
Telomere Length and Genetic Variations in Acquired Pediatric Aplastic Anemia: A Flow-FISH Study in Korean Patients

 , , , , , , , and
, , , , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample
2.2. Telomere Length Measurement Using Flow-FISH
2.3. Genetic Evaluation
2.4. Statistical Analysis
3. Results
3.1. Patients
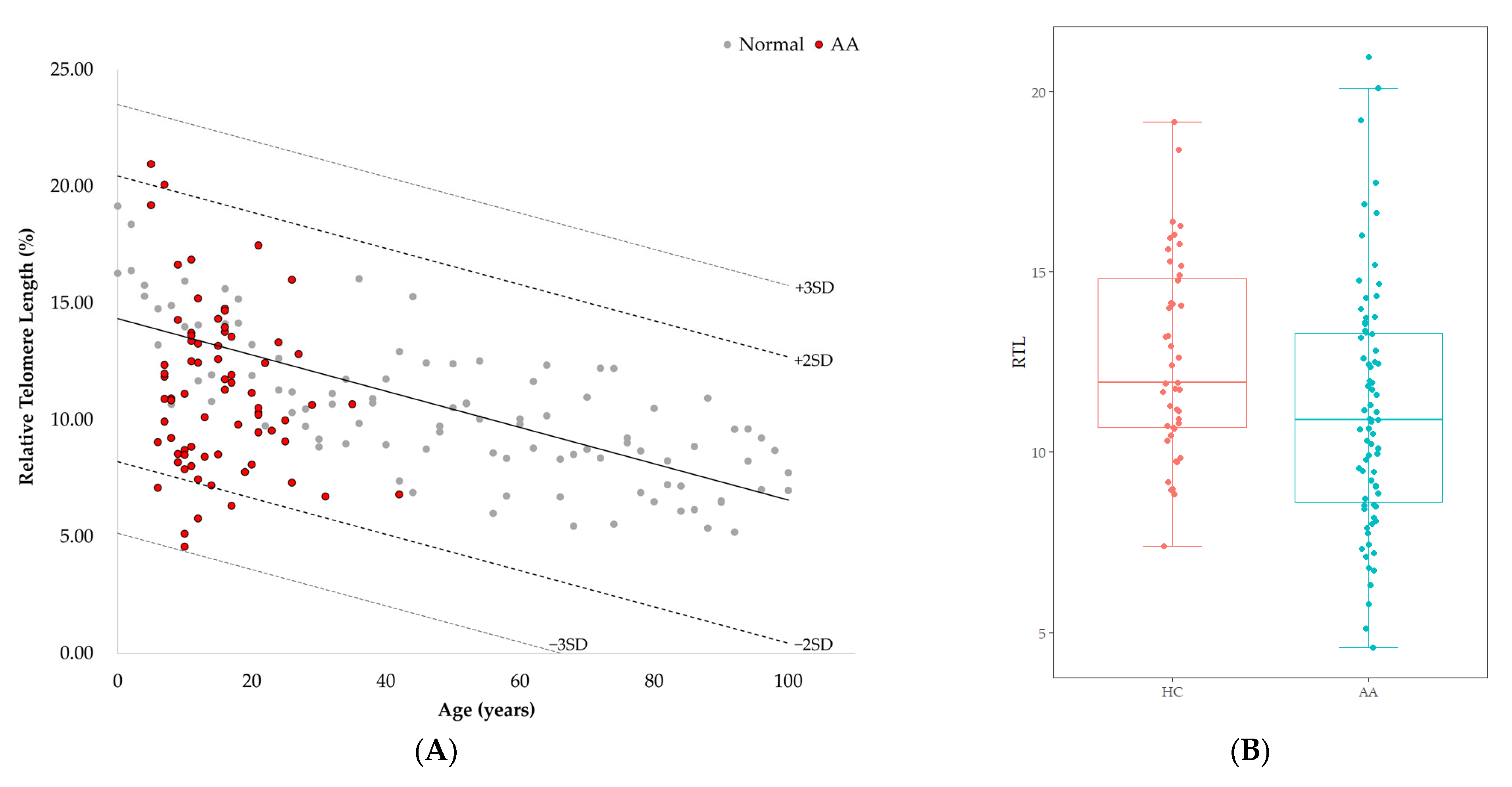
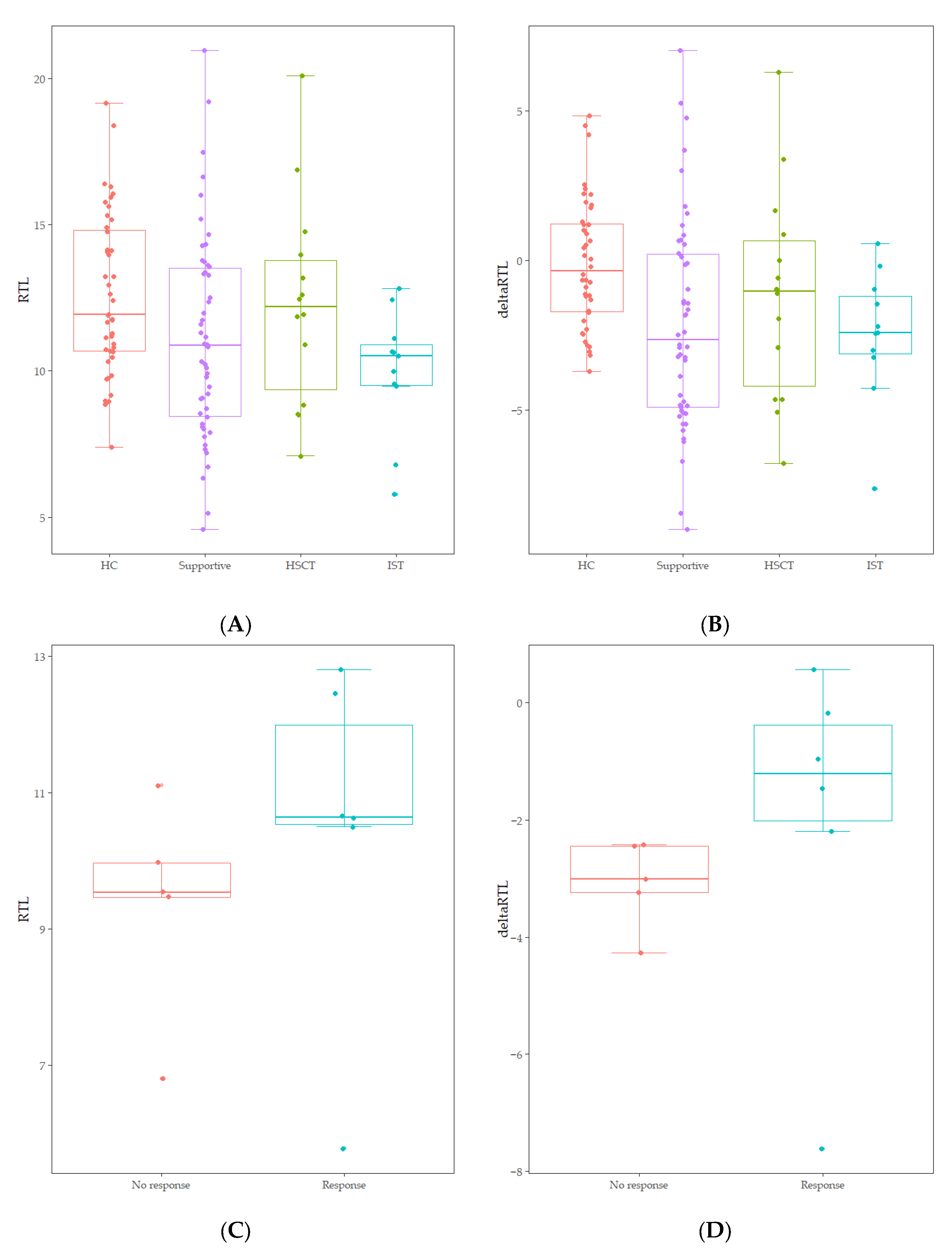
3.2. Telomere Length of Patients with AA
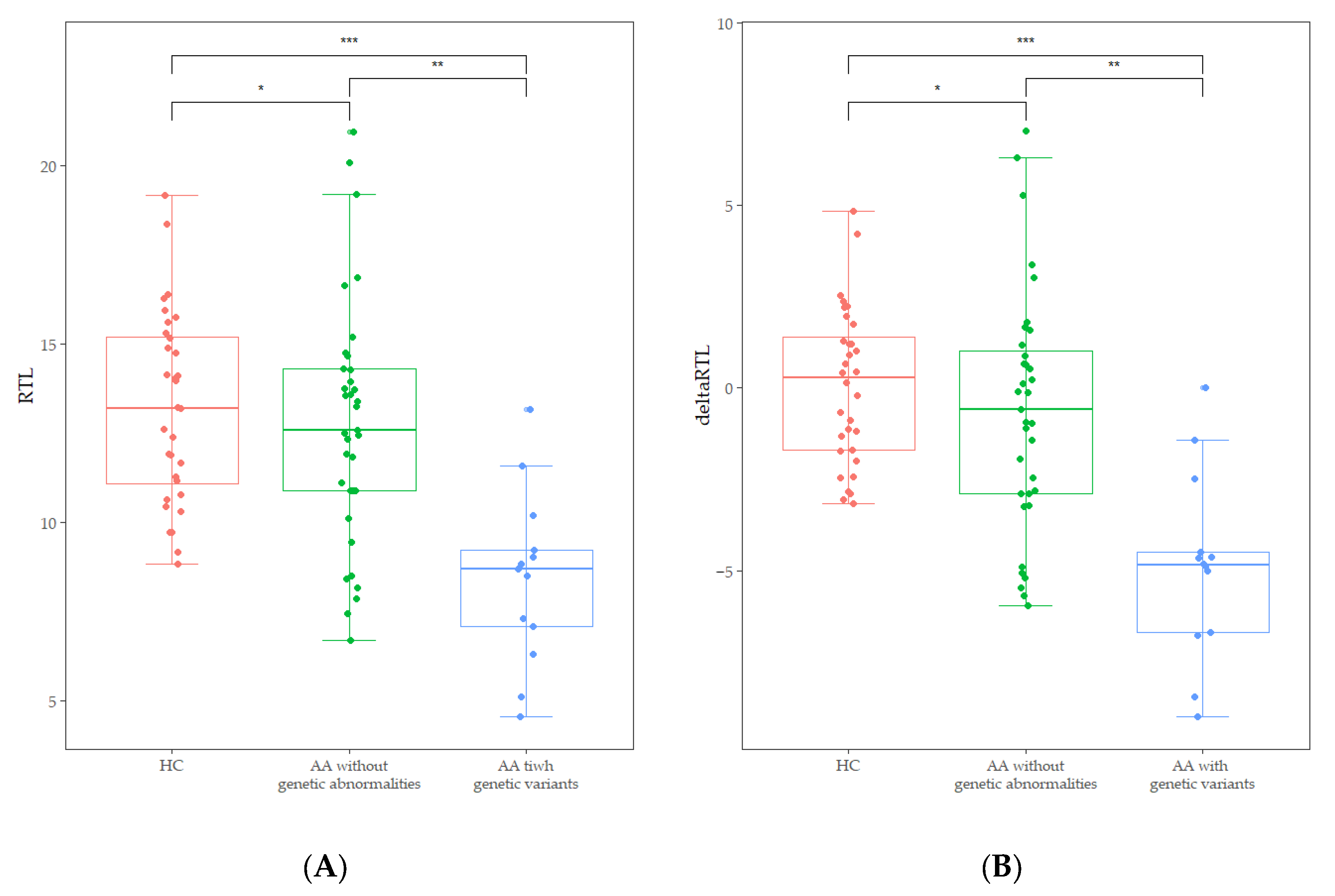
3.3. Telomere Length and Genetic Variants
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Young, N.S.; Kaufman, D.W. The epidemiology of acquired aplastic anemia. Haematologica 2008, 93, 489–492. [Google Scholar] [CrossRef] [PubMed]
- Shimamura, A.; Alter, B.P. Pathophysiology and management of inherited bone marrow failure syndromes. Blood Rev. 2010, 24, 101–122. [Google Scholar] [CrossRef] [PubMed]
- Shimano, K.A.; Narla, A.; Rose, M.J.; Gloude, N.J.; Allen, S.W.; Bergstrom, K.; Broglie, L.; Carella, B.A.; Castillo, P.; de Jong, J.L.O.; et al. Diagnostic work-up for severe aplastic anemia in children: Consensus of the North American Pediatric Aplastic Anemia Consortium. Am. J. Hematol. 2021, 96, 1491–1504. [Google Scholar] [PubMed]
- Jeong, D.C.; Chung, N.G.; Kang, H.J.; Koo, H.H.; Kook, H.; Kim, S.K.; Kim, S.Y.; Kim, H.S.; Kim, H.M.; Park, K.D.; et al. Epidemiology and Clinical Long-term Outcome of Childhood Aplastic Anemia in Korea for 15 Years: Retrospective Study of the Korean Society of Pediatric Hematology Oncology (KSPHO). J. Pediatr. Hematol./Oncol. 2011, 33, 172–178. [Google Scholar]
- Dokal, I.; Tummala, H.; Vulliamy, T. Inherited bone marrow failure in the pediatric patient. Blood 2022, 140, 556–570. [Google Scholar]
- Park, J.; Kim, M.; Jang, W.; Chae, H.; Kim, Y.; Chung, N.G.; Lee, J.W.; Cho, B.; Jeong, D.C.; Park, I.Y.; et al. Founder haplotype analysis of Fanconi anemia in the Korean population finds common ancestral haplotypes for a FANCG variant. Ann. Hum. Genet. 2015, 79, 153–161. [Google Scholar] [CrossRef]
