
Modulation of the Dental Pulp Stem Cell Secretory Profile by Hypoxia Induction Using Cobalt Chloride

 , , ,
, , ,  , , and
, , and 
Abstract

1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Isolation and Culture of DPSCs
2.3. Characterization Using Flow Cytometry
2.4. 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide (MTT) Assay
2.5. Cell Proliferation Assay CCK-8 (Cell Counting Kit 8 Assay)
2.6. Quantitative Real-Time PCR for Analysis of Gene Expression
2.7. Cytometric Bead Array for the Detection of Cytokines and Growth Factors
2.8. Statistical Analysis
3. Results
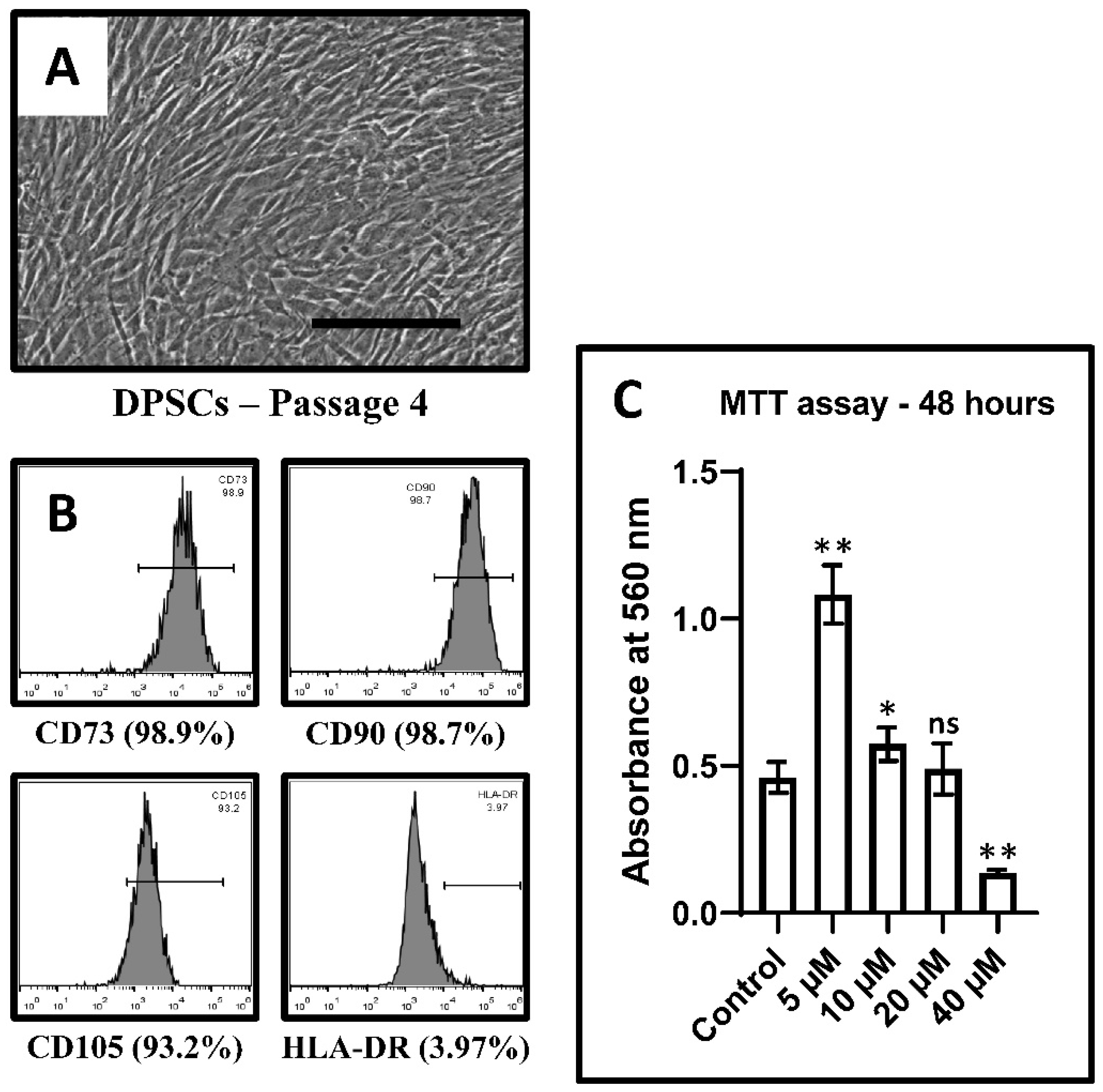
3.1. DPSC Characterization
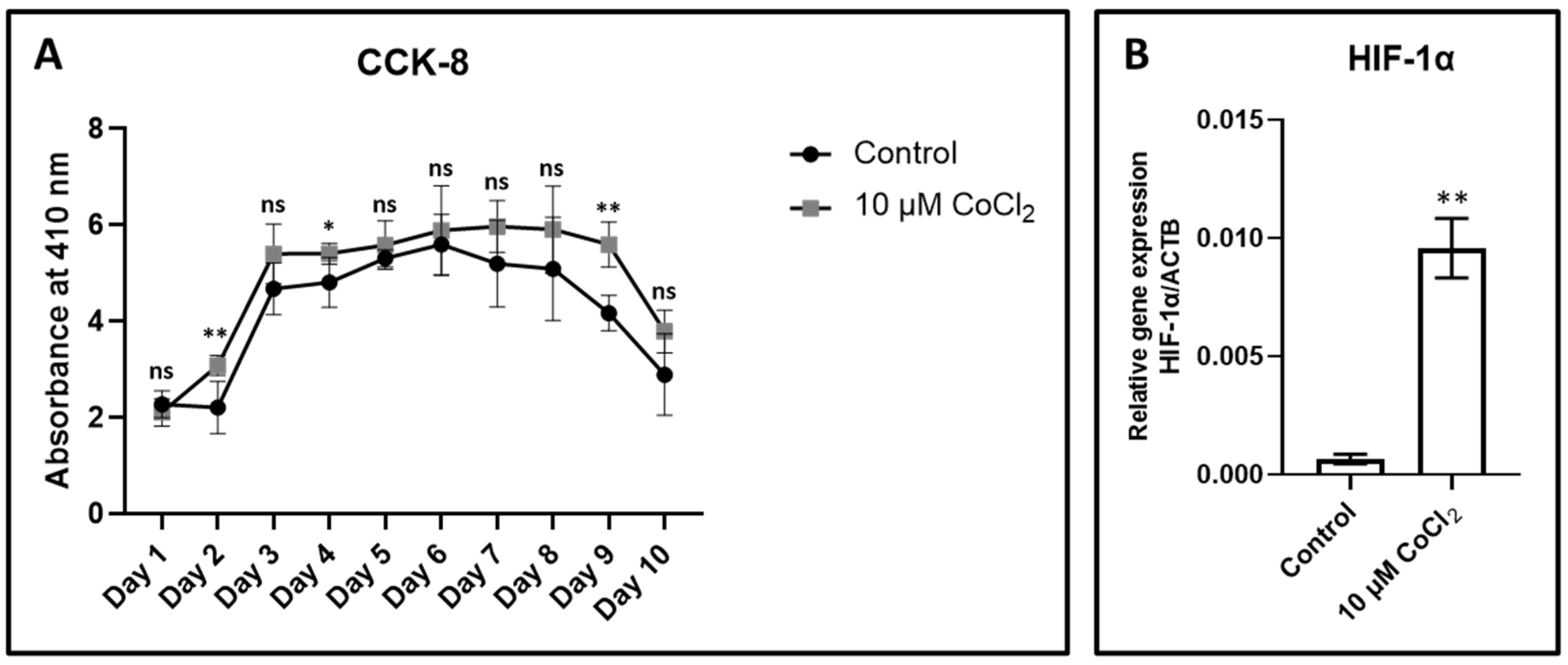
3.2. Effect of CoCl2 on Proliferation and Viability
3.3. Expression of Hypoxia-Inducing Gene HIF-1α
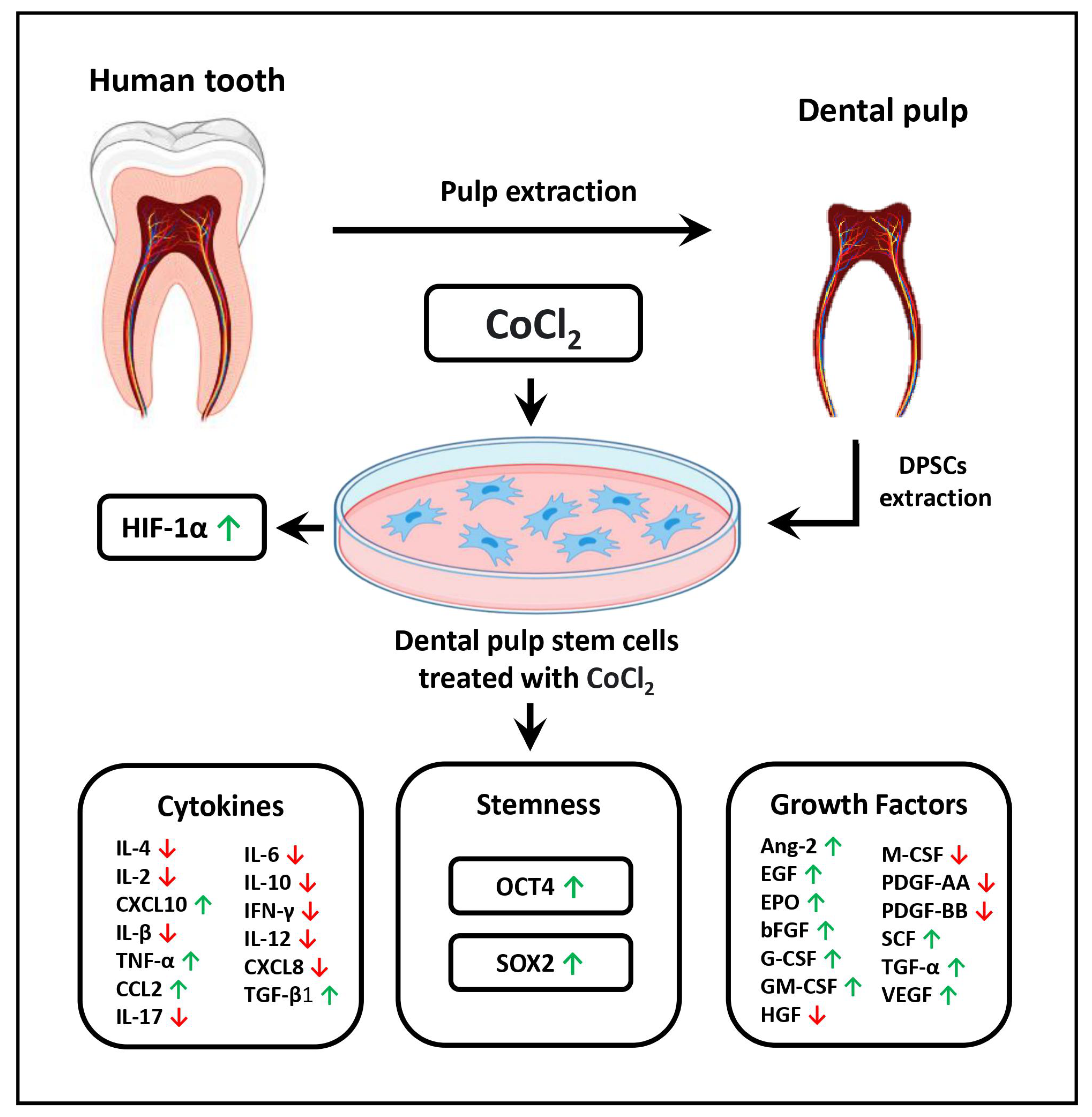
3.4. Modulation of the Secretory Profile of DPSCs in Terms of Growth Factors and Cytokines
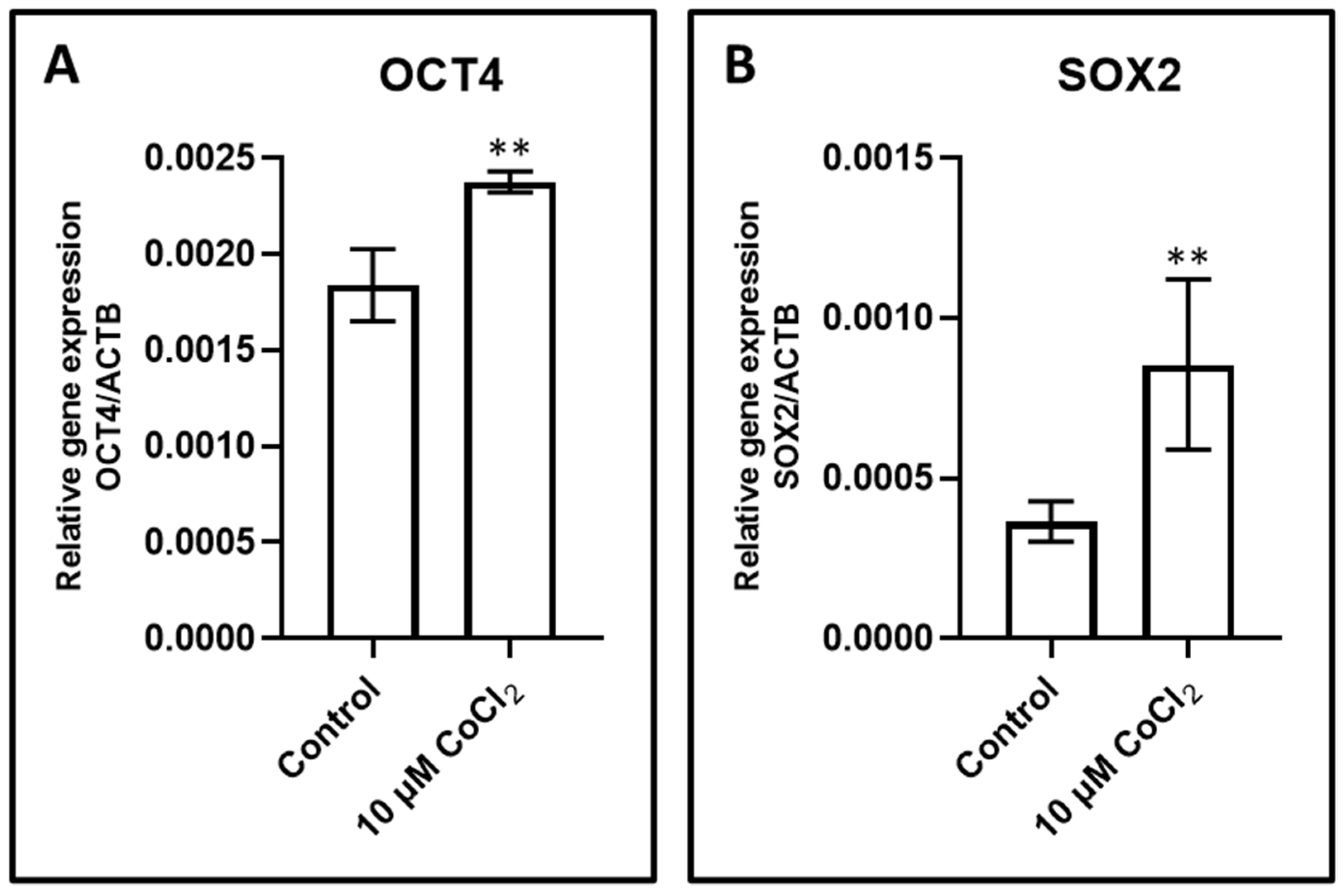
3.5. Effect of CoCl2 Induced Hypoxia on Gene Expression of OCT4 and SOX2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [PubMed]
- Kerkis, I.; Kerkis, A.; Dozortsev, D.; Stukart-Parsons, G.C.; Gomes Massironi, S.M.; Pereira, L.V.; Caplan, A.I.; Cerruti, H.F. Isolation and characterization of a population of immature dental pulp stem cells expressing OCT-4 and other embryonic stem cell markers. Cells Tissues Organs 2007, 184, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.E.M.B.; Murakami, M.; Kaneko, S.; Nakashima, M. The effects of hypoxia on the stemness properties of human dental pulp stem cells (DPSCs). Sci. Rep. 2016, 6, 35476. [Google Scholar] [CrossRef] [PubMed]
