
Medical Relevance, State-of-the-Art and Perspectives of “Sweet Metacode” in Liquid Biopsy Approaches

 , and
, and
Abstract
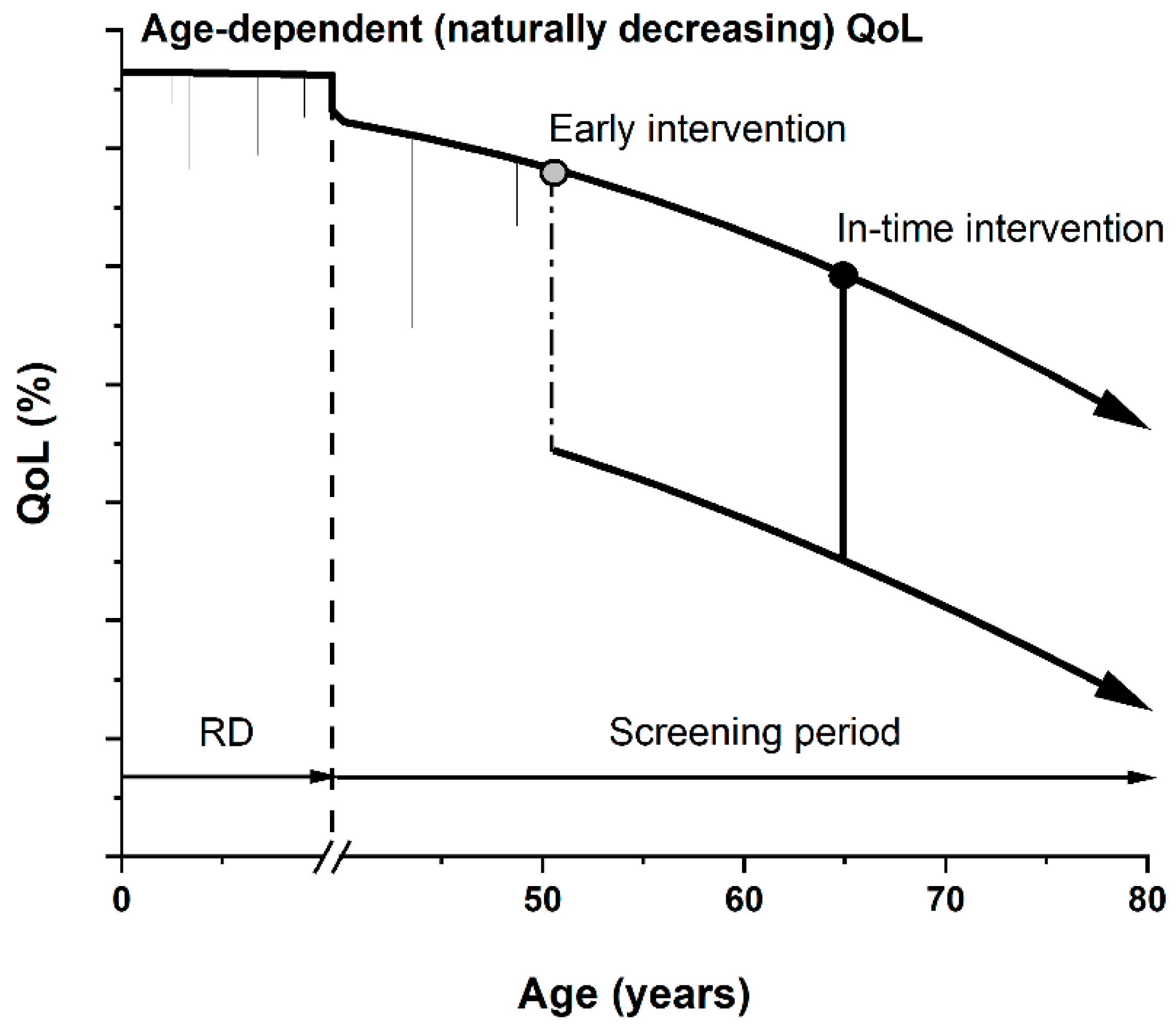
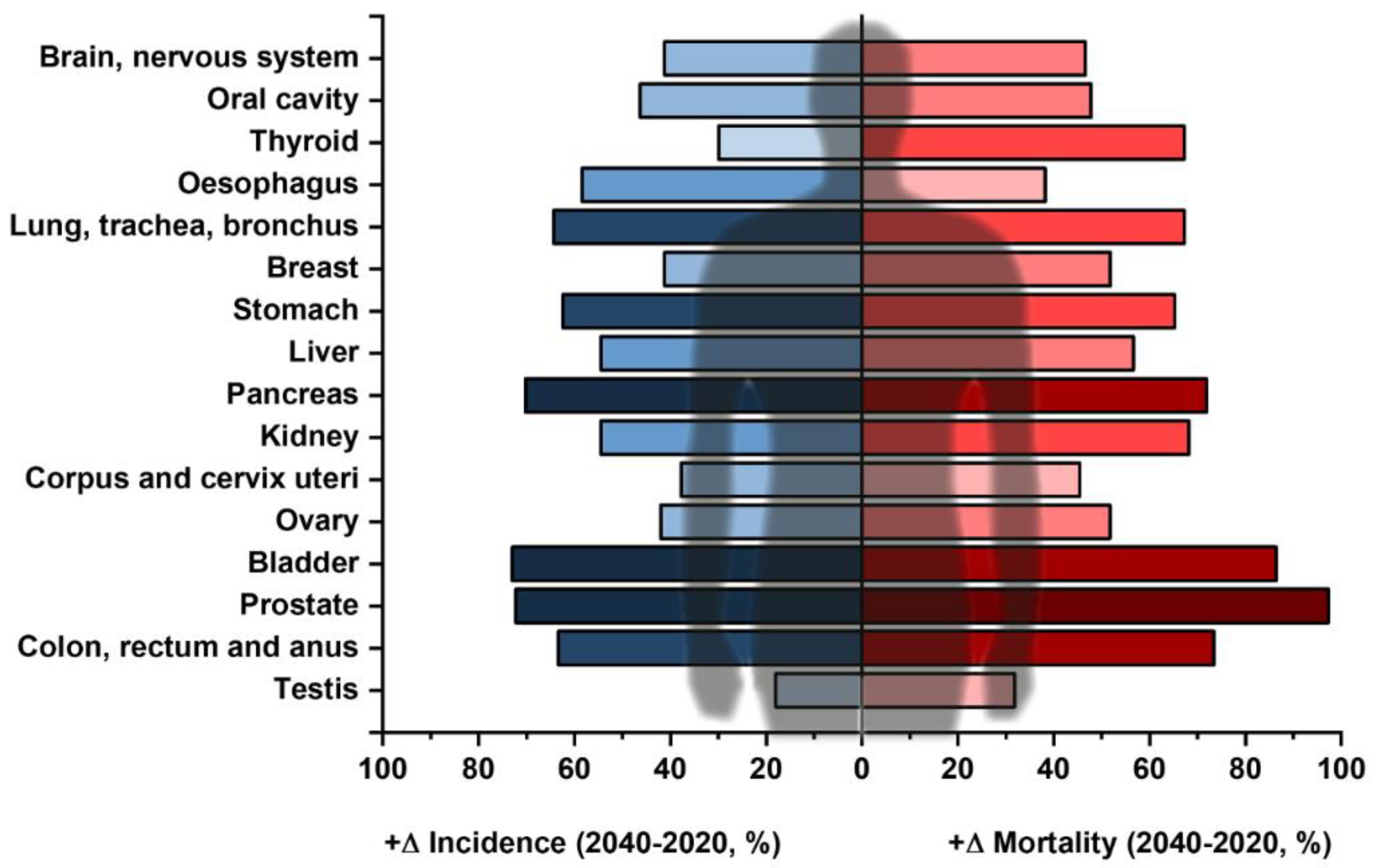
1. Current Challenges in Oncodiagnostics
Future Decades and Human Health
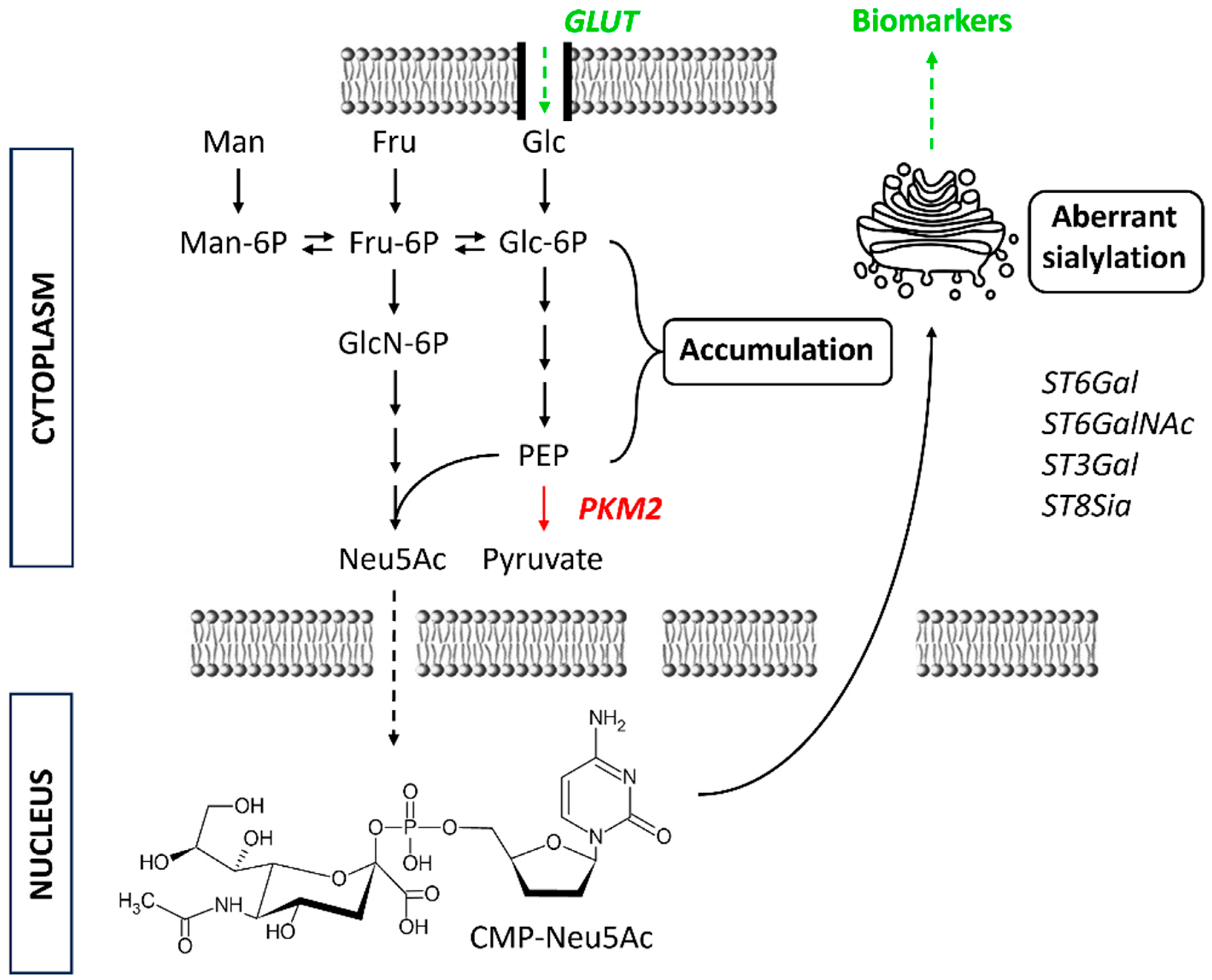
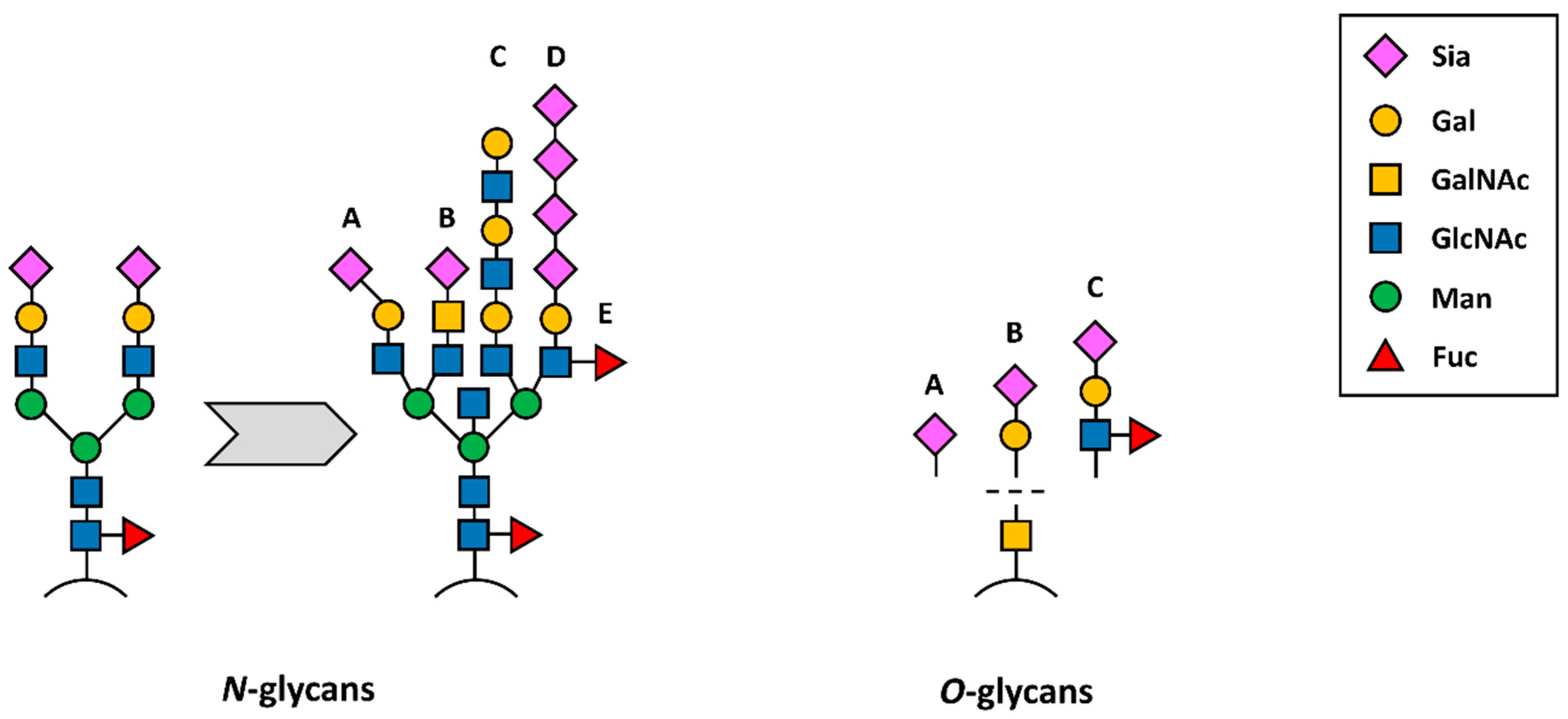
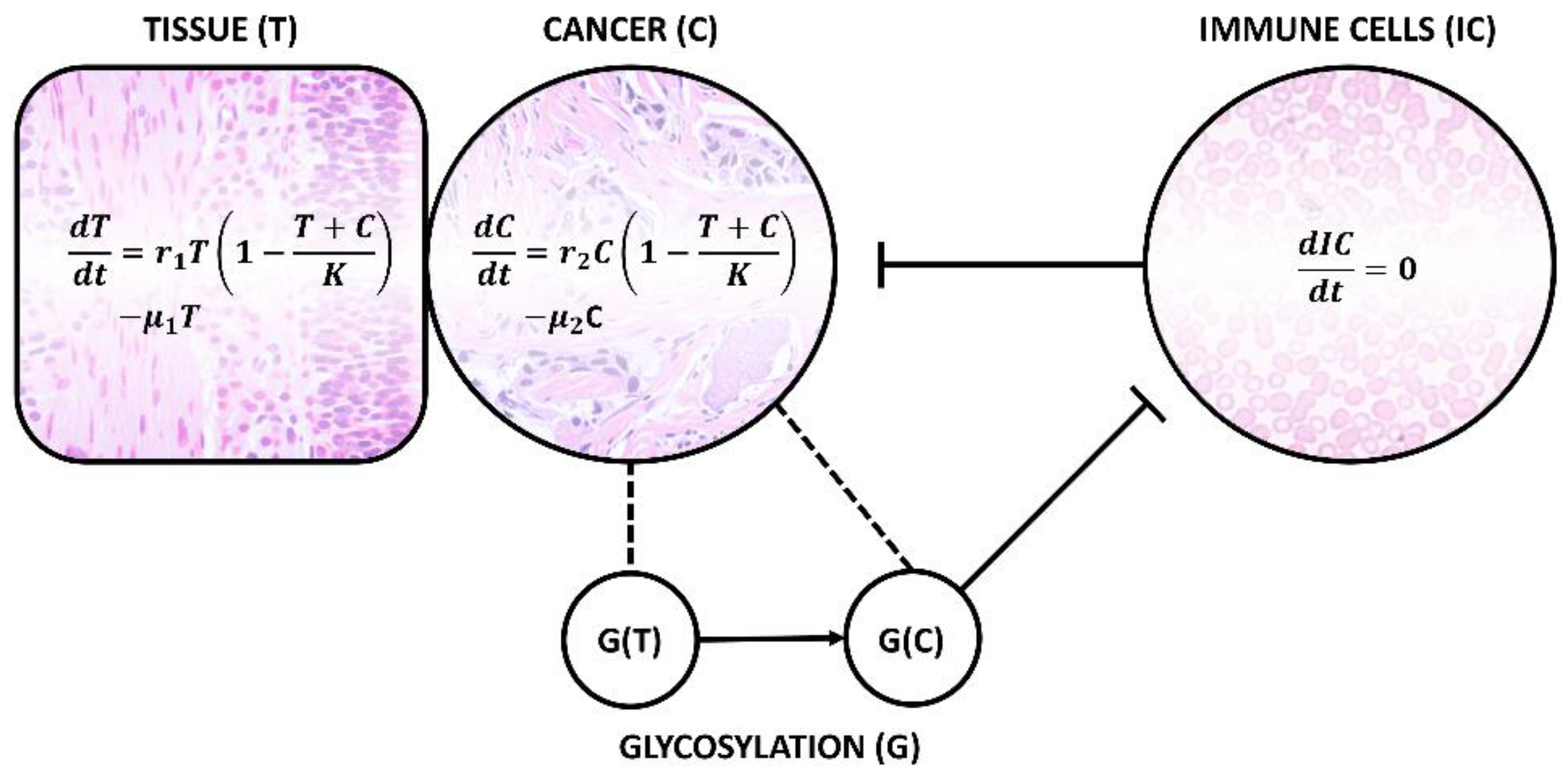
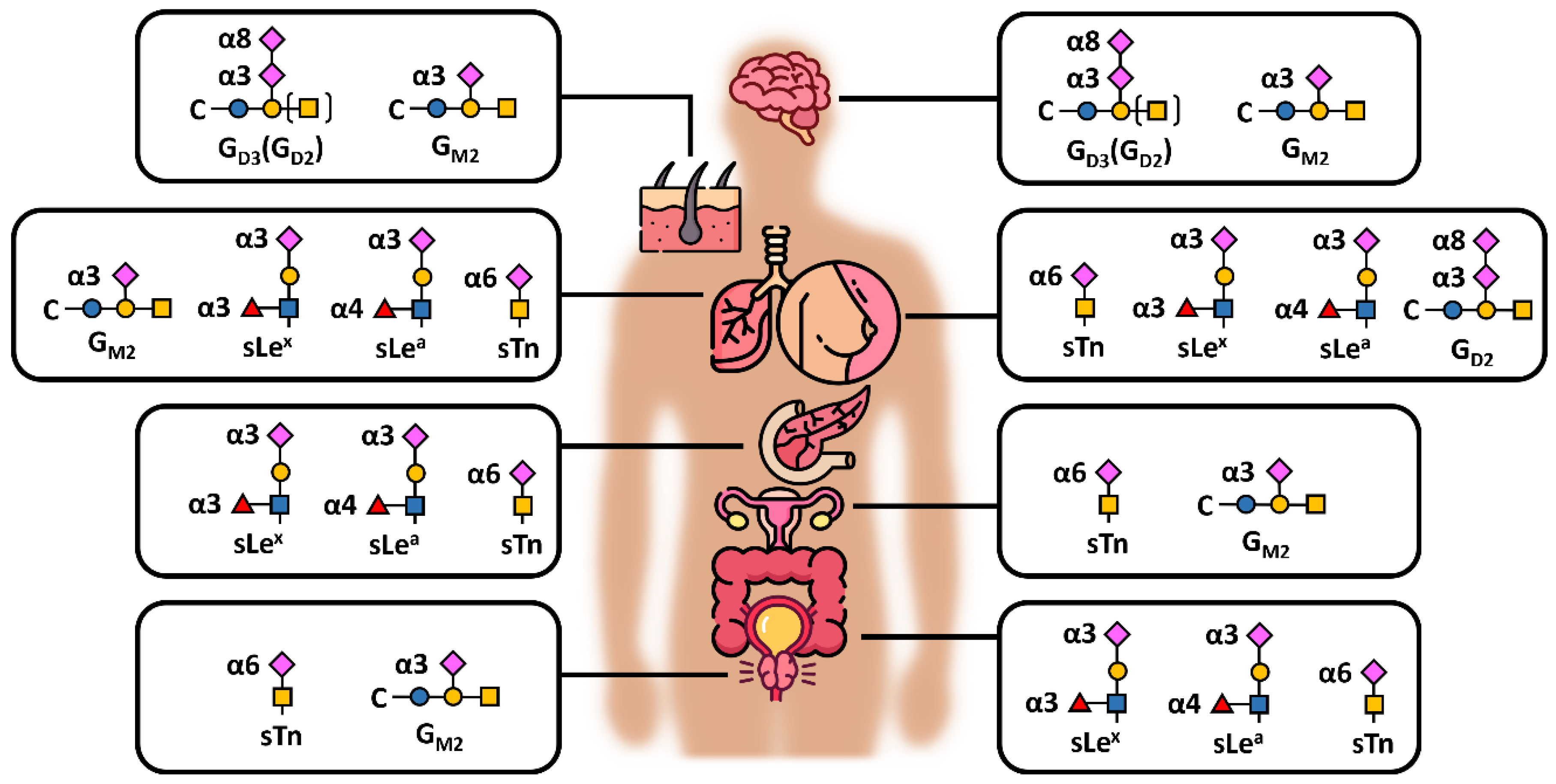
2. “Unpredictable” Metacode of Life—Biochemistry of Sialoglycoconjugates
Three Levels of Regulation—Genes, Substrates and Golgi

3. Techniques for Mapping the Human Oncosialome

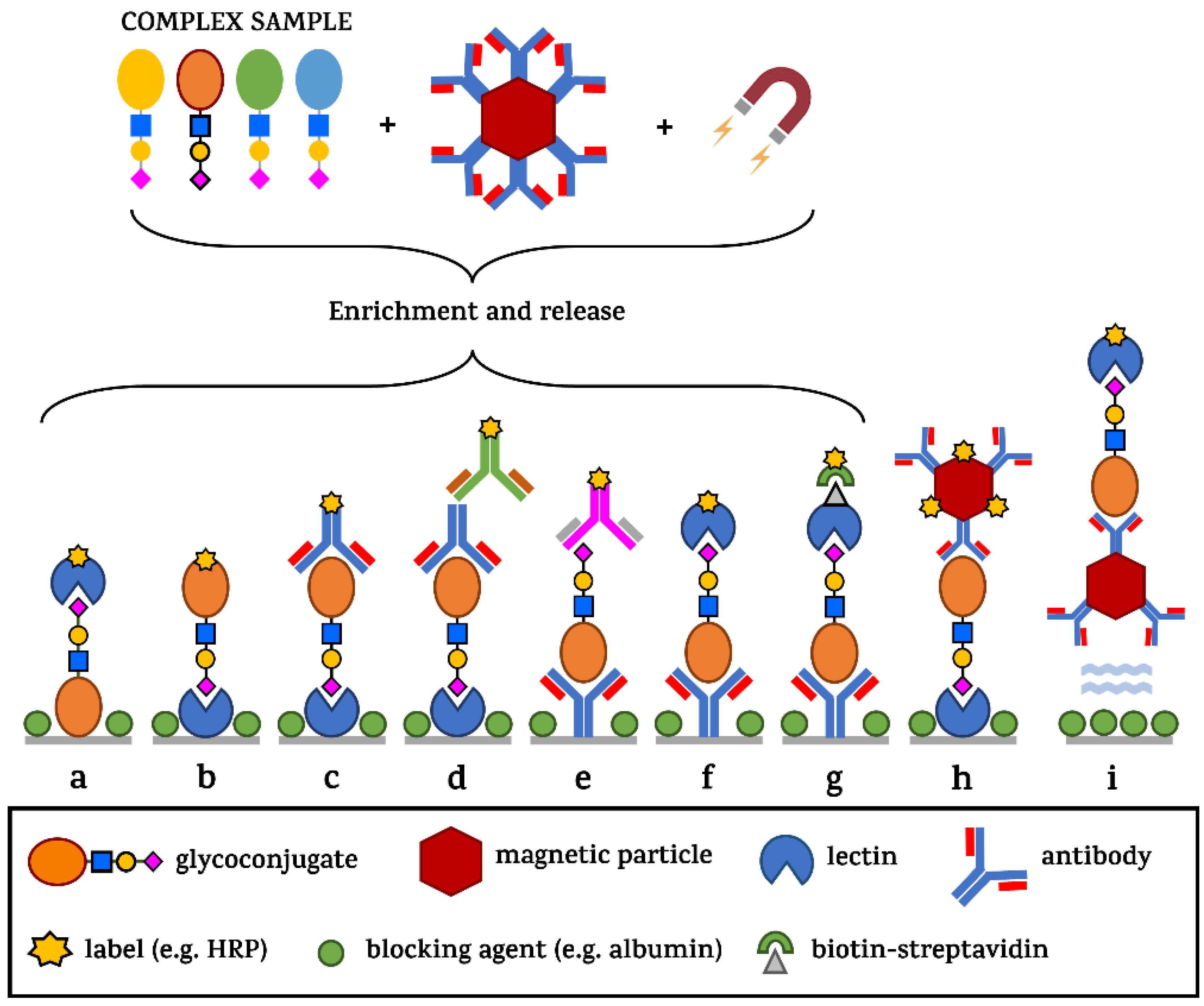
3.1. Complexity of Glycans as a Hope and Barrier in Early Diagnostics

3.2. Alternatives to Mass Spectrometry in Glycan Analysis
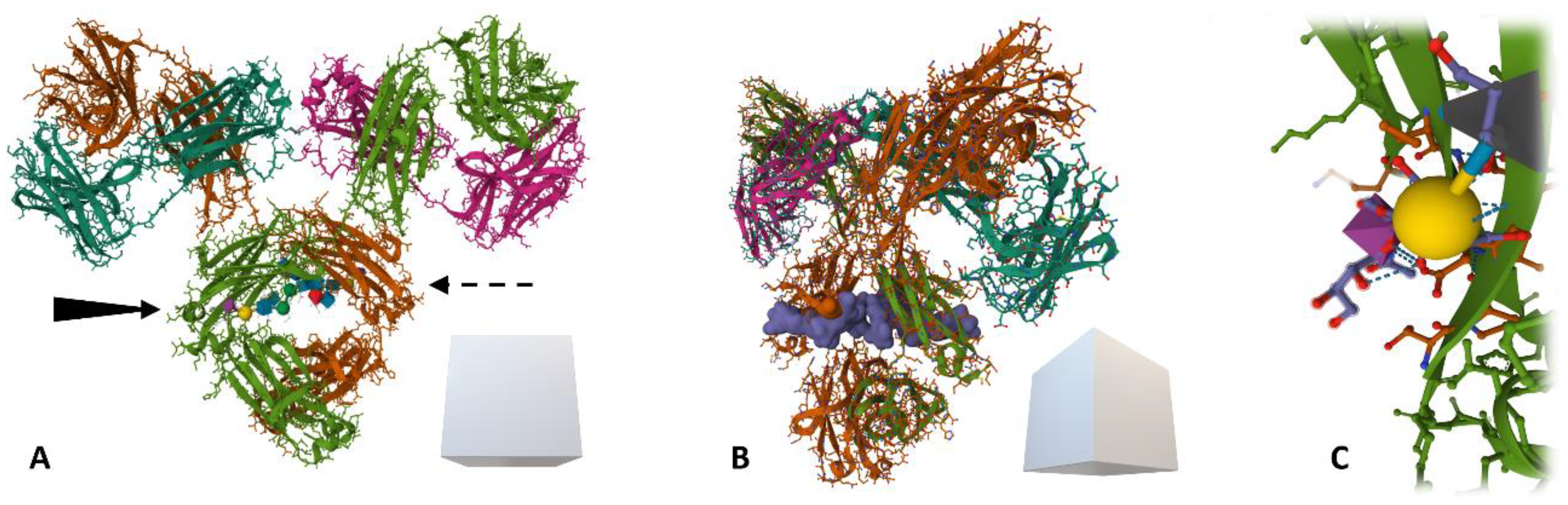
3.3. Glycosylation of Immunoglobulins in Diagnostics and Therapy


4. Perspectives of Glycan Liquid Biopsies and Clinical Validations
Performance of Glycans in Clinical Practice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumour Location | miRNAs (Combinations) | AUC | Ref. | Sia-Containing Glycans | AUC | Ref. |
|---|---|---|---|---|---|---|
