
Prevention of Covid-19 Infection and Related Complications by Ozonized Oils

 ,
,  ,
,  , ,
, ,  , , ,
, , , 
Abstract
1. Introduction
2. Methods
2.1. In Vitro Studies
2.1.1. Cell Culture for the Viral Challenge Experiments
2.1.2. SARS-CoV-2 Challenge Experiments
2.1.3. RNA Extraction and qPCR Analyses
2.1.4. Prevention of SARS-CoV-2 Infection in Cell Culture
- (a)
- UV-C 254 nm radiation generated by LED, power 0.3 mW/cm2 (measured by Referenz Radiometer, Epigap Optoelekronik, GmbH, Bergkirchen, Germany) for 15 min, corresponding to a dose of 270 mJ/cm2.
- (b)
- Hydrogen peroxide: Analytical grade sterile hydrogen peroxide (Sigma, Milan City, Italy) was added at a final concentration of 1% vol/vol and incubated at room temperature for 15 min.
- (c)
- HOO (O3zone, GS Pharma, La Valletta, Malta): 0.5 mL was added to cultures and incubated at room temperature for 15 min. This ozonized oil was selected because (a) it had the highest level of ozonide available, i.e., >900 ozonides, and (b) it was the only ozonized oil among those tested which was able to penetrate inside pulmonary A549 cells (see below).
- (d)
- HOOws (O3zone spray, GS Pharma, La Valletta, Malta): It contained water, lecithin, polysorbate 20, and ozonized peanuts oil. In addition, 0.5 mL of HOOws was added to cultures and incubated at room temperature for 15 min.
- (e)
- HOO and HOOws in combination (1/1 vol/vol) incubated at room temperature for 15 min.
2.1.5. Evaluation of Anti-Inflammatory Capacity of Ozonized Oils
2.1.6. Evaluation of Ozonized Oil Penetration Inside Cultured Cells
2.2. In Vivo Studies
2.2.1. Evaluation of Increased Oxygen Availability in the Lungs after HOO Treatment
2.2.2. Evaluation of Safety and Efficacy of Ozonized Oils in Human Patients
Study Design and Participants
Statistical Analyses
3. Results
3.1. In Vitro Studies
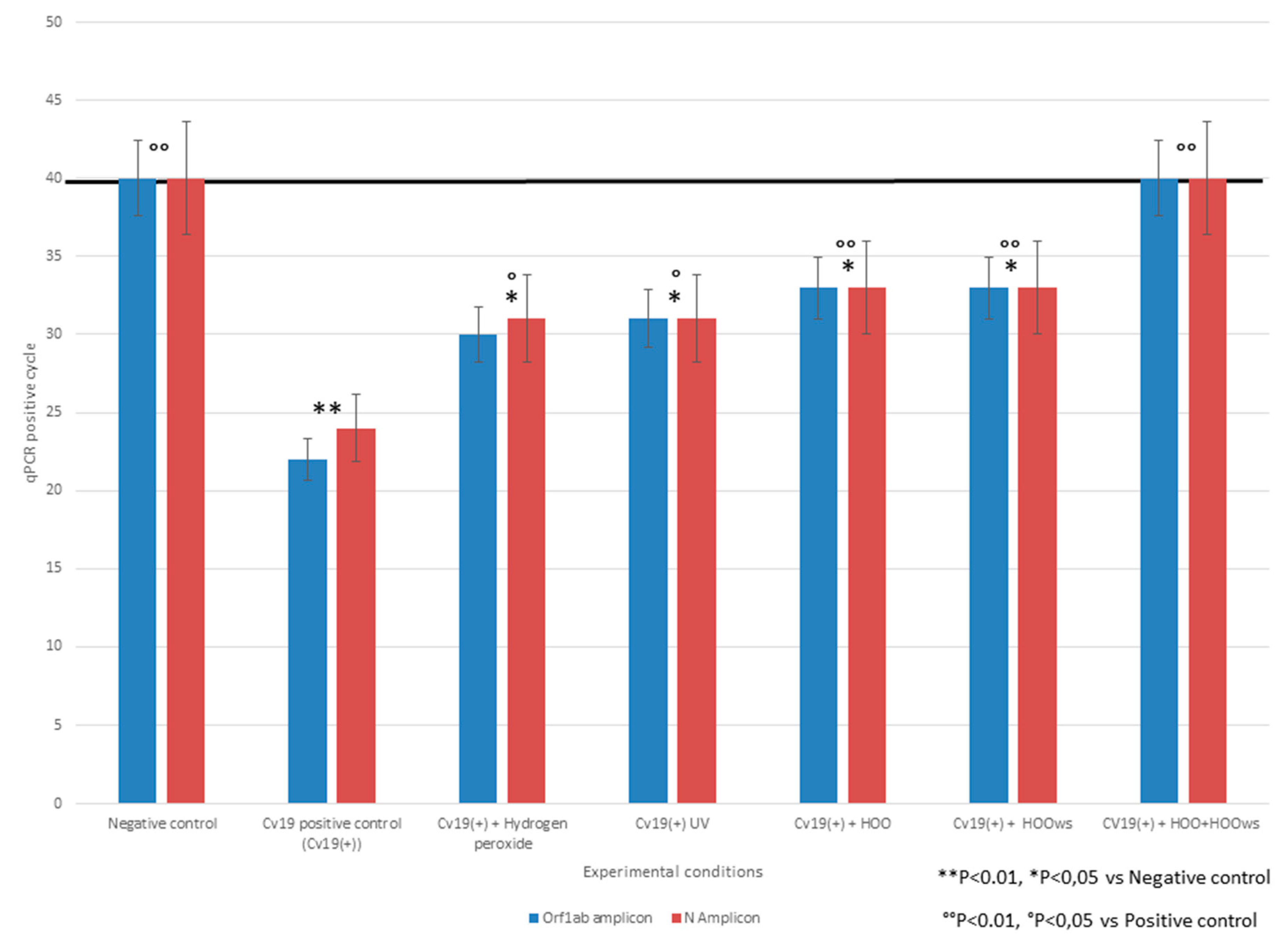
3.1.1. SARS-Cov-2 Challenge Test and qPCR Analyses
3.1.2. Efficacy in Preventing SARS-Cov-2 Infection
3.1.3. Evaluation of Anti-Inflammatory Capacity of Ozonized Oils
3.1.4. Evaluation of Ozonized Oil’s Capacity to Penetrate inside Cultured Cells
3.2. In Vivo Studies
Evaluation of Increased Oxygen Availability in the Lungs after Ozonized Oil Intake
3.3. In Vivo Safety in Human Patients
Anti-Viral Efficacy in Human Patients
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Jiang, H.J.; Chen, N.; Shen, Z.Q.; Jing, Y.; Gang, Q.Z.; Jing, M.; Wei, Y.Z.; Yang, S.D.; Wang, H.R.; Wei, W.X.; et al. Inactivation of Poliovirus by Ozone and the Impact of Ozone on the Viral Genome. Biomed. Environ. Sci. 2019, 32, 324–333. [Google Scholar] [PubMed]
- Torrey, J.; von Gunten, U.; Kohn, T. Differences in Viral Disinfection Mechanisms as Revealed by Quantitative Transfection of Echovirus 11 Genomes. Appl Environ. Microbiol. 2019, 85, e00961-19. [Google Scholar] [CrossRef] [PubMed]