- Gronthos, S.; Brahim, J.; Li, W.; Fisher, L.W.; Cherman, N.; Boyde, A.; Denbesten, P.; Robey, P.G.; Shi, S. Stem Cell Properties of human dental pulp stem cells. J. Dent. Res. 2002, 81, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Walboomers, X.F.; Shi, S.; Fan, M.; Jansen, J.A. Multilineage differentiation potential of stem cells derived from human dental pulp after cryopreservation. Tissue Eng. 2006, 12, 2813–2823. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Huang, G.W.; Shiung, J.N.; Huang, Y.H.; Jeng, J.H.; Kuo, T.F.; Yang, J.C.; Yang, W.C.V. Magnetic cryopreservation for dental pulp stem cells. Cells Tissues Organs 2012, 196, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Bhattacharyya, S.; Rattan, V. Effect of uncontrolled freezing on biological characteristics of human dental pulp stem cells. Cell Tissue Bank. 2015, 16, 513–522. [Google Scholar] [CrossRef]
- Yang, X.; Li, L.; Xiao, L.; Zhang, D. Recycle the dental fairy’s package: Overview of dental pulp stem cells. Stem Cell Res. Ther. 2018, 9, 347. [Google Scholar] [CrossRef]
- Salehi, H.; Al-Arag, S.; Middendorp, E.; Gergely, C.; Cuisinier, F.; Orti, V. Dental pulp stem cells used to deliver the anticancer drug paclitaxel. Stem Cell Res. Ther. 2018, 9, 103. [Google Scholar] [CrossRef]
- Gao, J.; Dennis, J.E.; Muzic, R.F.; Lundberg, M.; Caplan, A.I. The dynamic in vivo distribution of bone marrow-derived MSC after infusion.pdf. Cells Tissue Organs 2001, 7080, 12–20. [Google Scholar] [CrossRef]
- Cunningham, C.J.; Redondo-Castro, E.; Allan, S.M. The therapeutic potential of the mesenchymal stem cell secretome in ischaemic stroke. J. Cereb. Blood Flow Metab. 2018, 38, 1276–1292. [Google Scholar] [CrossRef] [PubMed]
- Vizoso, F.J.; Eiro, N.; Cid, S.; Schneider, J.; Perez-Fernandez, R. Mesenchymal stem cell secretome: Toward cell-free therapeutic strategies in regenerative medicine. Int. J. Mol. Sci. 2017, 18, 1852. [Google Scholar] [CrossRef] [PubMed]
