Identification of Metabolic Biomarkers in Relation to Methotrexate Response in Early Rheumatoid Arthritis

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Subjects
2.2. Metabolomics Study
2.3. Statistics
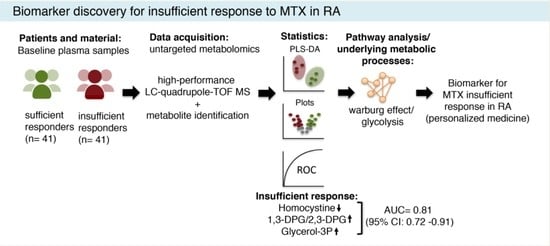
3. Results
3.1. Baseline Comparisons
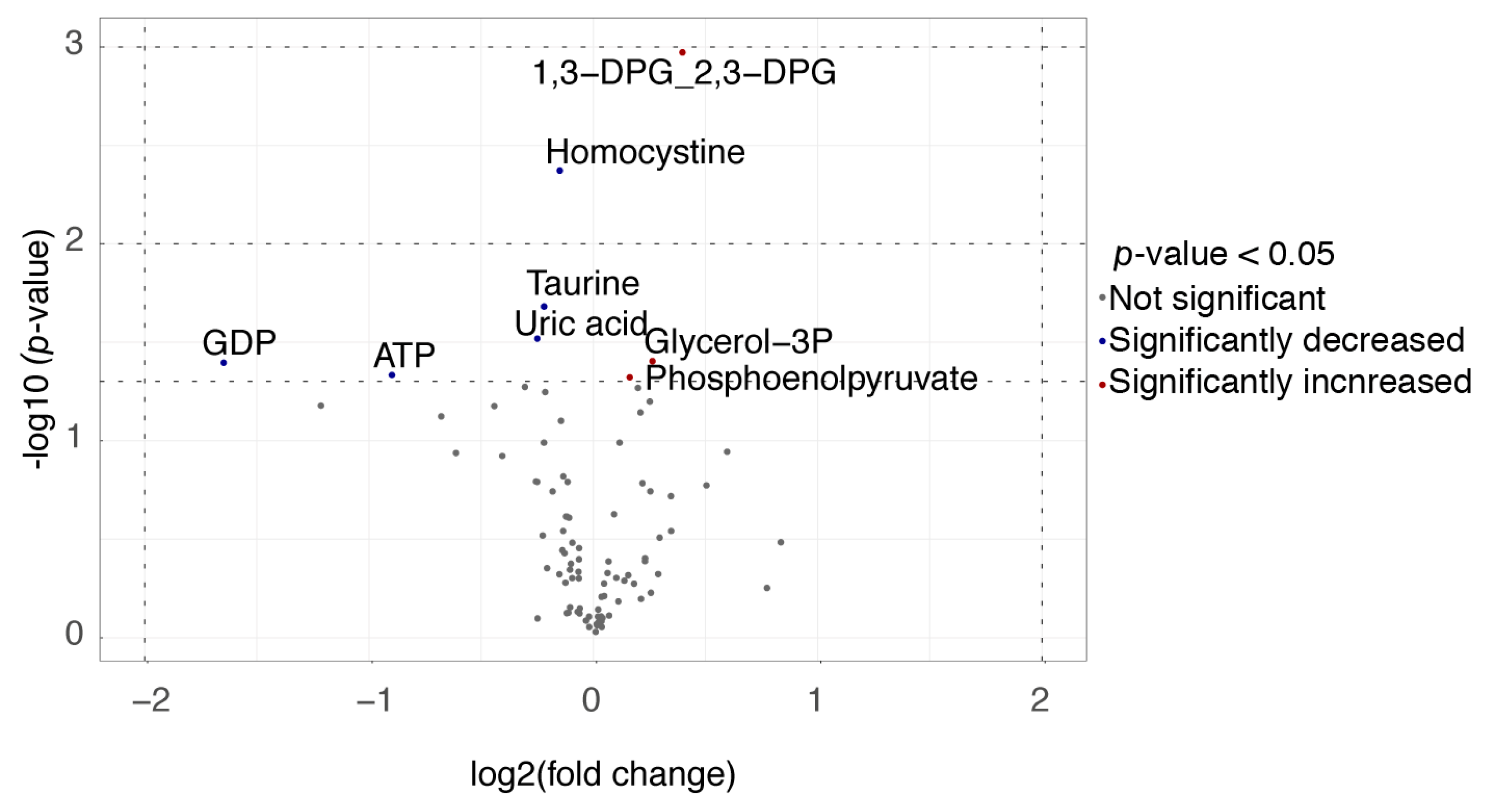
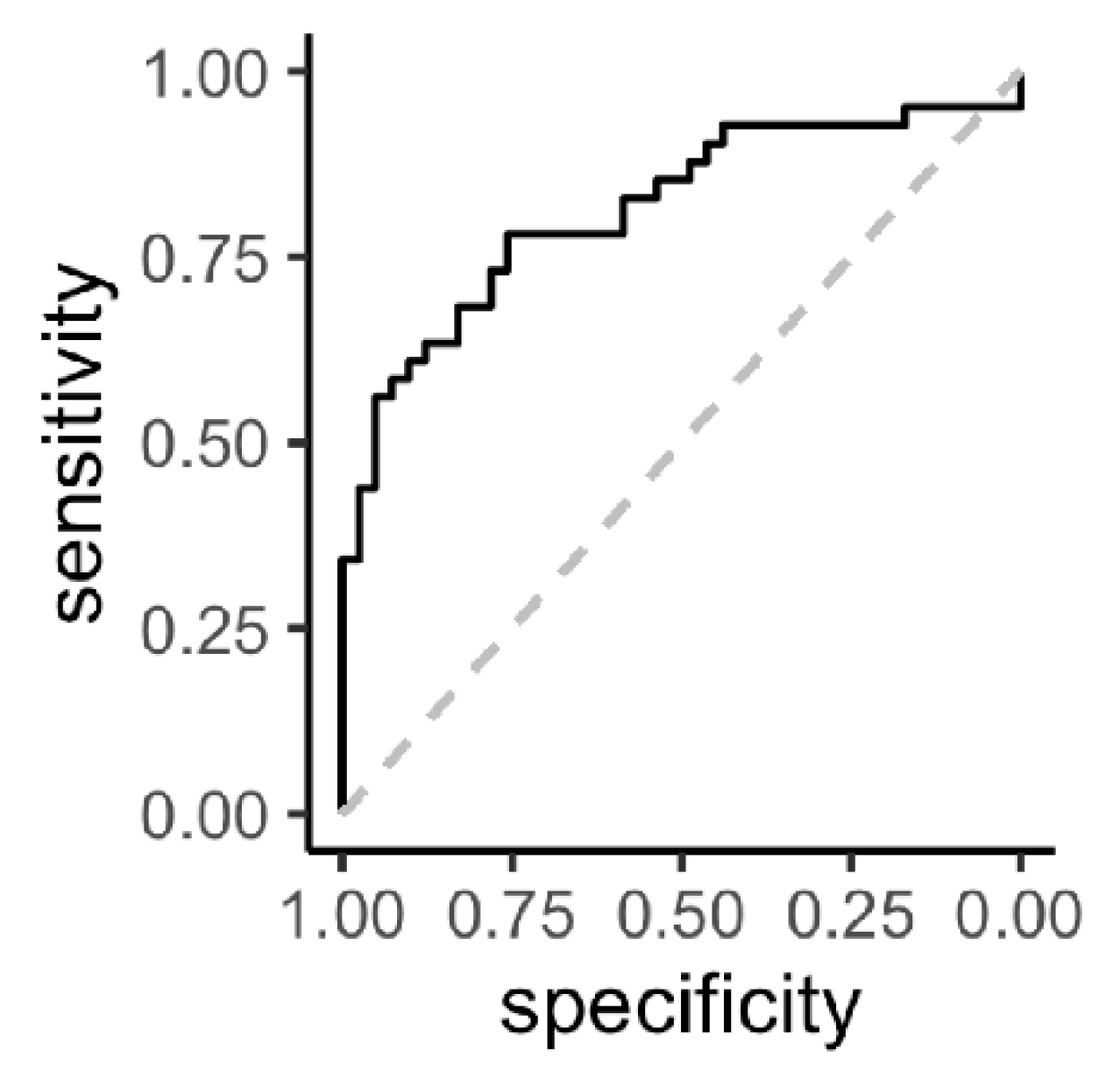
3.2. Metabolite Analysis
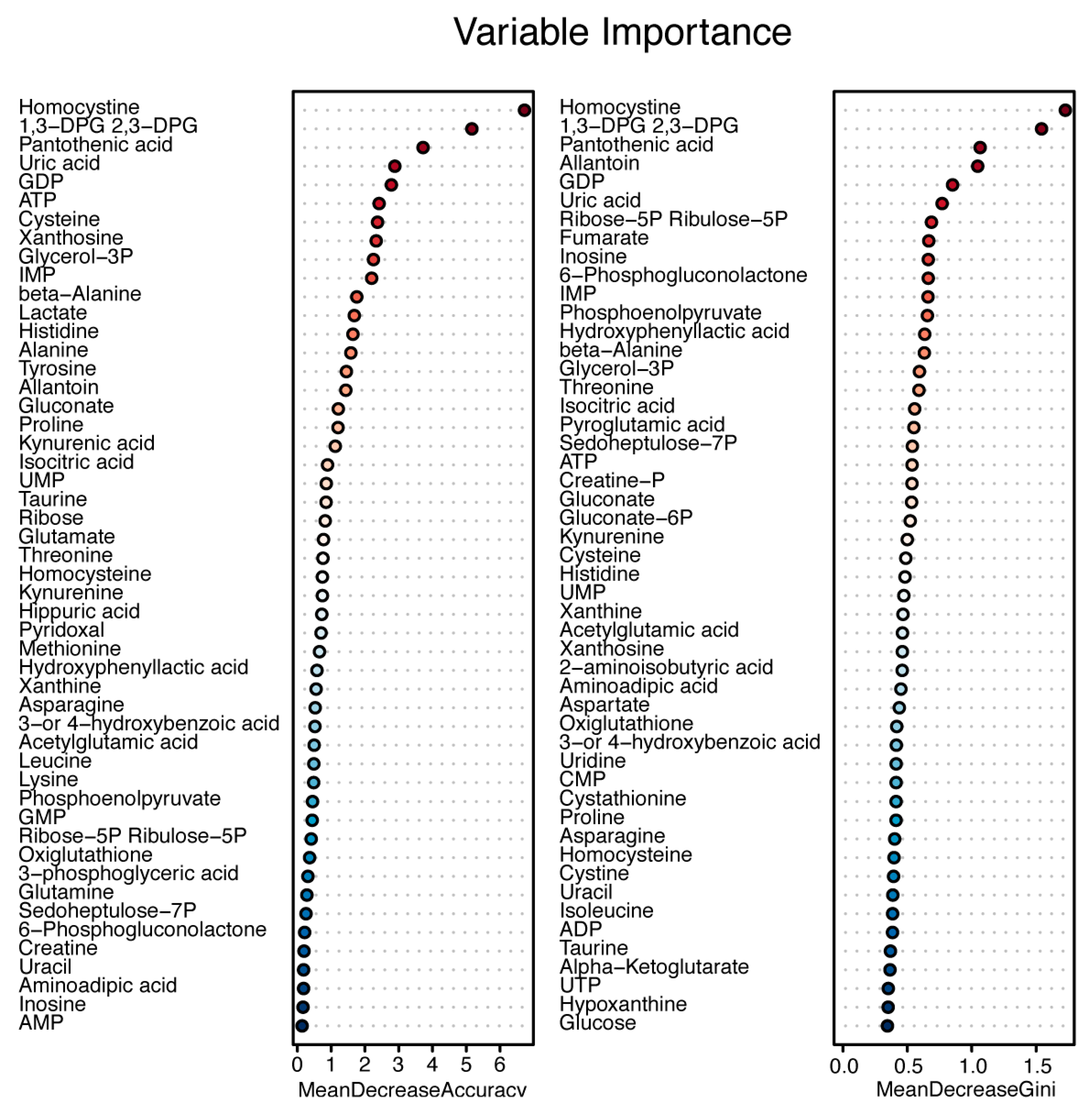
3.3. Random Forest Analysis
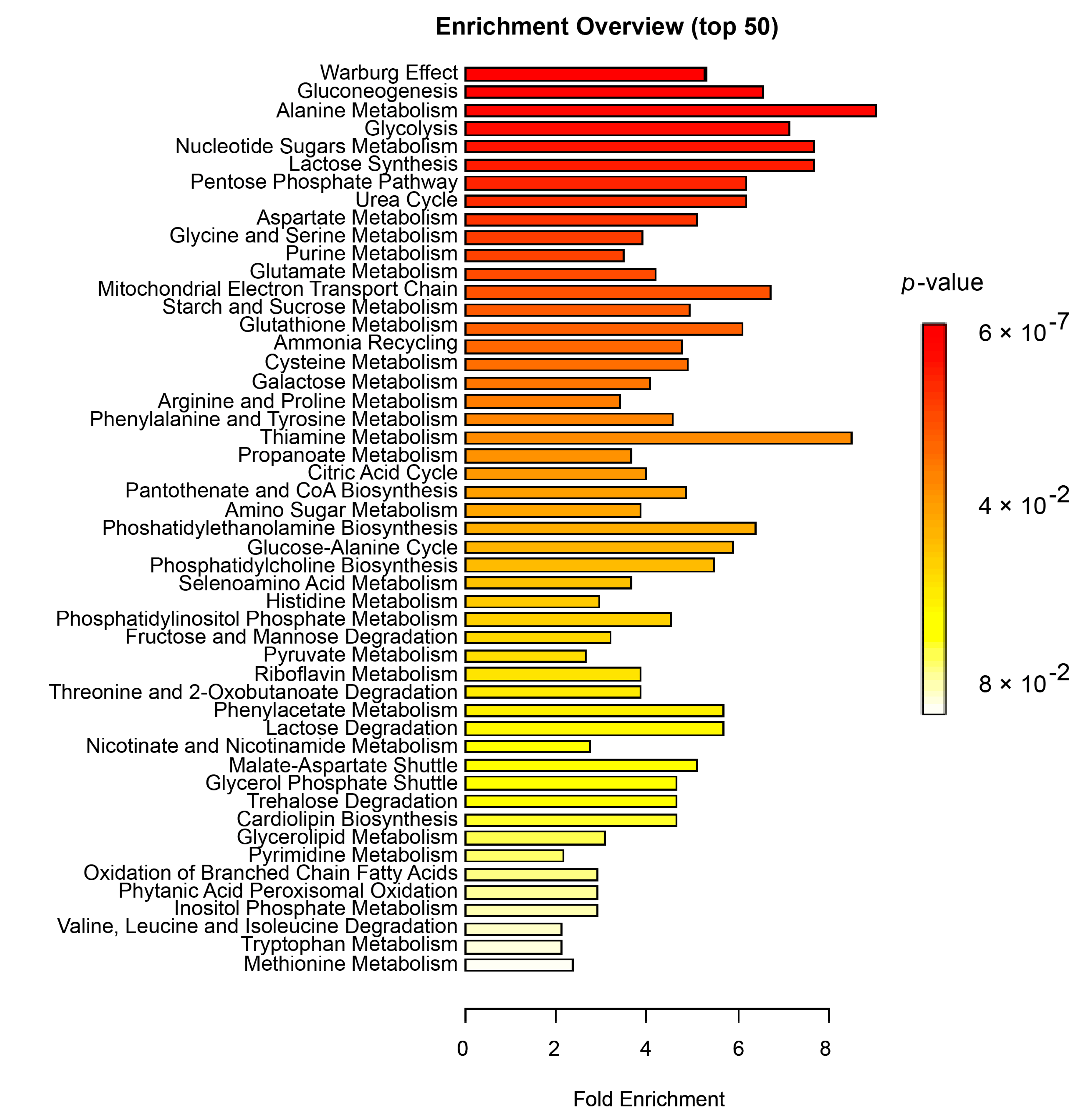
3.4. Enrichment Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smolen, J.S.; Aletaha, D.; McInnes, I.B. Rheumatoid arthritis. Lancet 2016, 388, 2023–2038. [Google Scholar] [CrossRef]
- Smolen, J.S.; Landewé, R.B.M.M.; Bijlsma, J.W.J.J.; Burmester, G.R.; Dougados, M.; Kerschbaumer, A.; McInnes, I.B.; Sepriano, A.; van Vollenhoven, R.F.; De Wit, M.; et al. EULAR recommendations for the management of rheumatoid arthritis with synthetic and biological disease-modifying antirheumatic drugs: 2019 update. Ann. Rheum. Dis. 2020, 79, 685–699. [Google Scholar] [CrossRef] [PubMed]
- Gonen, N.; Assaraf, Y.G. Antifolates in cancer therapy: Structure, activity and mechanisms of drug resistance. Drug Resist. Updat. 2012, 15, 183–210. [Google Scholar] [CrossRef]
- Brown, P.M.; Pratt, A.G.; Isaacs, J.D. Mechanism of action of methotrexate in rheumatoid arthritis, and the search for biomarkers. Nat. Rev. Rheumatol. 2016, 12, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Cronstein, B.N.; Aune, T.M. Methotrexate and its mechanisms of action in inflammatory arthritis. Nat. Rev. Rheumatol. 2020, 16, 145–154. [Google Scholar] [CrossRef]
- Choquette, D.; Bessette, L.; Alemao, E.; Haraoui, B.; Postema, R.; Raynauld, J.P.; Coupal, L. Persistence rates of abatacept and TNF inhibitors used as first or second biologic DMARDs in the treatment of rheumatoid arthritis: 9 years of experience from the Rhumadata® clinical database and registry. Arthritis Res. Ther. 2019, 21, 1–9. [Google Scholar] [CrossRef]
- Harrington, R.; Al Nokhatha, S.A.; Conway, R. Jak inhibitors in rheumatoid arthritis: An evidence-based review on the emerging clinical data. J. Inflamm. Res. 2020, 13, 519–531. [Google Scholar] [CrossRef]