- Kook, H.; Chung, N.G.; Kang, H.J.; Im, H.J. Acquired aplastic anemia in Korean children: Treatment guidelines from the Bone Marrow Failure Committee of the Korean Society of Pediatric Hematology Oncology. Int. J. Hematol. 2016, 103, 380–386. [Google Scholar]
- Lee, J.M.; Kim, H.S.; Yoo, J.; Lee, J.; Ahn, A.; Cho, H.; Han, E.H.; Jung, J.; Yoo, J.W.; Kim, S.; et al. Genomic insights into inherited bone marrow failure syndromes in a Korean population. Br. J. Haematol. 2024, 205, 1581–1589. [Google Scholar] [CrossRef]
- Peslak, S.A.; Olson, T.; Babushok, D.V. Diagnosis and Treatment of Aplastic Anemia. Curr. Treat. Options Oncol. 2017, 18, 70. [Google Scholar]
- Yoshida, N. Recent advances in the diagnosis and treatment of pediatric acquired aplastic anemia. Int. J. Hematol. 2024, 119, 240–247. [Google Scholar]
- Baerlocher, G.M.; Vulto, I.; de Jong, G.; Lansdorp, P.M. Flow cytometry and FISH to measure the average length of telomeres (flow FISH). Nat. Protoc. 2006, 1, 2365–2376. [Google Scholar] [CrossRef] [PubMed]
- Aubert, G.; Baerlocher, G.M.; Vulto, I.; Poon, S.S.; Lansdorp, P.M. Collapse of telomere homeostasis in hematopoietic cells caused by heterozygous mutations in telomerase genes. PLoS Genet. 2012, 8, e1002696. [Google Scholar] [CrossRef] [PubMed]
- Holohan, B.; Wright, W.E.; Shay, J.W. Cell biology of disease: Telomeropathies: An emerging spectrum disorder. J. Cell Biol. 2014, 205, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Kook, H.; Chung, I.J.; Na, J.A.; Park, M.R.; Hwang, T.J.; Kwak, J.Y.; Sohn, S.K.; Kim, H.J. Telomere length changes in patients with aplastic anaemia. Br. J. Haematol. 2001, 112, 1025–1030. [Google Scholar] [CrossRef]
- Edison, E.S.; Barade, A.; Korula, A.; Nisham, P.N.; Devasia, A.J.; Lakshmi, K.M.; Prakash, K.U.; Abraham, A.; Srivastava, A.; Mathews, V.; et al. Telomere Length Is Significantly Reduced in Patients with Aplastic Anemia and Is Not Influenced By Gender and Severity. Blood 2017, 130, 4964. [Google Scholar]