- Paschalidis, T.; Bakopoulou, A.; Papa, P.; Leyhausen, G.; Geurtsen, W.; Koidis, P. Dental pulp stem cells’ secretome enhances pulp repair processes and compensates TEGDMA-induced cytotoxicity. Dent. Mater. 2014, 30, e405–e418. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, N.; Kano, F.; Hashimoto, N.; Mori, H.; Liu, Y.; Xia, L.; Sakamaki, T.; Hibi, H.; Iwamoto, T.; Tanaka, E.; et al. Factors secreted from dental pulp stem cells show multifaceted benefits for treating experimental temporomandibular joint osteoarthritis. Osteoarthr. Cartil. 2020, 28, 831–841. [Google Scholar] [CrossRef]
- Spencer, J.A.; Ferraro, F.; Roussakis, E.; Klein, A.; Runnels, J.M.; Zaher, W.; Mortensen, L.J.; Alt, C.; Yusuf, R.; Côté, D.; et al. Direct measurement of local oxygen concentration in the bone marrow of live animals. Nature 2014, 508, 269–273. [Google Scholar] [CrossRef]
- Lee, J.H.; Yoon, Y.M.; Lee, S.H. Hypoxic preconditioning promotes the bioactivities of mesenchymal stem cells via the HIF-1α-GRP78-Akt axis. Int. J. Mol. Sci. 2017, 18, 1320. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.Y.; Boyd, N.M.; Cringle, S.J.; Alder, V.A.; Yu, D.Y. Oxygen distribution and consumption in rat lower incisor pulp. Arch. Oral Boil. 2002, 47, 529–536. [Google Scholar] [CrossRef]
- Kozam, G. Oxygen Tension of Rabbit Incisor Pulp. J. Dent. Res. 1697, 46, 352–358. [Google Scholar] [CrossRef]
- Huang, Y.C.; Parolini, O.; Deng, L.; Yu, B.S. Should hypoxia preconditioning become the standardized procedure for bone marrow MSCs preparation for clinical use? Stem Cells 2016, 34, 1992–1993. [Google Scholar] [CrossRef]
- Zeng, H.L.; Zhong, Q.; Qin, Y.L.; Bu, Q.Q.; Han, X.A.; Jia, H.T.; Liu, H.W. Hypoxia-mimetic agents inhibit proliferation and alter the morphology of human umbilical cord-derived mesenchymal stem cells. BMC Cell Biol. 2011, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Sánchez, J.; Chánez-Cárdenas, M.E. The use of cobalt chloride as a chemical hypoxia model. J. Appl. Toxicol. 2019, 39, 556–570. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. Life with oxygen. Science 2007, 318, 62–64. [Google Scholar] [CrossRef] [PubMed]