| Bladder | miR-106a-5p, miR-145-5p, miR-132-3p, miR-7-5p, miR-148b-3p | 0.922 | [153] | Protein-bound Sia | 0.825 | [154] |
| Breast | miR15a, miR16 | 0.884 | [155] | Different Sia-glycan isoforms | Up to 0.980 | [156] |
| Colorectum | miR-1246, miR1268b, miR4648 | 0.821 | [157] | H5N4F1, H4N4F1, H5N4F1S2,61 | 0.830 | [158] |
| Lung | miR-210, miR-1290, miR-150, miR-21-5p | 0.930 | [159] | Different glycan isoforms + CRP | 0.942 * | [160] |
| Skin (melanoma) | miR-149-3p, miR-150-5p, miR-193a-3p | 0.970 | [161] | Total serum Neu5Gc | 0.925 | [162] |
| Ovaries | miR-92a, miR-200c, miR-320b, miR-320c, miR-335, miR-375, miR-486 | 0.870 | [163] | Total sialylation ratio (α-2,3-Sia) + CA125 | 0.985 * | [164] |
| Pancreas | miR-215-5p, miR-122-5p, miR-192-5p, miR-30b-5p, miR-320b | 0.811 | [165] | Combination of CA4, A3F0L and CFa glycan-isoforms | 0.807 | [166] |
| Prostate | miR-4286, miR-27a-3p, miR-29b-3p | 0.892 * (+PSA and PV) | [167] | α-2,3-Sia/PSA + PHI | 0.985 * | [168] |
| Stomach | 12-miRs panel | 0.920 | [169] | H5N5F1E2 glycan + other markers | 0.892 * | [170] |
| Testis | miR-371a-3p | 0.966 | [171] | 5 N-glycan score | 0.870 | [172] |