- Barh, D.; Tiwari, S.E.; Weener, M.E.; Azevedo, V.; Góes-Neto, A.; Gromiha, M.M.; Ghosh, P. Multi-omics-based identification of SARS-CoV-2 infection biology and candidate drugs against COVID-19. Comput Biol Med. 2020, 126, 104051. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Sánchez, G.; Schwartz, A.; Donna, V.D. Potential Cytoprotective Activity of Ozone Therapy in SARS-CoV-2/COVID-19. Antioxidants 2020, 9, 389. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Dong, M.; Hu, K. A preliminary evaluation on the efficacy of ozone therapy in the treatment of COVID-19. J. Med. Virol. 2020. [Google Scholar] [CrossRef]
- Valdenassi, L.; Franzini, M.; Ricevuti, G.; Rinaldi, L.; Galoforo, A.C.; Tirelli, U. Potential mechanisms by which the oxygen-ozone (O2-O3) therapy could contributeto the treatment against the coronavirus COVID-19. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 4059–4061. [Google Scholar]
- Fernández-Cuadros, M.E.; Albaladejo-Florín, M.J.; Peña-Lora, D.; Álava-Rabasa, S.; Pérez-Moro, O.S. Ozone (O3) and SARS-CoV-2: Physiological Bases and Their Therapeutic Possibilities According to COVID-19 Evolutionary Stage. SN Compr. Clin. Med. 2020, 1–9. [Google Scholar] [CrossRef]
- Franzini, M.; Valdenassi, L.; Ricevuti, G.; Chirumbolo, S.; Depfenhart, M.; Bertossi, D.; Tirelli, U. Oxygen-ozone (O 2-O 3) immunoceutical therapy for patients with COVID-19. Preliminary evidence reported. Int. Immunopharmacol. 2020, 88, 106879. [Google Scholar] [CrossRef]
- Ugazio, E.; Tullio, V.; Binello, A.; Tagliapietra, S.; Dosio, F. Ozonated Oils as Antimicrobial Systems in Topical Applications. Their Characterization, Current Applications, and Advances in Improved Delivery Techniques. Molecules 2020, 25, 334. [Google Scholar] [CrossRef]
- Kampf, G.; Todt, D.; Pfaender, S.; Steinmann, E. Persistence of coronaviruses on inanimate surfaces and their inactivation with biocidal agents. J. Hosp. Infect. 2020, 104, 246–251. [Google Scholar] [CrossRef]
- Kam, Y.W.; Okumura, Y.; Kido, H.; Ng, L.F.; Bruzzone, R.; Altmeyer, R. Cleavage of the SARS coronavirus spike glycoprotein by airway proteases enhances virus entry into human bronchial epithelial cells in vitro. PLoS ONE 2009, 4, e7870. [Google Scholar] [CrossRef]
- Calzia, D.; Ottaggio, L.; Cora, A.; Chiappori, G.; Cuccarolo, P.; Cappelli, E.; Izzotti, A.; Tavella, S.; Degan, P. Characterization of C2C12 cells in simulated microgravity: Possible use for myoblast regeneration. J. Cell Physiol. 2020, 235, 3508–3518. [Google Scholar] [CrossRef]
- Keyaerts, E.; Vijgen, L.; Maes, P.; Neyts, J.; Van Ranst, M. Growth kinetics of SARS-coronavirus in Vero E6 cells. Biochem. Biophys. Res. Commun. 2005, 15, 1147–1151. [Google Scholar] [CrossRef]