- Laksana, K.; Sooampon, S.; Pavasant, P.; Sriarj, W. Cobalt Chloride Enhances the Stemness of Human Dental Pulp Cells. J. Endod. 2017, 43, 760–765. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, Q.; Yang, X.; Yu, X.; Yu, D.; Zhao, W. Effects of cobalt chloride on the stem cell marker expression and osteogenic differentiation of stem cells from human exfoliated deciduous teeth. Cell Stress Chaperones 2019, 24, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Osathanon, T.; Vivatbutsiri, P.; Sukarawan, W.; Sriarj, W.; Pavasant, P.; Sooampon, S. Cobalt chloride supplementation induces stem-cell marker expression and inhibits osteoblastic differentiation in human periodontal ligament cells. Arch. Oral Biol. 2015, 60, 29–36. [Google Scholar] [CrossRef]
- Patil, V.R.; Kharat, A.H.; Kulkarni, D.G.; Kheur, S.M.; Bhonde, R.R. Long term explant culture for harvesting homogeneous population of human dental pulp stem cells. Cell Biol. Int. 2018, 42, 1602–1610. [Google Scholar] [CrossRef]
- Lei, M.; Li, K.; Li, B.; Gao, L.-N.; Chen, F.-M.; Jin, Y. Mesenchymal stem cell characteristics of dental pulp and periodontal ligament stem cells after in vivo transplantation. Biomaterials 2014, 35, 6332–6343. [Google Scholar] [CrossRef]
- Kwon, S.Y.; Chun, S.Y.; Ha, Y.S.; Kim, D.H.; Kim, J.; Song, P.H.; Kim, H.T.; Yoo, E.S.; Kim, B.S.; Kwon, T.G. Hypoxia Enhances Cell Properties of Human Mesenchymal Stem Cells. Tissue Eng. Regen. Med. 2017, 14, 595–604. [Google Scholar] [CrossRef]
- Hung, S.P.; Ho, J.H.; Shih, Y.R.V.; Lo, T.; Lee, O.K. Hypoxia promotes proliferation and osteogenic differentiation potentials of human mesenchymal stem cells. J. Orthop. Res. 2012, 30, 260–266. [Google Scholar] [CrossRef]
