
Knockout of Hepatocyte Growth Factor by CRISPR/Cas9 System Induces Apoptosis in Hepatocellular Carcinoma Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemical Reagents and Antibodies
2.2. Cell Culture
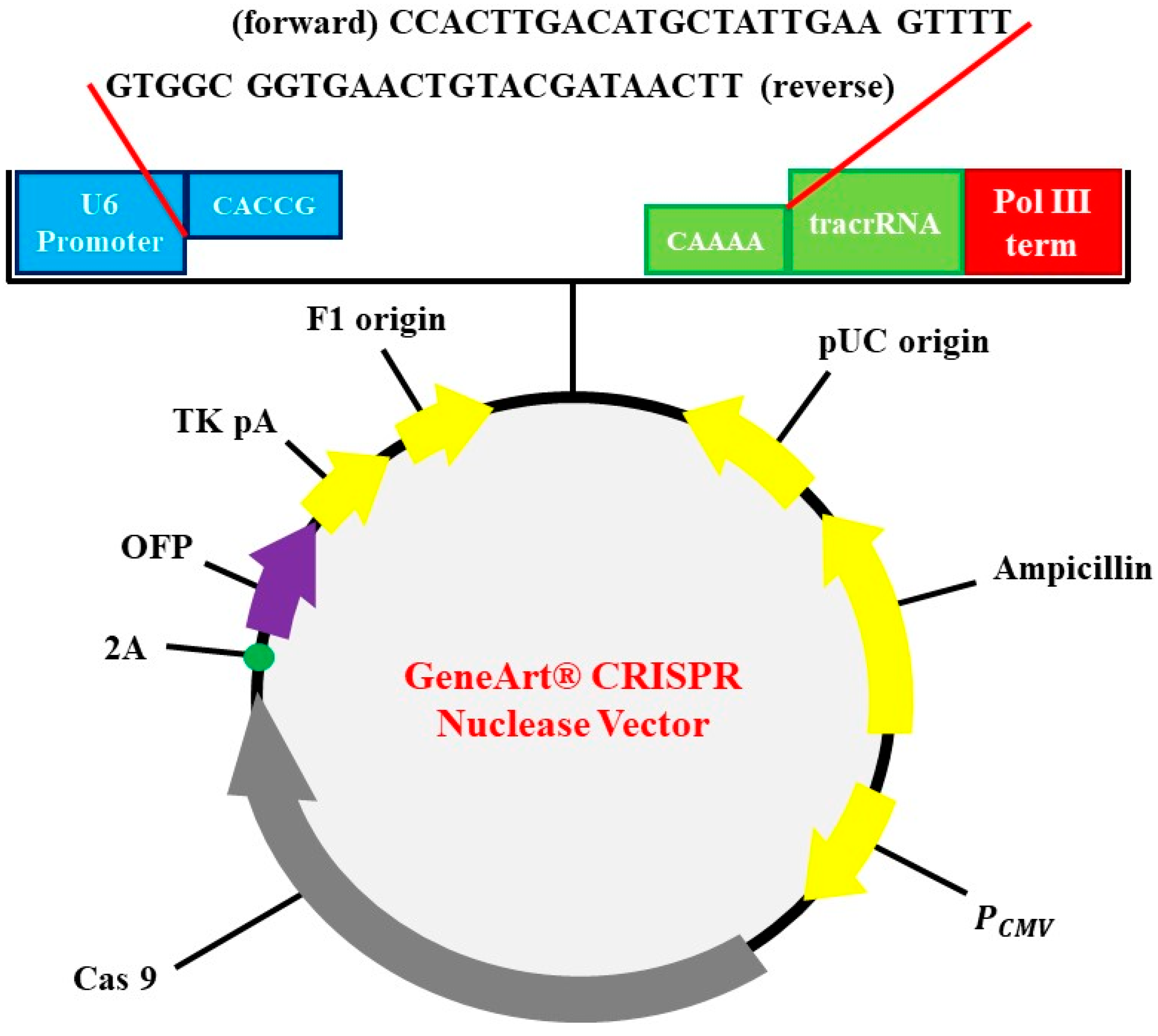
2.3. Plasmid Construction
2.4. Transfection
2.5. Real-Time Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.6. Conditioned Media Preparation
2.7. Cell Counting Assay
2.8. Colony Formation Assay
2.9. Wound Healing Assay
2.10. Invasion Assay
2.11. Cell Cycle Analysis
2.12. Annexin V/PI Staining Assay
2.13. Western Blot Assay
2.14. Statistical Analysis
3. Results
3.1. Transfection of Crispr-HGF Downregulated the Expression Levels of HGF mRNA in Huh7 and Hep3B Cells
3.2. HGF Stimulated c-Met Signaling Pathways and Transfection of Crispr-HGF Regulated the Expression of HGF and MAPKs in Huh7 and Hep3B Cells
3.3. Transfection of Crispr-HGF Inhibited the Cell Proliferation in Huh7 and Hep3B Cells
3.4. Transfection of Crispr-HGF Suppressed Migration and Invasion Ability in Huh7 and Hep3B Cells
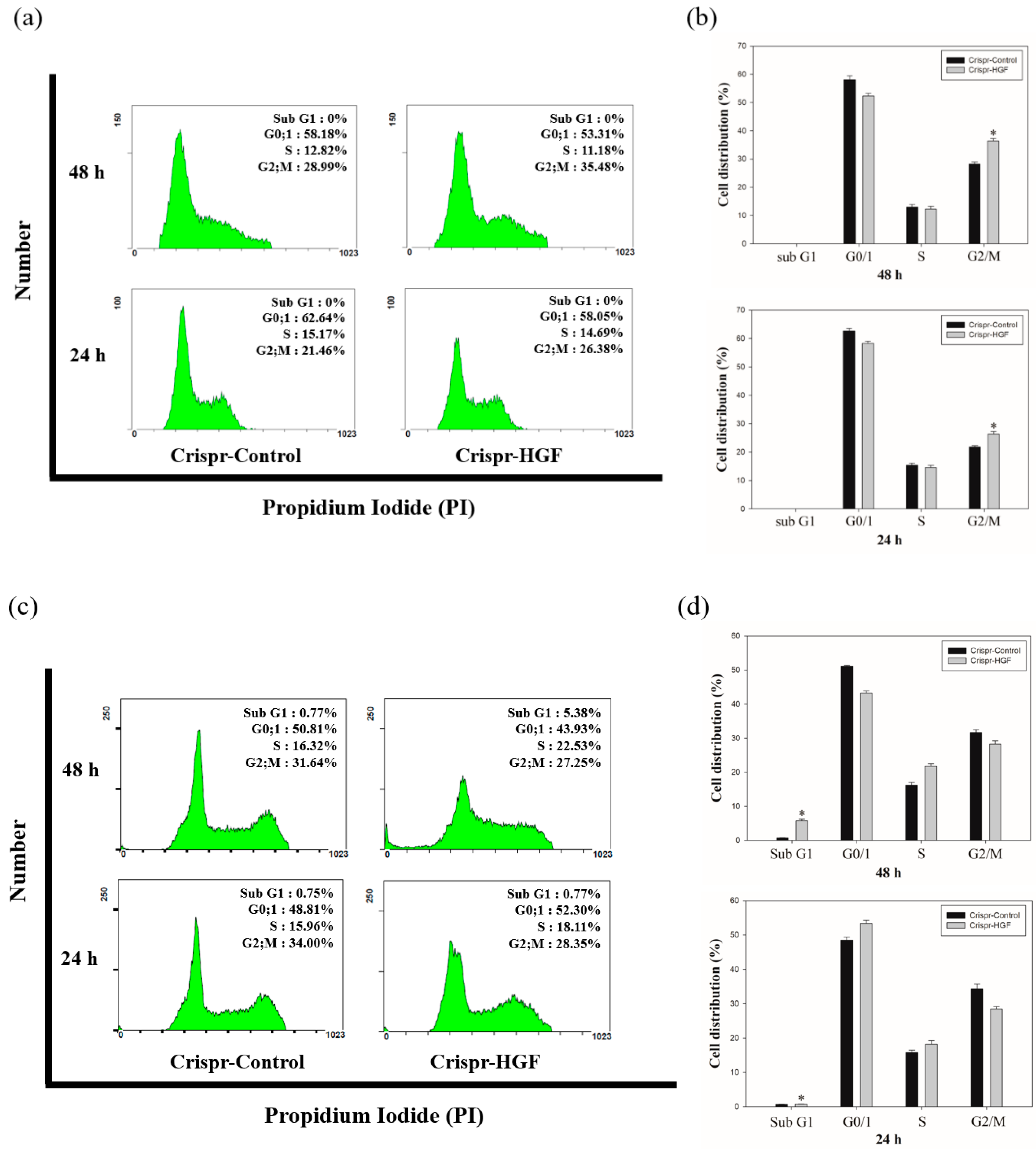
3.5. Transfection of Crispr-HGF Induced Cell Cycle Arrest in Huh7 and Hep3B Cells
3.6. Transfection of Crispr-HGF Induced Apoptosis in Huh7 and Hep3B Cells
3.7. H2O2 Induced Synergistic Apoptotic Effects with Transfection of Crispr-HGF against Huh7 and Hep3B Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| HCC | Hepatocellular carcinoma |
| CRISPR/Cas9 | Clustered regularly interspaced short palindromic repeats (CRISPR)-associated (Cas) protein 9 (CRISPR/Cas9) system |
| HGF | Hepatocyte growth factor |
| RIPA | Radioimmunoprecipitation assay |
| PARP | Poly (ADP-ribose) polymerase |
| Akt | Protein kinase B (PKB) |
| DMSO | Dimethyl sulfoxide |
| FBS | Fetal bovine serum |
| PBS | Phosphate-buffered saline |
| PI | Propidium iodide |
References
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular carcinoma. Nat. Rev. Dis. Primers 2021, 7, 6. [Google Scholar] [CrossRef]
- Puoti, C. New insights on hepatocellular carcinoma: Epidemiology and clinical aspects. Hepatoma Res. 2018, 4, 57. [Google Scholar] [CrossRef]