- Mass spectrometry (a robust gold standard in structural glycobiology) workflows need to be further transformed to advance the translation to clinics for suitable quantification of glycoconjugates. Mass spectrometry imaging for the observation of spatial distribution of glycans in histological samples is also of eminent importance.
- From a regulatory point of view, since there is currently a lack of any glycan detection diagnostic kits on the market based on common immunosorbent assays, any new products will be reviewed and judged according to common practices with ELISA. There are some aspects, however, which are unique for Enzyme-Linked Lecin Binding Assay (ELLBA) platforms, such as suppressing non-specificity based on lectin substrate promiscuity and low affinity between the lectins and glycans [173].
- In the therapeutic area, the first anti-core 1 O-glycans monoclonal antibody NEO-201 has been involved and registered in a phase I clinical trial in 2018, showing promising results in the treatment of solid tumours in 2023 [174]. Investments and support from large pharma companies in next rounds of clinical trials could help to accelerate advancement in the area and thus help to bring new molecules into clinical routine much sooner.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AFP | α-fetoprotein |
| AUC | area under the curve |
| BCa | breast cancer |
| CA | carbohydrate antigen (mucin) |
| CD | cluster of differentiation |
| CEA | carcinoembryonic antigen |
| ELLBA | Enzyme-Linked Lectin Binding Assay |
| FDA | Food and Drug Administration |
| GD, GM | tumour associated glycan antigens |
| HRP | horseradish peroxidase |
| IgG | Immunoglobulin G |
| KRAS | Kirsten rat sarcoma virus |
| (s)Le | (sialyl)-Lewis antigen |
| mTOR | mammalian target of rapamycin |
| NANA | N-acetylneuraminic acid (human form of sialic acid) |
| Neu5Ac | N-acetylneuraminic acid (human form of sialic acid) |
| NK | natural killer cells |
| PCa | prostate cancer |
| PHI | prostate health index by Beckman Coulter |
| PKM | pyruvate kinase muscle isoenzyme |
| PSA | prostate-specific antigen |
| PV | prostate volume |
| QoL | quality of life |
| ROC | receiver operating characteristic curve |
| ROS | reactive oxygen species |
| RT-qPCR | real-time quantitative polymerase chain reaction |
References
- The Top 10 Causes of Death. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 1 February 2024).