- Aranha, A.M.F.; Zhang, Z.; Neiva, K.G.; Costa, C.A.S.; Hebling, J.; Nör, J.E. Hypoxia enhances the angiogenic potential of human dental pulp cells. J. Endod. 2010, 36, 1633–1637. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Cao, Y.; Zhao, Q.; Li, J.; Zhou, C.; Liao, L.; Jia, M.; Zhao, Q.; Cai, H.; Han, Z.C.; et al. Proliferation and differentiation of bone marrow stromal cells under hypoxic conditions. Biochem. Biophys. Res. Commun. 2006, 347, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Dou, L.; Yan, Q.; Liang, P.; Zhou, P.; Zhang, Y.; Ji, P. iTRAQ-Based Proteomic Analysis Exploring the Influence of Hypoxia on the Proteome of Dental Pulp Stem Cells under 3D Culture. Proteomics 2018, 18, 1–23. [Google Scholar] [CrossRef]
- Du, H.C.; Jiang, L.; Geng, W.X.; Li, J.; Zhang, R.; Dang, J.G.; Shu, M.G.; Li, L.W. Growth Factor-Reinforced ECM Fabricated from Chemically Hypoxic MSC Sheet with Improved in Vivo Wound Repair Activity. BioMed Res. Int. 2017, 2017. [Google Scholar] [CrossRef]
- Wang, G.L.; Semenza, G.L. General involvement of hypoxia-inducible factor 1 in transcriptional response to hypoxia. Proc. Natl. Acad. Sci. USA 1993, 90, 4304–4308. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. Defining the role of hypoxia-inducible factor 1 in cancer biology and therapeutics. Oncogene 2010, 29, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Majmundar, A.J.; Wong, W.J.; Simon, M.C. Hypoxia-Inducible Factors and the Response to Hypoxic Stress. Mol. Cell 2010, 40, 294–309. [Google Scholar] [CrossRef] [PubMed]
- Chow, D.C.; Wenning, L.A.; Miller, W.M.; Papoutsakis, E.T. Modeling pO2 distributions in the bone marrow hematopoietic compartment. I. Krogh’s model. Biophys. J. 2001, 81, 675–684. [Google Scholar] [CrossRef]