- Lodato, F.; Mazzella, G.; Festi, D.; Azzaroli, F.; Colecchia, A.; Roda, E. Hepatocellular carcinoma prevention: A worldwide emergence between the opulence of developed countries and the economic constraints of developing nations. World J. Gastroenterol. 2006, 12, 7239–7249. [Google Scholar] [CrossRef] [PubMed]
- Moini, M.; Schilsky, M.L.; Tichy, E.M. Review on immunosuppression in liver transplantation. World J. Hepatol. 2015, 7, 1355–1368. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.A.; Jeon, Y.K.; Nam, M.J. Auricularia auricula increases an apoptosis in human hepatocellular carcinoma cells via a regulation of the peroxiredoxin1. J. Food Biochem. 2020, 44, e13373. [Google Scholar] [CrossRef]
- Akram, F.; Ikram Ul, H.; Ahmed, Z.; Khan, H.; Ali, M.S. CRISPR-Cas9, A Promising Therapeutic Tool for Cancer Therapy: A Review. Protein Pept. Lett. 2020, 27, 931–944. [Google Scholar] [CrossRef]
- Kuang, W.; Deng, Q.; Deng, C.; Li, W.; Shu, S.; Zhou, M. Hepatocyte growth factor induces breast cancer cell invasion via the PI3K/Akt and p38 MAPK signaling pathways to up-regulate the expression of COX2. Am. J. Transl. Res. 2017, 9, 3816–3826. [Google Scholar]
- Lesko, E.; Majka, M. The biological role of HGF-MET axis in tumor growth and development of metastasis. Front. Biosci. 2008, 13, 1271–1280. [Google Scholar] [CrossRef]
- Xiang, C.; Chen, J.; Fu, P. HGF/Met Signaling in Cancer Invasion: The Impact on Cytoskeleton Remodeling. Cancers 2017, 9, 44. [Google Scholar] [CrossRef]
- Matsumoto, K.; Umitsu, M.; De Silva, D.M.; Roy, A.; Bottaro, D.P. Hepatocyte growth factor/MET in cancer progression and biomarker discovery. Cancer Sci. 2017, 108, 296–307. [Google Scholar] [CrossRef]
- Boschert, V.; Klenk, N.; Abt, A.; Janaki Raman, S.; Fischer, M.; Brands, R.C.; Seher, A.; Linz, C.; Müller-Richter, U.D.A.; Bischler, T.; et al. The Influence of Met Receptor Level on HGF-Induced Glycolytic Reprogramming in Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2020, 21, 471. [Google Scholar] [CrossRef] [PubMed]