- Williams, J.G.; Maier, R.V. Ketoconazole inhibits alveolar macrophage production of inflammatory mediators involved in acute lung injury (adult respiratory distress syndrome). Surgery 1992, 112, 270–277. [Google Scholar]
- Tartari, A.P.S.; Moreira, F.F.; Pereira, M.C.D.; Carraro, E.; Cidral-Filho, F.J.; Salgado, A.I.; Kerppers, I.I. Anti-inflammatory Effect of Ozone Therapy in an Experimental Model of Rheumatoid Arthritis. Inflammation 2020, 43, 985–993. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Chen, J.; Ren, G.; Zhang, Y.; Tan, X.; Yang, L. Punicalagin Prevents Inflammation in LPS-Induced RAW264.7 Macrophages by Inhibiting FoxO3a/Autophagy Signaling Pathway. Nutrients 2019, 11, 2794. [Google Scholar] [CrossRef] [PubMed]
- Thevenet, D.; Tardieu-Berger, M.; Zouhal, H.; Jacob, C.; Abderrahman, B.A.; Prioux, J. Influence of exercise intensity on time spent at high percentage of maximal oxygen uptake during an intermittent session in young endurance-trained athletes. Eur. J. Appl. Physiol. 2007, 102, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Sungnak, W.; Huang, N.; Bécavin, C.; Berg, M.; Queen, R.; Litvinukova, M.; Talavera-López, C.; Maatz, H.; Reichart, D.; Sampaziotis, F.; et al. HCA Lung Biological Network. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat. Med. 2020, 26, 681–687. [Google Scholar] [CrossRef]
- Kurosaki, Y.; Abe, H.; Morioka, H.; Hirayama, J.; Ikebuchi, K.; Kamo, N.; Nikaido, O.; Azuma, H.; Ikeda, H. Pyrimidine dimer formation and oxidative damage in M13 bacteriophage inactivation by ultraviolet C irradiation. Photochem. Photobiol. 2003, 78, 349–354. [Google Scholar] [CrossRef]
- Hessling, M.; Haag, R.; Sieber, N.; Vatter, P. The impact of far-UVC radiation (200-230 nm) on pathogens, cells, skin, and eyes—A collection and analysis of a hundred years of data. GMS Hyg. Infect. Control. 2021, 16, 16. [Google Scholar]
- Heilingloh, C.S.; Aufderhorst, U.W.; Schipper, L.; Dittmer, U.; Witzke, O.; Yang, D.; Zheng, X.; Sutter, K.; Trilling, M.; Alt, M.; et al. Susceptibility of SARS-CoV-2 to UV irradiation. Am. J. Infect. Control. 2020, 48, 1273–1275. [Google Scholar] [CrossRef]
- Ke, Z.; Oton, J.; Qu, K.; Cortese, M.; Zila, V.; McKeane, L.; Nakane, T.; Zivanov, J.; Neufeldt, C.J.; Cerikan, B.; et al. Structures and distributions of SARS-CoV-2 spike proteins on intact virions. Nature 2020. [Google Scholar] [CrossRef] [PubMed]
- Turunova, B.; Sikora, M.; Schurman, C.; Hagen, W.J.; Welsch, S.; Blanc, F.E.C.; von Bülow, S.; Gecht, M.; Bagol, K.; Hörner, C.; et al. In situ structural analysis of SARS-CoV-2 spike reveas flexibility mediated by three hinges. Science 2020, 370, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Iadecola, C.; Anrather, J.; Kamel, H. Effects of COVID-19 on the Nervous System. Cell 2020, 183, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Jin, H.; Wang, M.; Hu, Y.; Chen, S.; He, Q.; Chang, J.; Hong, C.; Zhou, Y.; Wang, D.; et al. Neurologic Manifestations of Hospitalized Patients With Coronavirus Disease 2019 in Wuhan, China. JAMA Neurol. 2020, 77, 683–690. [Google Scholar] [CrossRef]