- Singh, S.; Singh, S. JAK-STAT inhibitors: Immersing therapeutic approach for management of rheumatoid arthritis. Int. Immunopharmacol. 2020, 86, 106731. [Google Scholar] [CrossRef]
- Roodenrijs, N.M.T.; van der Goes, M.C.; Welsing, P.M.J.; Tekstra, J.; van Laar, J.M.; Lafeber, F.P.J.G.; Bijlsma, J.W.J.; Jacobs, J.W.G. Is prediction of clinical response to methotrexate in individual rheumatoid arthritis patients possible? A systematic literature review. Jt. Bone Spine 2020, 87, 13–23. [Google Scholar] [CrossRef]
- De Rotte, M.C.; Pluijm, S.M.; De Jong, P.H.; Bulatović-Calasan, M.; Wulffraat, N.; Weel, A.; Lindemans, J.; Hazes, J.; De Jonge, R. Development and validation of a prognostic multivariable model to predict insufficient clinical response to methotrexate in rheumatoid arthritis. PLoS ONE 2018, 13, e0208534. [Google Scholar] [CrossRef]
- Gosselt, H.R.; Verhoeven, M.M.A.; de Rotte, M.C.F.J.; Pluijm, S.M.F.; Muller, I.B.; Jansen, G.; Tekstra, J.; Bulatović-Ćalasan, M.; Heil, S.G.; Lafeber, F.P.J.G.; et al. Validation of a Prognostic Multivariable Prediction Model for Insufficient Clinical Response to Methotrexate in Early Rheumatoid Arthritis and Its Clinical Application in Evidencio. Rheumatol. Ther. 2020, 7, 837–850. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Chen, J.; Hu, C.; Xie, Z.; Li, H.; Wei, S.; Wang, D.; Wen, C.; Xu, G. Exploration of the serum metabolite signature in patients with rheumatoid arthritis using gas chromatography–mass spectrometry. J. Pharm. Biomed. Anal. 2016, 127, 60–67. [Google Scholar] [CrossRef]
- Li, J.; Che, N.; Xu, L.; Zhang, Q.; Wang, Q.; Tan, W.; Zhang, M. LC-MS-based serum metabolomics reveals a distinctive signature in patients with rheumatoid arthritis. Clin. Rheumatol. 2018, 37, 1493–1502. [Google Scholar] [CrossRef]
- Weyand, C.M.; Goronzy, J.J. Immunometabolism in early and late stages of rheumatoid arthritis. Nat. Rev. Rheumatol. 2017, 13, 1–11. [Google Scholar] [CrossRef]
- Okano, T.; Saegusa, J.; Takahashi, S.; Ueda, Y.; Morinobu, A. Immunometabolism in rheumatoid arthritis. Immunol. Med. 2018, 41, 89–97. [Google Scholar] [CrossRef]