- About Medical Professionals Laboratory Week. Available online: https://www.cdc.gov/lab-week/about-archive.html#:~:text=Laboratory%20Heroes%20Save%20Lives&text=These%20laboratory%20heroes%20conduct%20approximately (accessed on 1 February 2024).
- Tkac, J.; Bertok, T. How glycomic studies can impact on prostate cancer. Expert Rev. Proteom. 2023, 20, 189–191. [Google Scholar] [CrossRef]
- Xie, W.; Suryaprakash, S.; Wu, C.; Rodriguez, A.; Fraterman, S. Trends in the use of liquid biopsy in oncology. Nat. Rev. Drug Discov. 2023, 22, 612–613. [Google Scholar] [CrossRef] [PubMed]
- Vrablova, V.; Kosutova, N.; Blsakova, A.; Bertokova, A.; Kasak, P.; Bertok, T.; Tkac, J. Glycosylation in extracellular vesicles: Isolation, characterization, composition, analysis and clinical applications. Biotechnol. Adv. 2023, 67, 108196. [Google Scholar] [CrossRef]
- Tkac, J.; Gajdosova, V.; Hroncekova, S.; Bertok, T.; Hires, M.; Jane, E.; Lorencova, L.; Kasak, P. Prostate-specific antigen glycoprofiling as diagnostic and prognostic biomarker of prostate cancer. Interface Focus. 2019, 9, 20180077. [Google Scholar] [CrossRef]
- Tkac, J.; Bertok, T.; Hires, M.; Jane, E.; Lorencova, L.; Kasak, P. Glycomics of prostate cancer: Updates. Expert. Rev. Proteom. 2019, 16, 65–76. [Google Scholar] [CrossRef]
- Dale, W.; Bilir, P.; Han, M.; Meltzer, D. The role of anxiety in prostate carcinoma: A structured review of the literature. Cancer 2005, 104, 467–478. [Google Scholar] [CrossRef]
- Kesch, C.; Heidegger, I.; Kasivisvanathan, V.; Kretschmer, A.; Marra, G.; Preisser, F.; Tilki, D.; Tsaur, I.; Valerio, M.; van den Bergh, R.C.N.; et al. Radical Prostatectomy: Sequelae in the Course of Time. Front. Surg. 2021, 8, 684088. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, A.N. Cancer diagnosis in primary care: Six steps to reducing the diagnostic interval. Can. Fam. Physician 2021, 67, 265–268. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Xu, J.-W.; Cheng, Y.-G.; Gao, J.-Y.; Hu, S.-Y.; Wang, L.; Zhan, H.-X. Early detection of pancreatic cancer: Where are we now and where are we going? Int. J. Cancer 2017, 141, 231–241. [Google Scholar] [CrossRef]
- Rao, A.; Vaught, J.; Tulskie, B.; Olson, D.; Odeh, H.; McLean, J.; Moore, H.M. Critical Financial Challenges for Biobanking: Report of a National Cancer Institute Study. Biopreserv. Biobank. 2019, 17, 129–138. [Google Scholar] [CrossRef]
- Austin, C.P. Opportunities and challenges in translational science. Clin. Transl. Sci. 2021, 14, 1629–1647. [Google Scholar] [CrossRef] [PubMed]
- Seyhan, A.A. Lost in translation: The valley of death across preclinical and clinical divide—Identification of problems and overcoming obstacles. Transl. Med. Commun. 2019, 4, 18. [Google Scholar] [CrossRef]
- Belkahla, S.; Nahvi, I.; Biswas, S.; Nahvi, I.; Ben Amor, N. Advances and development of prostate cancer, treatment, and strategies: A systemic review. Front. Cell Dev. Biol. 2022, 10, 991330. [Google Scholar] [CrossRef]
- Martinez-Useros, J.; Martin-Galan, M.; Florez-Cespedes, M.; Garcia-Foncillas, J. Epigenetics of Most Aggressive Solid Tumors: Pathways, Targets and Treatments. Cancers 2021, 13, 3209. [Google Scholar] [CrossRef]
- Alahdal, M.; Perera, R.A.; Moschovas, M.C.; Patel, V.; Perera, R.J. Current advances of liquid biopsies in prostate cancer: Molecular biomarkers. Mol. Ther.-Oncolytics 2023, 30, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Souza, V.G.P.; Forder, A.; Brockley, L.J.; Pewarchuk, M.E.; Telkar, N.; de Araújo, R.P.; Trejo, J.; Benard, K.; Seneda, A.L.; Minutentag, I.W.; et al. Liquid Biopsy in Lung Cancer: Biomarkers for the Management of Recurrence and Metastasis. Int. J. Mol. Sci. 2023, 24, 8894. [Google Scholar] [CrossRef]
- Connal, S.; Cameron, J.M.; Sala, A.; Brennan, P.M.; Palmer, D.S.; Palmer, J.D.; Perlow, H.; Baker, M.J. Liquid biopsies: The future of cancer early detection. J. Transl. Med. 2023, 21, 118. [Google Scholar] [CrossRef] [PubMed]
- Mariotto, A.B.; Enewold, L.; Zhao, J.; Zeruto, C.A.; Yabroff, K.R. Medical Care Costs Associated with Cancer Survivorship in the United States. Cancer Epidemiol. Biomark. Prev. 2020, 29, 1304–1312. [Google Scholar] [CrossRef]
- Christou, N.; Veyrune, L.; Popeskou, S.G.; Mathonnet, M. Personalized Therapy and Liquid Biopsy-A Focus on Colorectal Cancer. J. Pers. Med. 2021, 11, 630. [Google Scholar] [CrossRef]
- Lindpaintner, K.; Srinivasan, A.; Mitchell, A.; Dixit, A.; Xu, G.; Cong, X.; Serie, D. 158 A novel, highly accurate liquid biopsy-based glycoproteomic predictor of checkpoint inhibitor treatment benefit in advanced non-small cell lung cancer. J. ImmunoTherapy Cancer 2022, 10 (Suppl. S2), A171. [Google Scholar]
- Mitchell, A.; Pickering, C.; Xu, G.; Rice, R.; Castellanos, A.; Bhadra, R.; Brcic, L.; Lindenmann, J.; Smolle, F.; Lindpaintner, K.; et al. Glycoproteomics as a powerful liquid biopsy-based screening tool for non-small cell lung cancer. J. Clin. Oncol. 2022, 40 (Suppl. S16), e21148. [Google Scholar] [CrossRef]
- Cancer Today. Available online: https://gco.iarc.fr/ (accessed on 1 February 2024).
- How Liquid Biopsies Have Made It Easier to Treat Cancer. Available online: https://www.nationalgeographic.com/magazine/article/how-liquid-biopsies-have-made-it-easier-to-treat-cancer (accessed on 1 February 2024).