- Sakaguchi, H.; Nishio, N.; Hama, A.; Kawashima, N.; Wang, X.; Narita, A.; Doisaki, S.; Xu, Y.; Muramatsu, H.; Yoshida, N.; et al. Peripheral blood lymphocyte telomere length as a predictor of response to immunosuppressive therapy in childhood aplastic anemia. Haematologica 2014, 99, 1312–1316. [Google Scholar] [CrossRef]
- Chung, N.G.; Kim, M. Current insights into inherited bone marrow failure syndromes. Korean J. Pediatr. 2014, 57, 337–344. [Google Scholar] [CrossRef]
- Park, S.Y.; Lee, J.M.; Kim, M.J.; Chung, N.G.; Lee, J.B.; Kim, Y.; Kim, M. Validation of Pathogenicity of Gene Variants in Fanconi Anemia Using Patient-derived Dermal Fibroblasts. Ann. Lab. Med. 2023, 43, 127–131. [Google Scholar] [CrossRef]
- Alix, T.; Chery, C.; Josse, T.; Bronowicki, J.P.; Feillet, F.; Gueant-Rodriguez, R.M.; Namour, F.; Gueant, J.L.; Oussalah, A. Predictors of the utility of clinical exome sequencing as a first-tier genetic test in patients with Mendelian phenotypes: Results from a referral center study on 603 consecutive cases. Hum. Genom. 2023, 17, 5. [Google Scholar] [CrossRef]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–424. [Google Scholar] [CrossRef]
- Doney, K.; Leisenring, W.; Storb, R.; Appelbaum, F.R. Primary treatment of acquired aplastic anemia: Outcomes with bone marrow transplantation and immunosuppressive therapy. Ann. Intern. Med. 1997, 126, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Meyne, J.; Ratliff, R.L.; Moyzis, R.K. Conservation of the Human Telomere Sequence (Ttaggg)N among Vertebrates. Proc. Natl. Acad. Sci. USA 1989, 86, 7049–7053. [Google Scholar] [PubMed]
- Savage, S.A. Human telomeres and telomere biology disorders. Prog. Mol. Biol. Transl. Sci. 2014, 125, 41–66. [Google Scholar] [PubMed]