- Li, C.; Chen, B.; Fang, Z.; Leng, Y.F.; Wang, D.W.; Chen, F.Q.; Xu, X.; Sun, Z.L. Metabolomics in the development and progression of rheumatoid arthritis: A systematic review. Jt. Bone Spine 2020, 87, 425–430. [Google Scholar] [CrossRef]
- Smolenska, Z.; Smolenski, R.T.; Zdrojewski, Z. Plasma concentrations of amino acid and nicotinamide metabolites in rheumatoid arthritis-potential biomarkers of disease activity and drug treatment. Biomarkers 2016, 21, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Teitsma, X.M.; Yang, W.; Jacobs, J.W.G.; Pethö-Schramm, A.; Borm, M.E.A.; Harms, A.C.; Hankemeier, T.; Van Laar, J.M.; Bijlsma, J.W.J.; Lafeber, F.P.J.G. Baseline metabolic profiles of early rheumatoid arthritis patients achieving sustained drug-free remission after initiating treat-to-target tocilizumab, methotrexate, or the combination: Insights from systems biology 11 Medical and Health Sciences 1103 Cl. Arthritis Res. Ther. 2018, 20, 230. [Google Scholar] [CrossRef]
- Sasaki, C.; Hiraishi, T.; Oku, T.; Okuma, K.; Suzumura, K.; Hashimoto, M.; Ito, H.; Aramori, I.; Hirayama, Y. Metabolomic approach to the exploration of biomarkers associated with disease activity in rheumatoid arthritis. PLoS ONE 2019, 14, e0219400. [Google Scholar] [CrossRef]
- Takahashi, S.; Saegusa, J.; Onishi, A.; Morinobu, A. Biomarkers identified by serum metabolomic analysis to predict biologic treatment response in rheumatoid arthritis patients. Rheumatology 2019, 58, 2153–2161. [Google Scholar] [CrossRef]
- Claessen, S.J.J.; Hazes, J.M.; Huisman, M.A.; Van Zeben, D.; Luime, J.J.; Weel, A.E. Use of risk stratification to target therapies in patients with recent onset arthritis; Design of a prospective randomized multicenter controlled trial. BMC Musculoskelet. Disord. 2009, 10, 71. [Google Scholar] [CrossRef] [PubMed]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O.; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. 2010 Rheumatoid arthritis classification criteria: An American College of Rheumatology/European League Against Rheumatism collaborative initiative. Arthritis Rheum. 2010, 62, 2569–2581. [Google Scholar] [CrossRef] [PubMed]