- Cohen, J.D.; Li, L.; Wang, Y.; Thoburn, C.; Afsari, B.; Danilova, L.; Douville, C.; Javed, A.A.; Wong, F.; Mattox, A.; et al. Detection and localization of surgically resectable cancers with a multi-analyte blood test. Science 2018, 359, 926–930. [Google Scholar] [CrossRef] [PubMed]
- Bertok, T.; Jane, E.; Bertokova, A.; Lorencova, L.; Zvara, P.; Smolkova, B.; Kucera, R.; Klocker, H.; Tkac, J. Validating fPSA Glycoprofile as a Prostate Cancer Biomarker to Avoid Unnecessary Biopsies and Re-Biopsies. Cancers 2020, 12, 2988. [Google Scholar] [CrossRef] [PubMed]
- Lévêque, L.; Outtas, M.; Liu, H.; Zhang, L. Comparative study of the methodologies used for subjective medical image quality assessment. Phys. Med. Biol. 2021, 66, 15TR02. [Google Scholar] [CrossRef]
- Reily, C.; Stewart, T.J.; Renfrow, M.B.; Novak, J. Glycosylation in health and disease. Nat. Rev. Nephrol. 2019, 15, 346–366. [Google Scholar] [CrossRef]
- Ohtsubo, K.; Marth, J.D. Glycosylation in cellular mechanisms of health and disease. Cell 2006, 126, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Pinho, S.S.; Reis, C.A. Glycosylation in cancer: Mechanisms and clinical implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef] [PubMed]
- Rini, J.M.; Moremen, K.W.; Davis, B.G.; Esko, J.D. Glycosyltransferases and glycan-processing enzymes. In Essentials of Glycobiology, 4th ed.; Cold Spring Harbor Laboratory Press: Woodbury, NY, USA, 2022. [Google Scholar]
- Thomas, D.; Rathinavel, A.K.; Radhakrishnan, P. Altered glycosylation in cancer: A promising target for biomarkers and therapeutics. Biochim. Biophys. Acta Rev. Cancer 2021, 1875, 188464. [Google Scholar] [CrossRef]
- Zhang, X. Alterations of Golgi Structural Proteins and Glycosylation Defects in Cancer. Front. Cell Dev. Biol. 2021, 9, 665289. [Google Scholar] [CrossRef]
- Mikkola, S. Nucleotide Sugars in Chemistry and Biology. Molecules 2020, 25, 5755. [Google Scholar] [CrossRef]
- Warburg, O.; Wind, F.; Negelein, E. The metabolism of tumors in the body. J. Gen. Physiol. 1927, 8, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does it Benefit Cancer Cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, Q.; Huang, X.; Yang, M.; Zhou, S.; Li, Z.; Fang, Z.; Tang, Y.; Chen, Q.; Hou, H.; et al. Lactate in the tumor microenvironment: A rising star for targeted tumor therapy. Front. Nutr. 2023, 10, 1113739. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Tomás, R.; Pérez-Guillén, I. Lactate in the Tumor Microenvironment: An Essential Molecule in Cancer Progression and Treatment. Cancers 2020, 12, 3244. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, A.; Molt, M.; Uribe, E.; Salas, M. Glut 1 in Cancer Cells and the Inhibitory Action of Resveratrol as A Potential Therapeutic Strategy. Int. J. Mol. Sci. 2019, 20, 3374. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Jiang, P. The crisscross between p53 and metabolism in cancer. Acta Biochim. Biophys. Sin. 2023, 55, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, M.; Riscal, R.; Arena, G.; Linares, L.K.; Le Cam, L. Metabolic functions of the tumor suppressor p53: Implications in normal physiology, metabolic disorders, and cancer. Mol. Metab. 2020, 33, 2–22. [Google Scholar] [CrossRef]
- Shin, E.; Koo, J.S. Glucose Metabolism and Glucose Transporters in Breast Cancer. Front. Cell Dev. Biol. 2021, 9, 728759. [Google Scholar] [CrossRef]
- Munemoto, M.; Mukaisho, K.I.; Miyashita, T.; Oyama, K.; Haba, Y.; Okamoto, K.; Kinoshita, J.; Ninomiya, I.; Fushida, S.; Taniura, N.; et al. Roles of the hexosamine biosynthetic pathway and pentose phosphate pathway in bile acid-induced cancer development. Cancer Sci. 2019, 110, 2408–2420. [Google Scholar] [CrossRef]
- Akella, N.M.; Ciraku, L.; Reginato, M.J. Fueling the fire: Emerging role of the hexosamine biosynthetic pathway in cancer. BMC Biol. 2019, 17, 52. [Google Scholar] [CrossRef]
- Chen, X.; Chen, S.; Yu, D. Protein kinase function of pyruvate kinase M2 and cancer. Cancer Cell Int. 2020, 20, 523. [Google Scholar] [CrossRef]
- Zahra, K.; Dey, T.; Ashish; Mishra, S.P.; Pandey, U. Pyruvate Kinase M2 and Cancer: The Role of PKM2 in Promoting Tumorigenesis. Front. Oncol. 2020, 10, 159. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, J.; Sunthar, P.; Balaji, P.V. The glycan alphabet is not universal: A hypothesis. Microb. Genom. 2020, 6, mgen000452. [Google Scholar] [CrossRef] [PubMed]
- Varki, A. Uniquely human evolution of sialic acid genetics and biology. Proc. Natl. Acad. Sci. USA 2010, 107 (Suppl. S2), 8939–8946. [Google Scholar] [CrossRef]
- Jahan, M.; Thomson, P.C.; Wynn, P.C.; Wang, B. The non-human glycan, N-glycolylneuraminic acid (Neu5Gc), is not expressed in all organs and skeletal muscles of nine animal species. Food Chem. 2021, 343, 128439. [Google Scholar] [CrossRef]
- Varki, A. Sialic acids in human health and disease. Trends Mol. Med. 2008, 14, 351–360. [Google Scholar] [CrossRef]
- Samraj, A.N.; Pearce, O.M.T.; Läubli, H.; Crittenden, A.N.; Bergfeld, A.K.; Banda, K.; Gregg, C.J.; Bingman, A.E.; Secrest, P.; Diaz, S.L.; et al. A red meat-derived glycan promotes inflammation and cancer progression. Proc. Natl. Acad. Sci. USA 2015, 112, 542–547. [Google Scholar] [CrossRef]
- Tang, W.; Liu, D.; Nie, S.-P. Food glycomics in food science: Recent advances and future perspectives. Curr. Opin. Food Sci. 2022, 46, 100850. [Google Scholar] [CrossRef]
- Mansoori, M.; Mirzaei, A.; Abdi Rad, I.; Mahmodlou, R.; Mansouri, F.; Saeednejad Zanjani, L.; Asadi-Lari, Z.; Madjd, Z. Upregulation of Ganglioside GD2 Synthase (GD2S), as a New Putative Cancer Stem Cell Marker in Breast Carcinomas. Med. J. Islam. Repub. Iran. 2021, 35, 148. [Google Scholar] [CrossRef]
- Shewell, L.K.; Day, C.J.; Kutasovic, J.R.; Abrahams, J.L.; Wang, J.; Poole, J.; Niland, C.; Ferguson, K.; Saunus, J.M.; Lakhani, S.R.; et al. N-glycolylneuraminic acid serum biomarker levels are elevated in breast cancer patients at all stages of disease. BMC Cancer 2022, 22, 334. [Google Scholar] [CrossRef]
- Yang, W.; Jiang, Y.; Guo, Q.; Tian, Z.; Cheng, Z. Aberrant N-glycolylneuraminic acid in breast MCF-7 cancer cells and cancer stem cells. Front. Mol. Biosci. 2022, 9, 1047672. [Google Scholar] [CrossRef]
- Shewell, L.K.; Wang, J.J.; Paton, J.C.; Paton, A.W.; Day, C.J.; Jennings, M.P. Detection of N-glycolylneuraminic acid biomarkers in sera from patients with ovarian cancer using an engineered N-glycolylneuraminic acid-specific lectin SubB2M. Biochem. Biophys. Res. Commun. 2018, 507, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Yehuda, S.; Padler-Karavani, V. Glycosylated Biotherapeutics: Immunological Effects of N-Glycolylneuraminic Acid. Front. Immunol. 2020, 11, 21. [Google Scholar] [CrossRef]
- Dammen-Brower, K.; Epler, P.; Zhu, S.; Bernstein, Z.J.; Stabach, P.R.; Braddock, D.T.; Spangler, J.B.; Yarema, K.J. Strategies for Glycoengineering Therapeutic Proteins. Front. Chem. 2022, 10, 863118. [Google Scholar] [CrossRef]
- Mahajan, V.S.; Pillai, S. Sialic acids and autoimmune disease. Immunol. Rev. 2016, 269, 145–161. [Google Scholar] [CrossRef]
