenetic clock theory of ageing. Nat. Rev. Genet. 2018, 19, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Jylhävä, J.; Pedersen, N.L.; Hägg, S. Biological Age Predictors. EBioMedicine 2017, 21, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Sugden, K.; Hannon, E.J.; Arseneault, L.; Belsky, D.W.; Broadbent, J.M.; Corcoran, D.L.; Hancox, R.J.; Houts, R.M.; Moffitt, T.E.; Poulton, R.; et al. Establishing a generalized polyepigenetic biomarker for tobacco smoking. Transl. Psychiatry 2019, 9, 92. [Google Scholar] [CrossRef] [PubMed]
- Bollepalli, S.; Korhonen, T.; Kaprio, J.; Anders, S.; Ollikainen, M. EpiSmokEr: A robust classifier to determine smoking status from DNA methylation data. Epigenomics 2019, 11, 1469–1486. [Google Scholar] [CrossRef]
- Odintsova, V.V.; Rebattu, V.; Hagenbeek, F.A.; Pool, R.; Beck, J.J.; Ehli, E.A.; van Beijsterveldt, C.E.M.; Ligthart, L.; Willemsen, G.; de Geus, E.J.C.; et al. Predicting Complex Traits and Exposures From Polygenic Scores and Blood and Buccal DNA Methylation Profiles. Front. Psychiatry 2021, 12, 688464. [Google Scholar] [CrossRef]
- Hamilton, O.K.L.; Zhang, Q.; McRae, A.F.; Walker, R.M.; Morris, S.W.; Redmond, P.; Campbell, A.; Murray, A.D.; Porteous, D.J.; Evans, K.L.; et al. An epigenetic score for BMI based on DNA methylation correlates with poor physical health and major disease in the Lothian Birth Cohort. Int. J. Obes. 2019, 43, 1795–1802. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Raut, J.R.; Schöttker, B.; Holleczek, B.; Zhang, Y.; Brenner, H. Individual and joint contributions of genetic and methylation risk scores for enhancing lung cancer risk stratification: Data from a population-based cohort in Germany. Clin. Epigenet. 2020, 12, 89. [Google Scholar] [CrossRef] [PubMed]
- Dijk, F.N.; Folkersma, C.; Gruzieva, O.; Kumar, A.; Wijga, A.H.; Gehring, U.; Kull, I.; Postma, D.S.; Vonk, J.M.; Melen, E.; et al. Genetic risk scores do not improve asthma prediction in childhood. J. Allergy Clin. Immunol. 2019, 144, 857–860.e7. [Google Scholar] [CrossRef]
- Jiang, Y.; Wei, J.; Zhang, H.; Ewart, S.; Rezwan, F.I.; Holloway, J.W.; Arshad, H.; Karmaus, W. Epigenome wide comparison of DNA methylation profile between paired umbilical cord blood and neonatal blood on Guthrie cards. Epigenetics 2020, 15, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Lewis, C.M.; Vassos, E. Polygenic risk scores: From research tools to clinical instruments. Genome Med. 2020, 12, 44. [Google Scholar] [CrossRef]
- Igartua, C.; Myers, R.A.; Mathias, R.A.; Pino-Yanes, M.; Eng, C.; Graves, P.E.; Levin, A.M.; Del-Rio-Navarro, B.E.; Jackson, D.J.; Livne, O.E.; et al. Ethnic-specific associations of rare and low-frequency DNA sequence variants with asthma. Nat. Commun. 2015, 6, 5965. [Google Scholar] [CrossRef]
- Mukherjee, N.; Arathimos, R.; Chen, S.; Kheirkhah Rahimabad, P.; Han, L.; Zhang, H.; Holloway, J.W.; Relton, C.; Henderson, A.J.; Arshad, S.H.; et al. DNA methylation at birth is associated with lung function development until age 26 years. Eur. Respir. J. 2021, 57, 2003505. [Google Scholar] [CrossRef]
- Choi, S.W.; Mak, T.S.; O’Reilly, P.F. Tutorial: A guide to performing polygenic risk score analyses. Nat. Protoc. 2020, 15, 2759–2772. [Google Scholar] [CrossRef] [PubMed]
- Custovic, A.; Simpson, B.M.; Murray, C.S.; Lowe, L.; Woodcock, A. The National Asthma Campaign Manchester Asthma and Allergy Study. Pediatr. Allergy Immunol. 2002, 13, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, D.; Zhou, X.; Li, B.; Zhang, Y.; Cheung, W.K.; Lu, A.; Zhang, L. Editorial: Predicting High-Risk Individuals for Common Diseases Using Multi-Omics and Epidemiological Data. Front. Genet. 2021, 12, 737598. [Google Scholar] [CrossRef]
- Morales, E.; Duffy, D. Genetics and Gene-Environment Interactions in Childhood and Adult Onset Asthma. Front. Pediatr. 2019, 7, 499. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| MRS Method † | Newborn MRS: 6 CpGs AUC (95% CI) | Childhood MRS: 110 CpGs AUC (95% CI) |
|---|---|---|
| 1 | 0.55 (0.50–0.60) ‡ | 0.53 (0.48–0.59) |
| 2 | 0.54 (0.48, 0.59) | 0.54 (0.49, 0.59) ‡ |
| 3 | 0.49 (0.44, 0.55) | 0.53 (0.48, 0.59) |
| 4 | 0.53 (0.48, 0.59) | 0.53 (0.48, 0.59) |
| 5 | 0.52 (0.47, 0.58) | 0.53 (0.48, 0.59) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kothalawala, D.M.; Kadalayil, L.; Curtin, J.A.; Murray, C.S.; Simpson, A.; Custovic, A.; Tapper, W.J.; Arshad, S.H.; Rezwan, F.I.; Holloway, J.W.; et al. Integration of Genomic Risk Scores to Improve the Prediction of Childhood Asthma Diagnosis. J. Pers. Med. 2022, 12, 75. https://doi.org/10.3390/jpm12010075
Kothalawala DM, Kadalayil L, Curtin JA, Murray CS, Simpson A, Custovic A, Tapper WJ, Arshad SH, Rezwan FI, Holloway JW, et al. Integration of Genomic Risk Scores to Improve the Prediction of Childhood Asthma Diagnosis. Journal of Personalized Medicine. 2022; 12(1):75. https://doi.org/10.3390/jpm12010075
Chicago/Turabian StyleKothalawala, Dilini M., Latha Kadalayil, John A. Curtin, Clare S. Murray, Angela Simpson, Adnan Custovic, William J. Tapper, S. Hasan Arshad, Faisal I. Rezwan, John W. Holloway, and et al. 2022. "Integration of Genomic Risk Scores to Improve the Prediction of Childhood Asthma Diagnosis" Journal of Personalized Medicine 12, no. 1: 75. https://doi.org/10.3390/jpm12010075
APA StyleKothalawala, D. M., Kadalayil, L., Curtin, J. A., Murray, C. S., Simpson, A., Custovic, A., Tapper, W. J., Arshad, S. H., Rezwan, F. I., Holloway, J. W., & on behalf of STELAR/UNICORN investigators. (2022). Integration of Genomic Risk Scores to Improve the Prediction of Childhood Asthma Diagnosis. Journal of Personalized Medicine, 12(1), 75. https://doi.org/10.3390/jpm12010075