- De Rotte, M.; De Jong, P.; Pluijm, S.; Bulatović-Ćalasan, M.; Barendregt, P.; van Zeben, D.; van der Lubbe, P. Association of low baseline levels of erythrocyte folate with treatment nonresponse at three months in rheumatoid arthritis patients receiving methotrexate. Arthritis Rheum. 2013, 65, 2803–2813. [Google Scholar] [CrossRef]
- Molenaars, M.; Janssens, G.E.; Williams, E.G.; Jongejan, A.; Lan, J.; Rabot, S.; Joly, F.; Moerland, P.D.; Schomakers, B.V.; Lezzerini, M.; et al. A Conserved Mito-Cytosolic Translational Balance Links Two Longevity Pathways. Cell Metab. 2020, 31, 549–563.e7. [Google Scholar] [CrossRef]
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.-A. mixOmics: An R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef]
- Wickham, H. Programming with ggplot2. In ggplot2; Springer: Cham, Switzerland, 2016; pp. 241–253. [Google Scholar]
- Thévenot, E.A.; Roux, A.; Xu, Y.; Ezan, E.; Junot, C. Analysis of the Human Adult Urinary Metabolome Variations with Age, Body Mass Index, and Gender by Implementing a Comprehensive Workflow for Univariate and OPLS Statistical Analyses. J. Proteome Res. 2015, 14, 3322–3335. [Google Scholar] [CrossRef]
- HMDB Human Metabolome Database. Available online: https://hmdb.ca/ (accessed on 9 December 2020).
- Chong, I.G.; Jun, C.H. Performance of some variable selection methods when multicollinearity is present. Chemom. Intell. Lab. Syst. 2005, 78, 103–112. [Google Scholar] [CrossRef]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinf. 2019, 68. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Z.; Yang, S.; Wang, Y.; Yu, L.; Zhang, B.; Rao, Z.; Gao, J.; Tu, S. 1H NMR-based metabolomic analysis for identifying serum biomarkers to evaluate methotrexate treatment in patients with early rheumatoid arthritis. Exp. Ther. Med. 2012, 4, 165–171. [Google Scholar] [CrossRef]
- Sengupta, S.; Wehbe, C.; Majors, A.K.; Ketterer, M.E.; DiBello, P.M.; Jacobsen, D.W. Relative Roles of Albumin and Ceruloplasmin in the Formation of Homocystine, Homocysteine-Cysteine-mixed Disulfide, and Cystine in Circulation. J. Biol. Chem. 2001, 276, 46896–46904. [Google Scholar] [CrossRef] [PubMed]
- Van Ede, A.E.; Laan, R.F.J.M.; Blom, H.J.; Boers, G.H.J.; Haagsma, C.J.; Thomas, C.M.G.; De Boo, T.M.; Van De Putte, L.B.A. Homocysteine and folate status in methotrexate-treated patients with rheumatoid arthritis. Rheumatology 2002, 41, 658–665. [Google Scholar] [CrossRef] [PubMed]