- Buzhdygan, T.P.; DeOre, B.J.; Baldwin-Leclair, A.; Bullock, T.A.; McGary, H.M.; Khan, J.A.; Razmpour, R.; Hale, J.F.; Galie, P.A.; Potula, R.; et al. The SARS-CoV-2 spike protein alters barrier function in 2D static and 3D microfluidic in-vitro models of the human blood-brain barrier. Neurobiol. Dis. 2020, 146, 105131. [Google Scholar] [CrossRef]
- Caldas, L.A.; Carneiro, F.A.; Hiha, L.M.; Monteiro, F.L.; da Silva, G.P.; da Costa, L.J.; Durigon, E.L.; Tanuri, A.; de Souza, W. Ultrastructural analysis of SARS-CoV-2 interactions with the host cell via high resolution scanning electron microscopy. Sci. Rep. 2020, 10, 16099. [Google Scholar] [CrossRef]
- Oh, W.K. COVID-19 infection in cancer patients: Early observations and unanswered questions. Ann. Oncol. 2020, 31, 838–839. [Google Scholar] [CrossRef]
- Cento, V.; Colagrossi, L.; Nava, A.; Lamberti, A.; Senatore, S.; Travi, G.; Rossotti, R.; Vecchi, M.; Casati, O.; Matarazzo, E.; et al. Persistent positivity and fluctuations of SARS-CoV2-RNA in clinically recovered COVID-19 patients. J. Infect. 2020, 81, 90–92. [Google Scholar] [CrossRef]
- Machhi, J.; Herskovitz, J.; Senan, A.M.; Dutta, D.; Nath, B.; Oleynikov, M.D.; Blomberg, W.R.; Meigs, D.D.; Hasan, M.; Patel, M.; et al. The Natural History, Pathobiology, and Clinical Manifestations of SARS-CoV-2 Infections. J. Neuroimmune Pharmacol. 2020, 15, 359–386. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| T0 PRE-TREATMENT TEST | VO2 Max (mL/kg/min) | VO2 (L/min) | VO2 Threshold (mL/kg/min) | VO2 Threshold (L/min) | % VO2 Max in Threshold |
|---|---|---|---|---|---|
| Subject 1 | 39.4 | 3.19 | 32.5 | 2.62 | 82 |
| Subject 2 | 36.6 | 2.42 | 30.6 | 2.07 | 86 |
| T1 POST-TREATMENT TEST | |||||
| Subject 1 | 40.9 | 3.27 | 35.6 | 2.94 | 90 |
| Subject 2 | 38.9 | 2.63 | 33.4 | 2.08 | 87 |
| T1-T0 delta after HOO TREATMENT | |||||
| Subject 1 | +1.5 | +0.08 | +2.9 | +0.32 | +8 |
| Subject 2 | +2.3 | +0.21 | +2.8 | +0.01 | +1 |
| Gender | Age | Previous Diseases | COVID-19 Infection | Clinical Outcome |
|---|---|---|---|---|
| Therapeutic purpose | ||||
| Female | 22 | None | Yes | Recovery |
| Male | 55 | COPD, Vascular ischemia | Yes | Recovery |
| Female | 54 | None | Yes | Recovery |
| Female | 52 | None | Yes | Recovery |
| Chemo-prophylactic purpose. Cancer patients | ||||
| Male | 47 | Brain cancer (glioblastoma) | No | No Covid-19 infection |
| Male | 40 | Brain cancer (glioblastoma) | No | No Covid-19 infection |
| Male | 19 | Brain cancer (glioblastoma) | No | No Covid-19 infection |