- Li, Y.; Chen, X. Sialic acid metabolism and sialyltransferases: Natural functions and applications. Appl. Microbiol. Biotechnol. 2012, 94, 887–905. [Google Scholar] [CrossRef]
- Hugonnet, M.; Singh, P.; Haas, Q.; von Gunten, S. The Distinct Roles of Sialyltransferases in Cancer Biology and Onco-Immunology. Front. Immunol. 2021, 12, 799861. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.A.H.; Deutzmann, A.; Correa, K.M.; Delaveris, C.S.; Dhanasekaran, R.; Dove, C.G.; Sullivan, D.K.; Wisnovsky, S.; Stark, J.C.; Pluvinage, J.V.; et al. MYC-driven synthesis of Siglec ligands is a glycoimmune checkpoint. Proc. Natl. Acad. Sci. USA 2023, 120, e2215376120. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Chi, K.; Zhang, C.; Liu, Q.; Yang, G. Sialylation: A Cloak for Tumors to Trick the Immune System in the Microenvironment. Biology 2023, 12, 832. [Google Scholar] [CrossRef]
- Altevogt, P.; Sammar, M.; Hüser, L.; Kristiansen, G. Novel insights into the function of CD24: A driving force in cancer. Int. J. Cancer 2021, 148, 546–559. [Google Scholar] [CrossRef]
- Li, F.; Lv, B.; Liu, Y.; Hua, T.; Han, J.; Sun, C.; Xu, L.; Zhang, Z.; Feng, Z.; Cai, Y.; et al. Blocking the CD47-SIRPα axis by delivery of anti-CD47 antibody induces antitumor effects in glioma and glioma stem cells. Oncoimmunology 2018, 7, e1391973. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Molina, N.; Parsons, J.; Decker, E.L.; Reski, R. Structural modelling of human complement FHR1 and two of its synthetic derivatives provides insight into their in-vivo functions. Comput. Struct. Biotechnol. J. 2023, 21, 1473–1486. [Google Scholar] [CrossRef] [PubMed]
- Rosenstock, P.; Kaufmann, T. Sialic Acids and Their Influence on Human NK Cell Function. Cells 2021, 10, 263. [Google Scholar] [CrossRef] [PubMed]
- Loureiro, L.R.; Sousa, D.P.; Ferreira, D.; Chai, W.; Lima, L.; Pereira, C.; Lopes, C.B.; Correia, V.G.; Silva, L.M.; Li, C.; et al. Novel monoclonal antibody L2A5 specifically targeting sialyl-Tn and short glycans terminated by alpha-2–6 sialic acids. Sci. Rep. 2018, 8, 12196. [Google Scholar] [CrossRef] [PubMed]
- Miyata, S.; Sato, C.; Kumita, H.; Toriyama, M.; Vacquier, V.D.; Kitajima, K. Flagellasialin: A novel sulfated alpha2,9-linked polysialic acid glycoprotein of sea urchin sperm flagella. Glycobiology 2006, 16, 1229–1241. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Martínez, I.E.; Ramos-Martínez, E.; Segura-Velázquez, R.; Saavedra-Montañez, M.; Cervantes-Torres, J.B.; Cerbón, M.; Papy-Garcia, D.; Zenteno, E.; Sánchez-Betancourt, J.I. Heparan Sulfate and Sialic Acid in Viral Attachment: Two Sides of the Same Coin? Int. J. Mol. Sci. 2022, 23, 9842. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.L. The role of cell surface sialic acids for SARS-CoV-2 infection. Glycobiology 2021, 31, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Wu, X.; Lei, T.; Zhong, R.; Wang, Y.; Zhang, L.; Zhao, Q.; Huang, Y.; Shi, Y.; Wu, L. The Role and Therapeutic Value of Syndecan-1 in Cancer Metastasis and Drug Resistance. Front. Cell Dev. Biol. 2022, 9, 784983. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Spetz, M.R.; Ho, M. The Role of Glypicans in Cancer Progression and Therapy. J. Histochem. Cytochem. 2020, 68, 841–862. [Google Scholar] [CrossRef]
- Habes, C.; Weber, G.; Goupille, C. Sulfated Glycoaminoglycans and Proteoglycan Syndecan-4 Are Involved in Membrane Fixation of LL-37 and Its Pro-Migratory Effect in Breast Cancer Cells. Biomolecules 2019, 9, 481. [Google Scholar] [CrossRef]
- Nardy, A.F.; Freire-de-Lima, L.; Freire-de-Lima, C.G.; Morrot, A. The Sweet Side of Immune Evasion: Role of Glycans in the Mechanisms of Cancer Progression. Front. Oncol. 2016, 6, 54. [Google Scholar] [CrossRef]
- Yin, A.; Moes, D.; van Hasselt, J.G.C.; Swen, J.J.; Guchelaar, H.J. A Review of Mathematical Models for Tumor Dynamics and Treatment Resistance Evolution of Solid Tumors. CPT Pharmacomet. Syst. Pharmacol. 2019, 8, 720–737. [Google Scholar] [CrossRef]
- Yang, Y.; Cao, Y. The impact of VEGF on cancer metastasis and systemic disease. Semin. Cancer Biol. 2022, 86, 251–261. [Google Scholar] [CrossRef]
- Cheng, W.K.; Oon, C.E. How glycosylation aids tumor angiogenesis: An updated review. Biomed. Pharmacother. 2018, 103, 1246–1252. [Google Scholar] [CrossRef]
- Peixoto, A.; Relvas-Santos, M.; Azevedo, R.; Santos, L.L.; Ferreira, J.A. Protein Glycosylation and Tumor Microenvironment Alterations Driving Cancer Hallmarks. Front. Oncol. 2019, 9, 380. [Google Scholar] [CrossRef]
- Anggara, K.; Sršan, L.; Jaroentomeechai, T.; Wu, X.; Rauschenbach, S.; Narimatsu, Y.; Clausen, H.; Ziegler, T.; Miller, R.L.; Kern, K. Direct observation of glycans bonded to proteins and lipids at the single-molecule level. Science 2023, 382, 219–223. [Google Scholar] [CrossRef]
- Tubiana, M. Tumor Cell Proliferation Kinetics and Tumor Growth Rate. Acta Oncol. 1989, 28, 113–121. [Google Scholar] [CrossRef]
- Revannasiddaiah, S.; Maka, V.V.; Devadas, S.K. Tumour growth kinetics as a potential predictive and prognostic factor in carcinoma of the lung. Ann. Transl. Med. 2019, 7, 295. [Google Scholar] [CrossRef]
- Freeman, H.J. Early stage colon cancer. World J. Gastroenterol. 2013, 19, 8468–8473. [Google Scholar] [CrossRef]
- Brierley, J.; Gospodarowicz, M.; O’Sullivan, B. The principles of cancer staging. Ecancermedicalscience 2016, 10, ed61. [Google Scholar] [CrossRef]
- Lee, T.; Teng, T.Z.J.; Shelat, V.G. Carbohydrate antigen 19-9—Tumor marker: Past, present, and future. World J. Gastrointest. Surg. 2020, 12, 468–490. [Google Scholar] [CrossRef]
- Yuan, Q.; Song, J.; Yang, W.; Wang, H.; Huo, Q.; Yang, J.; Yu, X.; Liu, Y.; Xu, C.; Bao, H. The effect of CA125 on metastasis of ovarian cancer: Old marker new function. Oncotarget 2017, 8, 50015–50022. [Google Scholar] [CrossRef]
- Jin, S.; Cheng, Y.; Reid, S.; Li, M.; Wang, B. Carbohydrate recognition by boronolectins, small molecules, and lectins. Med. Res. Rev. 2010, 30, 171–257. [Google Scholar] [CrossRef]
- Haab, B.B.; Klamer, Z. Advances in Tools to Determine the Glycan-Binding Specificities of Lectins and Antibodies. Mol. Cell. Proteom. 2020, 19, 224–232. [Google Scholar] [CrossRef]
- Arnaud, J.; Audfray, A.; Imberty, A. Binding sugars: From natural lectins to synthetic receptors and engineered neolectins. Chem. Soc. Rev. 2013, 42, 4798–4813. [Google Scholar] [CrossRef]
- Xiao, H.; Chen, W.; Smeekens, J.M.; Wu, R. An enrichment method based on synergistic and reversible covalent interactions for large-scale analysis of glycoproteins. Nat. Commun. 2018, 9, 1692. [Google Scholar] [CrossRef]
- Hashim, O.H.; Jayapalan, J.J.; Lee, C.S. Lectins: An effective tool for screening of potential cancer biomarkers. PeerJ 2017, 5, e3784. [Google Scholar] [CrossRef]
- Bertok, T.; Jane, E.; Chrenekova, N.; Hroncekova, S.; Bertokova, A.; Hires, M.; Vikartovska, A.; Kubanikova, P.; Sokol, R.; Fillo, J.; et al. Analysis of serum glycome by lectin microarrays for prostate cancer patients—A search for aberrant glycoforms. Glycoconj. J. 2020, 37, 703–711. [Google Scholar] [CrossRef]