- Shu, J.; Sun, X.; Li, J.; Li, F.; Tang, J.; Shi, L. Serum homocysteine levels and their association with clinical characteristics of inflammatory arthritis. Clin. Rheumatol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Killeen, M.E.; Ferris, L.; Kupetsky, E.A.; Falo, L.; Mathers, A.R. Signaling through Purinergic Receptors for ATP Induces Human Cutaneous Innate and Adaptive Th17 Responses: Implications in the Pathogenesis of Psoriasis. J. Immunol. 2013, 190, 4324–4336. [Google Scholar] [CrossRef]
- da Silva, J.L.G.; Passos, D.F.; Bernardes, V.M.; Leal, D.B.R. ATP and adenosine: Role in the immunopathogenesis of rheumatoid arthritis. Immunol. Lett. 2019, 214, 55–64. [Google Scholar] [CrossRef]
- Chandrupatla, D.M.S.H.; Molthoff, C.F.M.; Ritsema, W.I.G.R.; Vos, R.; Elshof, E.; Matsuyama, T.; Low, P.S.; Musters, R.J.P.; Hammond, A.; Windhorst, A.D.; et al. Prophylactic and therapeutic activity of alkaline phosphatase in arthritic rats: Single-agent effects of alkaline phosphatase and synergistic effects in combination with methotrexate. Transl. Res. 2018, 199, 24–38. [Google Scholar] [CrossRef]
- Peres, R.S.; Liew, F.Y.; Talbot, J.; Carregaro, V.; Oliveira, R.D.; Almeida, S.L.; França, R.F.O.; Donate, P.B.; Pinto, L.G.; Ferreira, F.I.S.; et al. Low expression of CD39 on regulatory T cells as a biomarker for resistance to methotrexate therapy in rheumatoid arthritis. Proc. Natl. Acad. Sci. USA 2015, 112, 2509–2514. [Google Scholar] [CrossRef]
- Gupta, V.; Katiyar, S.; Singh, A.; Misra, R.; Aggarwal, A. CD39 positive regulatory T cell frequency as a biomarker of treatment response to methotrexate in rheumatoid arthritis. Int. J. Rheum. Dis. 2018, 21, 1548–1556. [Google Scholar] [CrossRef]
- Lakomek, M.; Neubauer, B.; Lühe, A.V.D.; Hoch, G.; Schröter, W.; Winkler, H. Erythrocyte pyruvate kinase deficiency: Relations of residual enzyme activity, altered regulation of defective enzymes and concentrations of high-energy phosphates with the severity of clinical manifestation. Eur. J. Haematol. 1992, 49, 82–92. [Google Scholar] [CrossRef]