| Female | 11 | Brain cancer (glioblastoma) | No | No Covid-19 infection |
| Male | 48 | Brain cancer (glioblastoma) | No | No Covid-19 infection |
| Male | 33 | Brain cancer (glioblastoma) | No | No Covid-19 infection |
| Male | 40 | Brain cancer (glioblastoma) | No | No Covid-19 infection |
| Female | 37 | Brain cancer (glioblastoma) | No | No Covid-19 infection |
| Female | 40 | Brain cancer (glioblastoma) | No | No Covid-19 infection |
| Female | 50 | Breast cancer | No | No Covid-19 infection |
| Female | 56 | Breast cancer | No | No Covid-19 infection |
| Female | 60 | Breast cancer | No | No Covid-19 infection |
| Female | 69 | Breast cancer | No | No Covid-19 infection |
| Female | 65 | Breast cancer | No | No Covid-19 infection |
| Female | 55 | Breast cancer | No | No Covid-19 infection |
| Female | 52 | Breast cancer | No | No Covid-19 infection |
| Female | 70 | Breast cancer | No | No Covid-19 infection |
| Male | 57 | Colon cancer | No | No Covid-19 infection |
| Female | 61 | Colon cancer | No | No Covid-19 infection |
| Male | 53 | Colon cancer | No | No Covid-19 infection |
| Female | 57 | Colon cancer | No | No Covid-19 infection |
| Male | 78 | Kidney cancer | No | No Covid-19 infection |
| Male | 73 | Bladder cancer | No | No Covid-19 infection |
| Female | 82 | Non-Hodgkin Lymphoma | No | No Covid-19 infection |
| Male | 54 | Non-Hodgkin Lymphoma | No | No Covid-19 infection |
| Male | 81 | Lung cancer (NSCLC) | No | No Covid-19 infection |
| Male | 58 | Lung cancer (NSCLC) | No | No Covid-19 infection |
| Female | 55 | Lung cancer (SCLC) | No | No Covid-19 infection |
| Male | 27 | Lung cancer (SCLC) | No | No Covid-19 infection |
| Male | 79 | Lung cancer (NSCLC) | No | No Covid-19 infection |
| Male | 76 | Lung cancer (NSCLC) | No | No Covid-19 infection |
| Female | 74 | Ovarian cancer | No | No Covid-19 infection |
| Female | 75 | Ovarian cancer | No | No Covid-19 infection |
| Female | 66 | Ovarian cancer | No | No Covid-19 infection |
| Female | 28 | Womb cancer | No | No Covid-19 infection |
| Female | 62 | Pancreas cancer | No | No Covid-19 infection |
| Female | 78 | Pancreas cancer | No | No Covid-19 infection |
| Male | 72 | Pancreas cancer | No | No Covid-19 infection |
| Female | 58 | Pancreas cancer | No | No Covid-19 infection |
| Male | 63 | Pancreas cancer | No | No Covid-19 infection |
| Female | 79 | Pancreas cancer | No | No Covid-19 infection |
| Male | 60 | Pancreas cancer | No | No Covid-19 infection |
| Male | 67 | Pancreas cancer | No | No Covid-19 infection |
| Male | 71 | Prostate cancer | No | No Covid-19 infection |
| Male | 80 | Prostate cancer | No | No Covid-19 infection |
| Male | 83 | Prostate cancer | No | No Covid-19 infection |
| Male | 58 | Prostate cancer | No | No Covid-19 infection |