- Breunig, C.; Erdem, N.; Bott, A.; Greiwe, J.F.; Reinz, E.; Bernhardt, S.; Giacomelli, C.; Wachter, A.; Kanthelhardt, E.J.; Beißbarth, T.; et al. TGFβ1 regulates HGF-induced cell migration and hepatocyte growth factor receptor MET expression via C-ets-1 and miR-128-3p in basal-like breast cancer. Mol. Oncol. 2018, 12, 1447–1463. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Lee, C.M.; Lee, J.H.; Park, S.H.; Nam, M.J. Protective effects of hepatocyte growth factor gene overexpression against hydrogen peroxide-induced apoptosis in mesenchymal stem cells. Environ. Toxicol. 2019, 34, 1236–1245. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, A.; da Silva, G.J. CRISPR-Cas: Converting A Bacterial Defence Mechanism into A State-of-the-Art Genetic Manipulation Tool. Antibiotics 2019, 8, 18. [Google Scholar] [CrossRef] [PubMed]
- Zhan, T.; Rindtorff, N.; Betge, J.; Ebert, M.P.; Boutros, M. CRISPR/Cas9 for cancer research and therapy. Semin. Cancer Biol. 2019, 55, 106–119. [Google Scholar] [CrossRef]
- Chesnut, J.; Namritha, R.; Kumar, S.; Garza, J. Efficient and Multiplex Genome Editing Using GeneArt® CRISPR Nuclease mRNA; Thermo Fisher Scientific: Waltham, MA, USA, 2015. [Google Scholar]
- Lee, C.M.; Lee, J.; Jang, S.-N.; Shon, J.C.; Wu, Z.; Park, K.; Liu, K.-H.; Park, S.-H. 6,8-Diprenylorobol Induces Apoptosis in Human Hepatocellular Carcinoma Cells via Activation of FOXO3 and Inhibition of CYP2J2. Oxidative Med. Cell. Longev. 2020, 2020, 8887251. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.M.; Choi, Y.J.; Park, S.-H.; Nam, M.J. Indole-3-carbinol induces apoptosis in human hepatocellular carcinoma Huh-7 cells. Food Chem. Toxicol. 2018, 118, 119–130. [Google Scholar] [CrossRef]
- Park, S.H.; Phuc, N.M.; Lee, J.; Wu, Z.; Kim, J.; Kim, H.; Kim, N.D.; Lee, T.; Song, K.S.; Liu, K.H. Identification of acetylshikonin as the novel CYP2J2 inhibitor with anti-cancer activity in HepG2 cells. Phytomedicine 2017, 24, 134–140. [Google Scholar] [CrossRef]
- Li, B.; Leung, J.C.K.; Chan, L.Y.Y.; Yiu, W.H.; Li, Y.; Lok, S.W.Y.; Liu, W.H.; Chan, K.W.; Tse, H.F.; Lai, K.N.; et al. Amelioration of Endoplasmic Reticulum Stress by Mesenchymal Stem Cells via Hepatocyte Growth Factor/c-Met Signaling in Obesity-Associated Kidney Injury. STEM CELLS Transl. Med. 2019, 8, 898–910. [Google Scholar] [CrossRef]