- Al-Samkari, H.; van Beers, E.J.; Kuo, K.H.M.; Barcellini, W.; Bianchi, P.; Glenthøj, A.B.; Mañú-Pereira, M.; van Wijk, R.; Glader, B.; Grace, R.F. The variable manifestations of disease in pyruvate kinase deficiency and their management. Haematologica 2020, 105. [Google Scholar] [CrossRef]
- Staal, G.E.J.; Jansen, G.; Roos, D. Pyruvate kinase and the “high ATP syndrome”. J. Clin. Investig. 1984, 74, 231–235. [Google Scholar] [CrossRef]
- Weyand, C.M.; Goronzy, J.J. Immunometabolism in the development of rheumatoid arthritis. Immunol. Rev. 2020, 294, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Zaal, E.A.; Berkers, C.R. The influence of metabolism on drug response in cancer. Front. Oncol. 2018, 8, 500. [Google Scholar] [CrossRef] [PubMed]
- Palsson-Mcdermott, E.M.; O’Neill, L.A.J. The Warburg effect then and now: From cancer to inflammatory diseases. BioEssays 2013, 35, 965–973. [Google Scholar] [CrossRef]
- Heiden, M.G.V.; Cantley, L.C.; Thompson, C.B. Understanding the warburg effect: The metabolic requirements of cell proliferation. Science (80-) 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Carbonell, R.; Divakaruni, A.S.; Lodi, A.; Vicente-Suarez, I.; Saha, A.; Cheroutre, H.; Boss, G.R.; Tiziani, S.; Murphy, A.N.; Guma, M. Critical Role of Glucose Metabolism in Rheumatoid Arthritis Fibroblast-like Synoviocytes. Arthritis Rheumatol. 2016, 68, 1614–1626. [Google Scholar] [CrossRef] [PubMed]
- Rezaei, R.; Tahmasebi, S.; Atashzar, M.R.; Amani, D. Glycolysis and Autoimmune Diseases: A Growing Relationship. Biochem. Suppl. Ser. A Membr. Cell Biol. 2020, 14, 91–106. [Google Scholar] [CrossRef]
- Blits, M.; Jansen, G.; Assaraf, Y.G.; Van De Wiel, M.A.; Lems, W.F.; Nurmohamed, M.T.; Van Schaardenburg, D.; Voskuyl, A.E.; Wolbink, G.J.; Vosslamber, S.; et al. Methotrexate normalizes up-regulated folate pathway genes in rheumatoid arthritis. Arthritis Rheum. 2013, 65, 2791–2802. [Google Scholar] [CrossRef] [PubMed]
- Kornberg, M.D. The immunologic Warburg effect: Evidence and therapeutic opportunities in autoimmunity. Wiley Interdiscip. Rev. Syst. Biol. Med. 2020, e1486. [Google Scholar] [CrossRef]