- Montpetit, A.J.; Alhareeri, A.A.; Montpetit, M.; Starkweather, A.R.; Elmore, L.W.; Filler, K.; Mohanraj, L.; Burton, C.W.; Menzies, V.S.; Lyon, D.E.; et al. Telomere length: A review of methods for measurement. Nurs. Res. 2014, 63, 289–299. [Google Scholar]
- Harley, C.B.; Futcher, A.B.; Greider, C.W. Telomeres shorten during ageing of human fibroblasts. Nature 1990, 345, 458–460. [Google Scholar]
- Cawthon, R.M. Telomere length measurement by a novel monochrome multiplex quantitative PCR method. Nucleic Acids Res. 2009, 37, e21. [Google Scholar]
- Rufer, N.; Dragowska, W.; Thornbury, G.; Roosnek, E.; Lansdorp, P.M. Telomere length dynamics in human lymphocyte subpopulations measured by flow cytometry. Nat. Biotechnol. 1998, 16, 743–747. [Google Scholar]
- Lai, T.P.; Wright, W.E.; Shay, J.W. Comparison of telomere length measurement methods. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2018, 373, 20160451. [Google Scholar] [CrossRef]
- Gutierrez-Rodrigues, F.; Santana-Lemos, B.A.; Scheucher, P.S.; Alves-Paiva, R.M.; Calado, R.T. Direct comparison of flow-FISH and qPCR as diagnostic tests for telomere length measurement in humans. PLoS ONE 2014, 9, e113747. [Google Scholar]
- Barade, A.; Lakshmi, K.M.; Korula, A.; Abubacker, F.N.; Kulkarni, U.P.; Abraham, A.; Mathews, V.; George, B.; Edison, E.S. Comparison of telomere length in patients with bone marrow failure syndromes and healthy controls. Eur. J. Haematol. 2024, 112, 810–818. [Google Scholar]
- Scheinberg, P.; Cooper, J.N.; Sloand, E.M.; Wu, C.O.; Calado, R.T.; Young, N.S. Association of Telomere Length of Peripheral Blood Leukocytes With Hematopoietic Relapse, Malignant Transformation, and Survival in Severe Aplastic Anemia. JAMA J. Am. Med. Assoc. 2010, 304, 1358–1364. [Google Scholar] [CrossRef] [PubMed]
- Barade, A.; Aboobacker, F.; Korula, A.; Lakshmi, K.; Devasia, A.; Abraham, A.; Mathews, V.; Edison, E.; George, B. Impact of donor telomere length on survival in patients undergoing matched sibling donor transplantation for aplastic anaemia. Br. J. Haematol. 2022, 196, 724–734. [Google Scholar] [CrossRef] [PubMed]