- Zeng, W.; Ju, R.; Mao, M. Therapeutic potential of hepatocyte growth factor against cerebral ischemia (Review). Exp. Med. 2015, 9, 283–288. [Google Scholar] [CrossRef]
- Nakamura, T.; Mizuno, S. The discovery of hepatocyte growth factor (HGF) and its significance for cell biology, life sciences and clinical medicine. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2010, 86, 588–610. [Google Scholar] [CrossRef] [PubMed]
- Longati, P.; Albero, D.; Comoglio, P.M. Hepatocyte growth factor is a pleiotropic factor protecting epithelial cells from apoptosis. Cell Death Differ. 1996, 3, 23–28. [Google Scholar]
- Sato, H.; Aoki, S.; Kato, T.; Matsumoto, K. HGF (Hepatocyte Growth Factor). In Encyclopedia of Signaling Molecules; Choi, S., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 2352–2364. [Google Scholar]
- Fukushima, T.; Uchiyama, S.; Tanaka, H.; Kataoka, H. Hepatocyte Growth Factor Activator: A Proteinase Linking Tissue Injury with Repair. Int. J. Mol. Sci. 2018, 19, 3435. [Google Scholar] [CrossRef] [PubMed]
- Galimi, F.; Cottone, E.; Vigna, E.; Arena, N.; Boccaccio, C.; Giordano, S.; Naldini, L.; Comoglio, P.M. Hepatocyte Growth Factor Is a Regulator of Monocyte-Macrophage Function. J. Immunol. 2001, 166, 1241. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xia, M.; Jin, K.; Wang, S.; Wei, H.; Fan, C.; Wu, Y.; Li, X.; Li, X.; Li, G.; et al. Function of the c-Met receptor tyrosine kinase in carcinogenesis and associated therapeutic opportunities. Mol. Cancer 2018, 17, 45. [Google Scholar] [CrossRef]
- Organ, S.L.; Tsao, M.-S. An overview of the c-MET signaling pathway. Adv. Med. Oncol. 2011, 3, S7–S19. [Google Scholar] [CrossRef]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis (Review). Exp. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, H.T. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef]