- Pålsson-McDermott, E.M.; O’Neill, L.A.J. Targeting immunometabolism as an anti-inflammatory strategy. Cell Res. 2020, 30, 300–314. [Google Scholar] [CrossRef]
- Piranavan, P.; Bhamra, M.; Perl, A. Metabolic Targets for Treatment of Autoimmune Diseases. Immunometabolism 2020, 2, e200012. [Google Scholar] [CrossRef]
- Rhoads, J.P.; Major, A.S.; Rathmell, J.C. Fine tuning of immunometabolism for the treatment of rheumatic diseases. Nat. Rev. Rheumatol. 2017, 13, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Spurlock, C.F.; Tossberg, J.T.; Matlock, B.K.; Olsen, N.J.; Aune, T.M. Methotrexate inhibits NF-κB activity via long intergenic (noncoding) RNA-p21 induction. Arthritis Rheumatol. 2014, 66, 2947–2957. [Google Scholar] [CrossRef] [PubMed]
- Coras, R.; Murillo-Saich, J.; Guma, M. Circulating Pro- and Anti-Inflammatory Metabolites and Its Potential Role in Rheumatoid Arthritis Pathogenesis. Cells 2020, 9, 827. [Google Scholar] [CrossRef]
- Lee, J.J.; Bykerk, V.P.; Dresser, G.K.; Boire, G.; Haraoui, B.; Hitchon, C.; Thorne, C.; Tin, D.; Jamal, S.; Keystone, E.C.; et al. Reduction in serum uric acid may be related to methotrexate efficacy in early rheumatoid arthritis: Data from the Canadian Early Arthritis Cohort (CATCH). Clin. Med. Insights Arthritis Musculoskelet. Disord. 2016, 9, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Ling, S.; Bluett, J.; Barton, A. Prediction of response to methotrexate in rheumatoid arthritis. Expert Rev. Clin. Immunol. 2018, 14, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Szostak, B.; Machaj, F.; Rosik, J.; Pawlik, A. Using pharmacogenetics to predict methotrexate response in rheumatoid arthritis patients. Expert Opin. Drug Metab. Toxicol. 2020, 16, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Sergeant, J.C.; Hyrich, K.L.; Anderson, J.; Kopec-Harding, K.; Hope, H.F.; Symmons, D.P.M.M.; Barton, A.; Verstappen, S.M.M.M. Prediction of primary non-response to methotrexate therapy using demographic, clinical and psychosocial variables: Results from the UK Rheumatoid Arthritis Medication Study (RAMS). Arthritis Res. Ther. 2018, 20, 147. [Google Scholar] [CrossRef]