- Gong, Q.M.; Quan, J.J.; Jiang, H.W.; Ling, J.Q. Regulation of the stromal cell-derived factor-1α-CXCR4 axis in human dental pulp cells. J. Endod. 2010, 36, 1499–1503. [Google Scholar] [CrossRef]
- Bidkhori, H.R.; Ahmadiankia, N.; Matin, M.M.; Heiranitabasi, A.; Farshchian, M.; Meshkin, H.N.; Shahriyari, M.; Dastpak, M.; Bahrami, A.R. Chemically primed bone-marrow derived mesenchymal stem cells show enhanced expression of chemokine receptors contributed to their migration capability. Iran. J. Basic Med. Sci. 2016, 19, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Heirani-Tabasi, A.; Naderi-Meshkin, H.; Matin, M.M.; Mirahmadi, M.; Shahriyari, M.; Ahmadiankia, N.; Sanjar Moussavi, N.; Bidkhori, H.R.; Raeesolmohaddeseen, M.; Bahrami, A.R. Augmented migration of mesenchymal stem cells correlates with the subsidiary CXCR4 variant. Cell Adhes. Migr. 2018, 12, 118–126. [Google Scholar] [CrossRef]
- Kuang, R.; Zhang, Z.; Jin, X.; Hu, J.; Shi, S.; Ni, L.; Ma, P.X. Nanofibrous spongy microspheres for the delivery of hypoxia-primed human dental pulp stem cells to regenerate vascularized dental pulp. Acta Biomater. 2016, 33, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Maslowska, A.L.; Bryniarska, N.; Kubiak, A.; Kaczmarzyk, T.; Stryjewska, M.S.; Noga, S.; Boruczkowski, D.; Madeja, Z.; Surma, E.Z. Multilineage Differentiation Potential of Human Dental Pulp Stem Cells-Impact of 3D and Hypoxic Environment on Osteogenesis In Vitro. Int. J. Mol. Sci. 2020, 21, 6172. [Google Scholar] [CrossRef] [PubMed]
- Dissanayaka, W.L.; Han, Y.; Zhang, L.; Zou, T.; Zhang, C. Bcl-2 Overexpression and Hypoxia Synergistically Enhance Angiogenic Properties of Dental Pulp Stem Cells. Int. J. Mol. Sci. 2020, 21, 6159. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| HIF-1α | 5′-CTT CTG AGC TCT GAT GAG GC-3′ | 5′-GAA AGC ACC ATC AGG AAG CC-3′ |