- He, X.; Wang, C.; Wang, H.; Li, L.; Wang, C. The Function of MAPK Cascades in Response to Various Stresses in Horticultural Plants. Front. Plant. Sci. 2020, 11, 952. [Google Scholar] [CrossRef]
- Huynh, H.; Nguyen, T.T.T.; Chow, K.-H.K.-P.; Tan, P.H.; Soo, K.C.; Tran, E. Over-expression of the mitogen-activated protein kinase (MAPK) kinase (MEK)-MAPK in hepatocellular carcinoma: Its role in tumor progression and apoptosis. BMC Gastroenterol. 2003, 3, 19. [Google Scholar] [CrossRef]
- Jung, K.H.; Park, B.H.; Hong, S.S. Progress in cancer therapy targeting c-Met signaling pathway. Arch. Pharm. Res. 2012, 35, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Herrero-Fresneda, I.; Torras, J.; Franquesa, M.; Vidal, A.; Cruzado, J.M.; Lloberas, N.; Fillat, C.; Grinyó, J.M. HGF gene therapy attenuates renal allograft scarring by preventing the profibrotic inflammatory-induced mechanisms. Kidney Int. 2006, 70, 265–274. [Google Scholar] [CrossRef]
- Nagai, T.; Arao, T.; Furuta, K.; Sakai, K.; Kudo, K.; Kaneda, H.; Tamura, D.; Aomatsu, K.; Kimura, H.; Fujita, Y.; et al. Sorafenib inhibits the hepatocyte growth factor-mediated epithelial mesenchymal transition in hepatocellular carcinoma. Mol. Cancer 2011, 10, 169–177. [Google Scholar] [CrossRef]
- Dai, L.; Trillo-Tinoco, J.; Cao, Y.; Bonstaff, K.; Doyle, L.; Del Valle, L.; Whitby, D.; Parsons, C.; Reiss, K.; Zabaleta, J.; et al. Targeting HGF/c-MET induces cell cycle arrest, DNA damage, and apoptosis for primary effusion lymphoma. Blood 2015, 126, 2821–2831. [Google Scholar] [CrossRef]
- Derksen, P.W.B.; de Gorter, D.J.J.; Meijer, H.P.; Bende, R.J.; van Dijk, M.; Lokhorst, H.M.; Bloem, A.C.; Spaargaren, M.; Pals, S.T. The hepatocyte growth factor/Met pathway controls proliferation and apoptosis in multiple myeloma. Leukemia 2003, 17, 764–774. [Google Scholar] [CrossRef]
- Konturek, P.C.; Konturek, S.J.; Sulekova, Z.; Meixner, H.; Bielanski, W.; Starzynska, T.; Karczewska, E.; Marlicz, K.; Stachura, J.; Hahn, E.G. Expression of hepatocyte growth factor, transforming growth factor alpha, apoptosis related proteins Bax and Bcl-2, and gastrin in human gastric cancer. Aliment. Pharm. 