- Eektimmerman, F.; Allaart, C.F.; Hazes, J.M.; Den Broeder, A.A.; Fransen, J.; Swen, J.J.; Guchelaar, H.-J.J. Validation of a clinical pharmacogenetic model to predict methotrexate nonresponse in rheumatoid arthritis patients. Pharmacogenomics 2019, 20, 85–93. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insufficient Responders (DAS28-ESR > 3.2) N = 41 | Sufficient Responders (DAS28-ESR ≤ 3.2) N = 41 | p-Value | |
|---|---|---|---|
| Baseline DAS28, mean ± SD | 4.3 ± 1.3 | 5.6 ± 1.0 | <0.001 |
| Age, mean ± SD | 50.0 ± 11.9 | 52.6 ± 16.9 | 0.41 |
| Sex, Male, N (%) | 8 (20) | 15 (37) | 0.14 |
| BMI (kg/m2), mean ± SD | 28.1 ± 5.4 | 24.3 ± 4.1 # | <0.001 |
| RF positive, N (%) | 26 (63) | 33 (80) | 0.14 |
| ACPA positive, N (%) | 25 (61) | 31 (76) | 0.24 |
| Treatment | |||
| MTX + SSZ + HCQ + corticosteroids i.m. | 8 (20) | 15 (37) | 0.14 |
| MTX + SSZ + HCQ + corticosteroids per os | 11 (27) | 15 (37) | 0.48 |
| MTX + corticosteroids per os | 13 (32) | 7 (17) | 0.20 |
| MTX | 9 (22) | 4 (10) | 0.23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gosselt, H.R.; Muller, I.B.; Jansen, G.; van Weeghel, M.; Vaz, F.M.; Hazes, J.M.W.; Heil, S.G.; de Jonge, R. Identification of Metabolic Biomarkers in Relation to Methotrexate Response in Early Rheumatoid Arthritis. J. Pers. Med. 2020, 10, 271. https://doi.org/10.3390/jpm10040271
Gosselt HR, Muller IB, Jansen G, van Weeghel M, Vaz FM, Hazes JMW, Heil SG, de Jonge R. Identification of Metabolic Biomarkers in Relation to Methotrexate Response in Early Rheumatoid Arthritis. Journal of Personalized Medicine. 2020; 10(4):271. https://doi.org/10.3390/jpm10040271
Chicago/Turabian StyleGosselt, Helen R., Ittai B. Muller, Gerrit Jansen, Michel van Weeghel, Frédéric M. Vaz, Johanna M. W. Hazes, Sandra G. Heil, and Robert de Jonge. 2020. "Identification of Metabolic Biomarkers in Relation to Methotrexate Response in Early Rheumatoid Arthritis" Journal of Personalized Medicine 10, no. 4: 271. https://doi.org/10.3390/jpm10040271
APA StyleGosselt, H. R., Muller, I. B., Jansen, G., van Weeghel, M., Vaz, F. M., Hazes, J. M. W., Heil, S. G., & de Jonge, R. (2020). Identification of Metabolic Biomarkers in Relation to Methotrexate Response in Early Rheumatoid Arthritis. Journal of Personalized Medicine, 10(4), 271. https://doi.org/10.3390/jpm10040271