- Gadalla, S.M.; Wang, T.; Haagenson, M.; Spellman, S.R.; Lee, S.J.; Williams, K.M.; Wong, J.Y.; De Vivo, I.; Savage, S.A. Association between donor leukocyte telomere length and survival after unrelated allogeneic hematopoietic cell transplantation for severe aplastic anemia. JAMA 2015, 313, 594–602. [Google Scholar] [PubMed]
- Gadalla, S.M.; Aubert, G.; Wang, T.; Haagenson, M.; Spellman, S.R.; Wang, L.X.; Katki, H.A.; Savage, S.A.; Lee, S.J. Donor telomere length and causes of death after unrelated hematopoietic cell transplantation in patients with marrow failure. Blood 2018, 131, 2393–2398. [Google Scholar]
- Alsaggaf, R.; Katta, S.; Wang, T.; Hicks, B.D.; Zhu, B.; Spellman, S.R.; Lee, S.J.; Horvath, S.; Gadalla, S.M. Epigenetic Aging and Hematopoietic Cell Transplantation in Patients With Severe Aplastic Anemia. Transplant. Cell. Ther. 2021, 27, 313.e311–313.e318. [Google Scholar] [CrossRef]
- Przychodzen, B.; Makishima, H.; Sekeres, M.A.; Balasubramanian, S.K.; Thota, S.; Patel, B.J.; Clemente, M.; Hirsch, C.; Dienes, B.; Maciejewski, J.P. Fanconi Anemia germline variants as susceptibility factors in aplastic anemia, MDS and AML. Oncotarget 2018, 9, 2050–2057. [Google Scholar]
- Nie, D.; Zhang, J.; Wang, F.; Li, X.; Liu, L.; Zhang, W.; Cao, P.; Chen, X.; Zhang, Y.; Chen, J.; et al. Fanconi anemia gene-associated germline predisposition in aplastic anemia and hematologic malignancies. Front. Med. 2022, 16, 459–466. [Google Scholar]
- Gadalla, S.M.; Cawthon, R.; Giri, N.; Alter, B.P.; Savage, S.A. Telomere length in blood, buccal cells, and fibroblasts from patients with inherited bone marrow failure syndromes. Aging 2010, 2, 867–874. [Google Scholar]
- Alder, J.K.; Cogan, J.D.; Brown, A.F.; Anderson, C.J.; Lawson, W.E.; Lansdorp, P.M.; Phillips, J.A., 3rd; Loyd, J.E.; Chen, J.J.; Armanios, M. Ancestral mutation in telomerase causes defects in repeat addition processivity and manifests as familial pulmonary fibrosis. PLoS Genet. 2011, 7, e1001352. [Google Scholar]