| OCT4 | 5′-TTT TGG TAC CCC AGG CTA TG-3′ | 5′-GCA GGC ACC TCA GTT TGA AT-3′ |
| SOX2 | 5′-GAG CTT TGC AGG AAG TTT GC-3′ | 5′-GCA AGA AGC CTC TCC TTG AA-3′ |
| ACTB | 5′-AGA GCT ACG AGC TGC CTG AC-3′ | 5′-AGC ACT GTG TTG GCG TAC AG-3′ |
| CoCl2 Treatment | Control | 5 µM | 10 µM | 20 µM | 40 µM |
|---|---|---|---|---|---|
| Absorbance at 560 nm | 0.46 ± 0.052 | 1.082 ± 0.098 | 0.57 ± 0.056 | 0.49 ± 0.086 | 0.13 ± 0.010 |
| p value (vs. Control) | <0.0001 | 0.0116 | 0.5352 | <0.0001 |
| CCK-8 Assay | Control | 10 µM CoCl2 | p Value (vs. Control) |
|---|---|---|---|
| Day 1 | 2.27 ± 0.27 | 2.10 ± 0.28 | 0.3709 |
| Day 2 | 2.21 ± 0.54 | 3.08 ± 0.21 | 0.0096 |
| Day 3 | 4.67 ± 0.53 | 5.4 ± 0.62 | 0.0853 |
| Day 4 | 4.81 ± 0.52 | 5.41 ± 0.21 | 0.0449 |
| Day 5 | 5.31 ± 0.18 | 5.58 ± 0.50 | 0.2820 |
| Day 6 | 5.59 ± 0.63 | 5.88 ± 0.92 | 0.5721 |
| Day 7 | 5.19 ± 0.90 | 5.97 ± 0.53 | 0.1391 |
| Day 8 | 5.08 ± 1.07 | 5.91 ± 0.90 | 0.2270 |
| Day 9 | 4.16 ± 0.36 | 5.59 ± 0.46 | 0.0007 |
| Day 10 | 2.89 ± 0.84 | 3.78 ± 0.44 | 0.0685 |
| Relative Gene Expression/ACTB | Control | 10 µM CoCl2 | p Value (vs. Control) |
|---|---|---|---|
| HIF-1α | 0.00065 ± 0.000019 | 0.0095 ± 0.0012 | <0.0001 |
| Growth Factors | Control | 10 µM CoCl2 |
|---|---|---|
| Ang-2 | 668.16 | 1112.30 |
| EGF | 15.29 | 90.72 |
| EPO | 1.87 | 2.76 |
| bFGF | 74.04 | 665.68 |
| G-CSF | 1.09 | 2.07 |
| GM-CSF | 1.36 | 3.80 |
| HGF | 324.40 | 127.92 |
| M-CSF | 2.01 | 1.58 |
| PDGF-AA | 67.88 | 56.08 |
| PDGF-BB | 6.04 | 3.09 |
| SCF | 174.10 | 399.87 |
| TGF-α | 3006.60 | 4472.06 |
| VEGF | 5697.17 | 11,058.64 |
| Cytokines | Control | 10 µM CoCl2 |
|---|---|---|
| IL-4 | 4.02 | 2.85 |
| IL-2 | 1.45 | 1.06 |
| CXCL10 | 12.97 | 16.16 |
| IL-1β | 6.07 | 2.76 |
| TNF-α | 8.59 | 10.30 |
| CCL2 | 11.62 | 27.84 |
| IL-17A | 0.76 | 0.05 |