2001, 15, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Pallichankandy, S.; Thayyullathil, F.; Galadari, S. Critical role of H2O2 in mediating sanguinarine-induced apoptosis in prostate cancer cells via facilitating ceramide generation, ERK1/2 phosphorylation, and Par-4 cleavage. Free Radic. Biol. Med. 2019, 134, 527–544. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, S.; Kondo, S.; Shinomura, Y.; Kanayama, S.; Miyazaki, Y.; Kiyohara, T.; Hiraoka, S.; Matsuzawa, Y. Met/HGF receptor modulates bcl-w expression and inhibits apoptosis in human colorectal cancers. Br. J. Cancer 2000, 83, 668–673. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lim, H.M.; Lee, J.; Nam, M.J.; Park, S.-H. Acetylshikonin Induces Apoptosis in Human Colorectal Cancer HCT-15 and LoVo Cells via Nuclear Translocation of FOXO3 and ROS Level Elevation. Oxidative Med. Cell. Longev. 2021, 2021, 6647107. [Google Scholar] [CrossRef]
- Lim, H.M.; Park, S.H.; Nam, M.J. Induction of apoptosis in indole-3-carbinol-treated lung cancer H1299 cells via ROS level elevation. Hum. Exp. Toxicol. 2021, 40, 812–825. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.K.; Lim, H.M.; Park, S.-H.; Nam, M.J. Knockout of Hepatocyte Growth Factor by CRISPR/Cas9 System Induces Apoptosis in Hepatocellular Carcinoma Cells. J. Pers. Med. 2021, 11, 983. https://doi.org/10.3390/jpm11100983
Lee HK, Lim HM, Park S-H, Nam MJ. Knockout of Hepatocyte Growth Factor by CRISPR/Cas9 System Induces Apoptosis in Hepatocellular Carcinoma Cells. Journal of Personalized Medicine. 2021; 11(10):983. https://doi.org/10.3390/jpm11100983
Chicago/Turabian StyleLee, Han Ki, Heui Min Lim, See-Hyoung Park, and Myeong Jin Nam. 2021. "Knockout of Hepatocyte Growth Factor by CRISPR/Cas9 System Induces Apoptosis in Hepatocellular Carcinoma Cells" Journal of Personalized Medicine 11, no. 10: 983. https://doi.org/10.3390/jpm11100983
APA StyleLee, H. K., Lim, H. M., Park, S.-H., & Nam, M. J. (2021). Knockout of Hepatocyte Growth Factor by CRISPR/Cas9 System Induces Apoptosis in Hepatocellular Carcinoma Cells. Journal of Personalized Medicine, 11(10), 983. https://doi.org/10.3390/jpm11100983