- Arias-Salgado, E.G.; Galvez, E.; Planas-Cerezales, L.; Pintado-Berninches, L.; Vallespin, E.; Martinez, P.; Carrillo, J.; Iarriccio, L.; Ruiz-Llobet, A.; Catala, A.; et al. Genetic analyses of aplastic anemia and idiopathic pulmonary fibrosis patients with short telomeres, possible implication of DNA-repair genes. Orphanet. J. Rare Dis. 2019, 14, 82. [Google Scholar]
- van der Vis, J.J.; van der Smagt, J.J.; van Batenburg, A.A.; Goldschmeding, R.; van Es, H.W.; Grutters, J.C.; van Moorsel, C.H.M. Pulmonary fibrosis in non-mutation carriers of families with short telomere syndrome gene mutations. Respirology 2021, 26, 1160–1170. [Google Scholar] [CrossRef] [PubMed]
- Giudice, V.; Selleri, C. Aplastic anemia: Pathophysiology. Semin. Hematol. 2022, 59, 13–20. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Patient Subgroups | Number (%) | Age, Years * | Age at Diagnosis, Years | Sex, Male | ANC (×109/L) | Hb (g/dL) | Platelet (×109/L) | RTL | Delta RTL |
|---|---|---|---|---|---|---|---|---|---|
| Total | 75 (100) | 13 (5–42) | 9 (1–17) | 43 | 0.92 (0.05–1.94) | 9.7 (4.7–12.7) | 36.0 (5–84) | 10.89 (4.57–19.20) | −0.34 (−3.69–4.83) |
| Severity | |||||||||
| NSAA | 51 (68.0) | 14 (5–31) | 9 (2–16) | 29 | 1.10 (0.08–1.94) | 10.6 (4.7–12.7) | 41 (10–84) | 10.63 (4.57–19.20) | −2.42 (−8.99–5.26) |
| SAA | 19 (25.3) | 11 (5–42) | 7 (4–17) | 11 | 0.68 (0.27–1.94) | 8.2 (5.3–10.4) | 16 (5–50) | 11.11 (6.80–20.96) | −1.95 (−4.65–7.02) |
| VSAA | 5 (6.7) | 12 (6–17) | 10 (1–17) | 3 | 0.15 (0.05–0.26) | 8.2 (6.2–9.5) | 11 (7–16) | 8.50 (5.78–13.96) | −5.06 (−7.32–0.87) |
| Treatment | |||||||||
| Supportive | 50 (66.7) | 12 (5–31) | 9 (3–16) | 29 | 1.03 (0.08–1.94) | 10.5 (4.7–12.7) | 40 (10–84) | 10.86 (4.57–20.96) | −2.65 (−8.99–7.02) |
| IST | |||||||||