| IL-6 | 2.85 | 0.79 |
| IL-10 | 22.79 | 12.97 |
| IFN-g | 16.83 | 8.51 |
| IL-12p17 | 0.16 | 0.03 |
| CXCL8 | 0.08 | 0.07 |
| TGF-β1 | 12.23 | 20.72 |
| Relative Gene Expression/ACTB | Control | 10 µM CoCl2 | p Value (vs. Control) |
|---|---|---|---|
| OCT4 | 0.0018 ± 0.00019 | 0.0023 ± 0.000054 | 0.0003 |
| SOX2 | 0.00036 ± 0.000062 | 0.00084 ± 0.00026 | 0.0039 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhandi, S.; Al Kahtani, A.; Mashyakhy, M.; Alsofi, L.; Maganur, P.C.; Vishwanathaiah, S.; Testarelli, L.; Del Giudice, A.; Mehta, D.; Vyas, N.; et al. Modulation of the Dental Pulp Stem Cell Secretory Profile by Hypoxia Induction Using Cobalt Chloride. J. Pers. Med. 2021, 11, 247. https://doi.org/10.3390/jpm11040247
Bhandi S, Al Kahtani A, Mashyakhy M, Alsofi L, Maganur PC, Vishwanathaiah S, Testarelli L, Del Giudice A, Mehta D, Vyas N, et al. Modulation of the Dental Pulp Stem Cell Secretory Profile by Hypoxia Induction Using Cobalt Chloride. Journal of Personalized Medicine. 2021; 11(4):247. https://doi.org/10.3390/jpm11040247
Chicago/Turabian StyleBhandi, Shilpa, Ahmed Al Kahtani, Mohammed Mashyakhy, Loai Alsofi, Prabhadevi C. Maganur, Satish Vishwanathaiah, Luca Testarelli, Andrea Del Giudice, Deepak Mehta, Nishant Vyas, and et al. 2021. "Modulation of the Dental Pulp Stem Cell Secretory Profile by Hypoxia Induction Using Cobalt Chloride" Journal of Personalized Medicine 11, no. 4: 247. https://doi.org/10.3390/jpm11040247
APA StyleBhandi, S., Al Kahtani, A., Mashyakhy, M., Alsofi, L., Maganur, P. C., Vishwanathaiah, S., Testarelli, L., Del Giudice, A., Mehta, D., Vyas, N., Patil, V. R., Raj, A. T., & Patil, S. (2021). Modulation of the Dental Pulp Stem Cell Secretory Profile by Hypoxia Induction Using Cobalt Chloride. Journal of Personalized Medicine, 11(4), 247. https://doi.org/10.3390/jpm11040247