| Male | 61 | Prostate cancer | No | No Covid-19 infection |
| Female | 92 | Skin cancer (basal cell carcinoma) | No | No Covid-19 infection |
| Male | 70 | Oral cancer (squamous cell carcinoma) | No | No Covid-19 infection |
| Male | 89 | Skin cancer (basal cell carcinoma) | No | No Covid-19 infection |
| Male | 77 | Skin cancer (angiosarcoma) | No | No Covid-19 infection |
| Chemo-prophylactic purpose. Healthy subjects | ||||
| Female | 32 | None | No | No Covid-19 infection |
| Female | 12 | None | No | No Covid-19 infection |
| Female | 18 | None | No | No Covid-19 infection |
| Male | 72 | None | No | No Covid-19 infection |
| Female | 45 | None | No | No Covid-19 infection |
| Female | 32 | None | No | No Covid-19 infection |
| Female | 38 | None | No | No Covid-19 infection |
| Male | 45 | None | No | No Covid-19 infection |
| Male | 59 | None | No | No Covid-19 infection |
| Male | 64 | None | No | No Covid-19 infection |
| Female | 49 | None | No | No Covid-19 infection |
| Female | 93 | None | No | No Covid-19 infection |
| Male | 61 | None | No | No Covid-19 infection |
| Male | 52 | None | No | No Covid-19 infection |
| Male | 34 | None | No | No Covid-19 infection |
| Female | 36 | None | No | No Covid-19 infection |
| Female | 48 | None | No | No Covid-19 infection |
| Male | 62 | None | No | No Covid-19 infection |
| Male | 46 | None | No | No Covid-19 infection |
| Female | 51 | None | No | No Covid-19 infection |
| Male | 80 | None | No | No Covid-19 infection |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Izzotti, A.; Fracchia, E.; Au, W.; Colombo, M.; Pfeffer, U.; Emionite, L.; Pavan, S.; Miotto, D.; Lova, P.; Grasselli, E.; et al. Prevention of Covid-19 Infection and Related Complications by Ozonized Oils. J. Pers. Med. 2021, 11, 226. https://doi.org/10.3390/jpm11030226
Izzotti A, Fracchia E, Au W, Colombo M, Pfeffer U, Emionite L, Pavan S, Miotto D, Lova P, Grasselli E, et al. Prevention of Covid-19 Infection and Related Complications by Ozonized Oils. Journal of Personalized Medicine. 2021; 11(3):226. https://doi.org/10.3390/jpm11030226
Chicago/Turabian StyleIzzotti, Alberto, Enzo Fracchia, William Au, Monica Colombo, Ulrich Pfeffer, Laura Emionite, Simone Pavan, Daniele Miotto, Paola Lova, Elena Grasselli, and et al. 2021. "Prevention of Covid-19 Infection and Related Complications by Ozonized Oils" Journal of Personalized Medicine 11, no. 3: 226. https://doi.org/10.3390/jpm11030226
APA StyleIzzotti, A., Fracchia, E., Au, W., Colombo, M., Pfeffer, U., Emionite, L., Pavan, S., Miotto, D., Lova, P., Grasselli, E., Faelli, E., Piero, R., Tiso, M., & Pulliero, A. (2021). Prevention of Covid-19 Infection and Related Complications by Ozonized Oils. Journal of Personalized Medicine, 11(3), 226. https://doi.org/10.3390/jpm11030226