| Responder | 6 (8.0) | 25 (12–35) | 7 (1–15) | 2 | 0.90 (0.15–1.18) | 9.1 (7.7–12.3) | 36 (11–48) | 10.65 (5.78–12.81) | −1.21 (−7.62–0.57) |
| Non-responder | 5 (6.7) | 23 (10–42) | 12 (5–17) | 2 | 1.10 (0.31–1.94) | 8.6 (5.3–11.4) | 32 (7–80) | 9.54 (6.80–11.11) | −3.01 (−4.27–−2.42) |
| HSCT | 14 (18.7) | 12 (6–17) | 9 (4–17) | 10 | 0.44 (0.05–1.50) | 8.2 (6.2–12.1) | 15 (5–50) | 12.19 (7.09–20.08) | −1.02 (−6.78–6.29) |
| Genetics | |||||||||
| Variants (+) | 13 (27.1) | 11 (6–26) | 8 (3–16) | 10 | 1.30 (0.26–1.62) | 9.9 (4.7–12.4) | 44 (7–62) | 8.70 (4.57–13.17) | −4.83 (−8.99–0.00) |
| Variants (–) | 35 (72.9) | 11 (5–31) | 9 (4–17) | 21 | 0.83 (0.05–1.87) | 9.3 (6.2–12.7) | 32 (5–84) | 12.59 (6.72–20.96) | −0.58 (−5.95–7.02) |
| Gene | Nucleotide Change | Amino Acid Change | OMIM | Transcript | Functional Impact | ID |
|---|---|---|---|---|---|---|
| BRCA2 | c.1399A>T | p.(Lys467*) | Fanconi anemia, complementation group D1, (AR); Breast-ovarian cancer, familial, 2, (AD) | NM_000059.3 | Pathogenic | 8 |
| CDAN1 | c.3124C>T | p.(Arg1042Trp) | Dyserythropoietic anemia, congenital, type Ia (AR) | NM_138477.2 | Pathogenic | 57 |
| SLC37A5 | c.1042_1043del | p.(Leu348ValfsTer53) | Congenital disorder of glycosylation, type IIw, (AD) | NM_001164277.1 | Pathogenic | 73 |
| MPL | c.1025del | p.(Pro342GlnfsTer27) | Thrombocythemia 2 | NM_005373.2 | Pathogenic | 74 |
| PRF1 | c.65delC | p.(Pro22ArgfsTer29) | Aplastic anemia; Hemophagocytic lymphohistiocytosis, familial, 2, (AR); Lymphoma, non-Hodgkin | NM_001083116.1 | Likely Pathogenic | 18 |
| WRAP55 | c.914C>T | p.(Thr305Met) | Dyskeratosis congenita, autosomal recessive 3 (AR) | NM_018081.2 | VUS | 2 |
| JAGN1 | c.504G>T | p.(Lys168ASN) | Neutropenia, severe congenital, 6, autosomal recessive, 616022, (AR) | NM_032492.3 | VUS | 17 |
| BRIP1 | c.2258A>G | p.(Asp753Gly) | Fanconi anemia, complementation group J; {Breast cancer, early-onset, susceptibility to}, (AD, SMu) | NM_032043.2 | VUS | 18 |
| NBN | c.191C>G | p.(Pro64Arg) | Aplastic anemia; Leukemia, acute lymphoblastic; Nijmegen breakage syndrome, (AR) | NM_002485.4 | VUS | 24 |
| TERT | c.2795G>A | p.(gly932ASP) | Dyskeratosis congenita, autosomal dominant 2, (AD); Dyskeratosis congenita, autosomal recessive 4, (AR); Pulmonary fibrosis and/or bone marrow failure syndrome, telomere-related, 1, (AD) | NM_198253.3 | VUS | 48 |
| BRCA2 | c.4568G>T | p.(Gly1523Val) | Fanconi anemia, complementation group D1, (AR) | NM_000059.3 | VUS | 50 |
| WAS | c.1130G>A | p.(Arg377His) | Neutropenia, severe congenital; Thrombocytopenia; Wiskott–Aldrich syndrome | NM_000377.3 | VUS | 51 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, Y.; Lee, J.-M.; Lee, C.; Na, D.; Jung, J.; Ahn, A.; Yoo, J.W.; Lee, J.W.; Chung, N.-G.; Kim, M.; et al. Telomere Length and Genetic Variations in Acquired Pediatric Aplastic Anemia: A Flow-FISH Study in Korean Patients. Diagnostics 2025, 15, 931. https://doi.org/10.3390/diagnostics15070931
Hong Y, Lee J-M, Lee C, Na D, Jung J, Ahn A, Yoo JW, Lee JW, Chung N-G, Kim M, et al. Telomere Length and Genetic Variations in Acquired Pediatric Aplastic Anemia: A Flow-FISH Study in Korean Patients. Diagnostics. 2025; 15(7):931. https://doi.org/10.3390/diagnostics15070931
Chicago/Turabian StyleHong, Yuna, Jong-Mi Lee, Chaeyeon Lee, Duyeon Na, Jin Jung, Ari Ahn, Jae Won Yoo, Jae Wook Lee, Nack-Gyun Chung, Myungshin Kim, and et al. 2025. "Telomere Length and Genetic Variations in Acquired Pediatric Aplastic Anemia: A Flow-FISH Study in Korean Patients" Diagnostics 15, no. 7: 931. https://doi.org/10.3390/diagnostics15070931
APA StyleHong, Y., Lee, J.-M., Lee, C., Na, D., Jung, J., Ahn, A., Yoo, J. W., Lee, J. W., Chung, N.-G., Kim, M., & Kim, Y. (2025). Telomere Length and Genetic Variations in Acquired Pediatric Aplastic Anemia: A Flow-FISH Study in Korean Patients. Diagnostics, 15(7), 931. https://doi.org/10.3390/diagnostics15070931