- Bertok, T.; Bertokova, A.; Jane, E.; Hires, M.; Aguedo, J.; Potocarova, M.; Lukac, L.; Vikartovska, A.; Kasak, P.; Borsig, L.; et al. Identification of Whole-Serum Glycobiomarkers for Colorectal Carcinoma Using Reverse-Phase Lectin Microarray. Front. Oncol. 2021, 11, 735338. [Google Scholar] [CrossRef]
- Zhou, S.; Wooding, K.M.; Mechref, Y. Analysis of Permethylated Glycan by Liquid Chromatography (LC) and Mass Spectrometry (MS). Methods Mol. Biol. 2017, 1503, 83–96. [Google Scholar]
- Aguedo, J.; Pakanova, Z.; Lorencova, L.; Nemcovic, M.; Kasak, P.; Barath, M.; Farkas, P.; Tkac, J. MXene as a novel cartridge for N-glycan enrichment. Anal. Chim. Acta 2022, 1234, 340512. [Google Scholar] [CrossRef]
- Han, L.; Costello, C.E. Mass spectrometry of glycans. Biochemistry 2013, 78, 710–720. [Google Scholar] [CrossRef]
- Zacharias, L.G.; Hartmann, A.K.; Song, E.; Zhao, J.; Zhu, R.; Mirzaei, P.; Mechref, Y. HILIC and ERLIC Enrichment of Glycopeptides Derived from Breast and Brain Cancer Cells. J. Proteome Res. 2016, 15, 3624–3634. [Google Scholar] [CrossRef]
- Dosekova, E.; Filip, J.; Bertok, T.; Both, P.; Kasak, P.; Tkac, J. Nanotechnology in Glycomics: Applications in Diagnostics, Therapy, Imaging, and Separation Processes. Med. Res. Rev. 2017, 37, 514–626. [Google Scholar] [CrossRef]
- de Haan, N.; Reiding, K.R.; Haberger, M.; Reusch, D.; Falck, D.; Wuhrer, M. Linkage-specific sialic acid derivatization for MALDI-TOF-MS profiling of IgG glycopeptides. Anal. Chem. 2015, 87, 8284–8291. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, Z.; Wang, Y.; Zheng, Q.; Li, J. Mass spectrometry for protein sialoglycosylation. Mass. Spectrom. Rev. 2018, 37, 652–680. [Google Scholar] [CrossRef]
- Nishikaze, T. Sialic acid derivatization for glycan analysis by mass spectrometry. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2019, 95, 523–537. [Google Scholar] [CrossRef]
- Kadirvelraj, R.; Boruah, B.M.; Wang, S.; Chapla, D.; Huang, C.; Ramiah, A.; Hudson, K.L.; Prudden, A.R.; Boons, G.-J.; Withers, S.G.; et al. Structural basis for Lewis antigen synthesis by the α1,3-fucosyltransferase FUT9. Nat. Chem. Biol. 2023, 19, 1022–1030. [Google Scholar] [CrossRef]
- Cavdarli, S.; Dewald, J.H.; Yamakawa, N.; Guérardel, Y.; Terme, M.; Le Doussal, J.M.; Delannoy, P.; Groux-Degroote, S. Identification of 9-O-acetyl-N-acetylneuraminic acid (Neu5,9Ac(2)) as main O-acetylated sialic acid species of GD2 in breast cancer cells. Glycoconj. J. 2019, 36, 79–90. [Google Scholar] [CrossRef]
- Munkley, J. The Role of Sialyl-Tn in Cancer. Int. J. Mol. Sci. 2016, 17, 275. [Google Scholar] [CrossRef]
- Dube, D.H.; Bertozzi, C.R. Glycans in cancer and inflammation—Potential for therapeutics and diagnostics. Nat. Rev. Drug Discov. 2005, 4, 477–488. [Google Scholar] [CrossRef]
- Dang, K.; Zhang, W.; Jiang, S.; Lin, X.; Qian, A. Application of Lectin Microarrays for Biomarker Discovery. ChemistryOpen 2020, 9, 285–300. [Google Scholar] [CrossRef]
- Hires, M.; Jane, E.; Kalavska, K.; Chovanec, M.; Mego, M.; Kasak, P.; Bertok, T.; Tkac, J. Glycan signatures for the identification of cisplatin-resistant testicular cancer cell lines: Specific glycoprofiling of human chorionic gonadotropin (hCG). Cancer Med. 2022, 11, 968–982. [Google Scholar] [CrossRef]
- Silva, M.L.S. Lectin biosensors in cancer glycan biomarker detection. Adv. Clin. Chem. 2019, 93, 1–61. [Google Scholar]
- Silva, M.L.S. Lectin-based biosensors as analytical tools for clinical oncology. Cancer Lett. 2018, 436, 63–74. [Google Scholar] [CrossRef]
- Haab, B.B. Using lectins in biomarker research: Addressing the limitations of sensitivity and availability. Proteom. Clin. Appl. 2012, 6, 346–350. [Google Scholar] [CrossRef]
- Geisler, C.; Jarvis, D.L. Effective glycoanalysis with Maackia amurensis lectins requires a clear understanding of their binding specificities. Glycobiology 2011, 21, 988–993. [Google Scholar] [CrossRef]
- Ryva, B.; Zhang, K.; Asthana, A.; Wong, D.; Vicioso, Y.; Parameswaran, R. Wheat Germ Agglutinin as a Potential Therapeutic Agent for Leukemia. Front. Oncol. 2019, 9, 100. [Google Scholar] [CrossRef]
- Parasuraman, P.; Murugan, V.; Selvin, J.F.; Gromiha, M.M.; Fukui, K.; Veluraja, K. Insights into the binding specificity of wild type and mutated wheat germ agglutinin towards Neu5Acα(2-3)Gal: A study by in silico mutations and molecular dynamics simulations. J. Mol. Recognit. 2014, 27, 482–492. [Google Scholar] [CrossRef]
- Paleček, E.; Tkáč, J.; Bartošík, M.; Bertók, T.s.; Ostatná, V.; Paleček, J. Electrochemistry of Nonconjugated Proteins and Glycoproteins. Toward Sensors for Biomedicine and Glycomics. Chem. Rev. 2015, 115, 2045–2108. [Google Scholar] [CrossRef]
- Yaghoubi, M.; Rahimi, F.; Negahdari, B.; Rezayan, A.H.; Shafiekhani, A. A lectin-coupled porous silicon-based biosensor: Label-free optical detection of bacteria in a real-time mode. Sci. Rep. 2020, 10, 16017. [Google Scholar] [CrossRef]
- Xiao, Y.; Isaacs, S.N. Enzyme-linked immunosorbent assay (ELISA) and blocking with bovine serum albumin (BSA)—Not all BSAs are alike. J. Immunol. Methods 2012, 384, 148–151. [Google Scholar] [CrossRef]
- Chen, S.; LaRoche, T.; Hamelinck, D.; Bergsma, D.; Brenner, D.; Simeone, D.; Brand, R.E.; Haab, B.B. Multiplexed analysis of glycan variation on native proteins captured by antibody microarrays. Nat. Methods 2007, 4, 437–444. [Google Scholar] [CrossRef]
- Su, Z.; Xie, Q.; Wang, Y.; Li, Y. Abberant Immunoglobulin G Glycosylation in Rheumatoid Arthritis by LTQ-ESI-MS. Int. J. Mol. Sci. 2020, 21, 2045. [Google Scholar] [CrossRef]
- Malhotra, R.; Wormald, M.R.; Rudd, P.M.; Fischer, P.B.; Dwek, R.A.; Sim, R.B. Glycosylation changes of IgG associated with rheumatoid arthritis can activate complement via the mannose-binding protein. Nat. Med. 1995, 1, 237–243. [Google Scholar] [CrossRef]
- Parekh, R.B.; Dwek, R.A.; Sutton, B.J.; Fernandes, D.L.; Leung, A.; Stanworth, D.; Rademacher, T.W.; Mizuochi, T.; Taniguchi, T.; Matsuta, K.; et al. Association of rheumatoid arthritis and primary osteoarthritis with changes in the glycosylation pattern of total serum IgG. Nature 1985, 316, 452–457. [Google Scholar] [CrossRef]
- Boune, S.; Hu, P.; Epstein, A.L.; Khawli, L.A. Principles of N-Linked Glycosylation Variations of IgG-Based Therapeutics: Pharmacokinetic and Functional Considerations. Antibodies 2020, 9, 22. [Google Scholar] [CrossRef]
- Elias, S.; Kahlon, S.; Kotzur, R.; Kaynan, N.; Mandelboim, O. Obinutuzumab activates FcγRI more potently than other anti-CD20 antibodies in chronic lymphocytic leukemia (CLL). Oncoimmunology 2018, 7, e1428158. [Google Scholar] [CrossRef]
- Pereira, N.A.; Chan, K.F.; Lin, P.C.; Song, Z. The “less-is-more” in therapeutic antibodies: Afucosylated anti-cancer antibodies with enhanced antibody-dependent cellular cytotoxicity. MAbs 2018, 10, 693–711. [Google Scholar] [CrossRef]
- Gabius, H.J.; Cudic, M.; Diercks, T.; Kaltner, H.; Kopitz, J.; Mayo, K.H.; Murphy, P.V.; Oscarson, S.; Roy, R.; Schedlbauer, A.; et al. What is the Sugar Code? Chembiochem 2021, 23, e202100327. [Google Scholar] [CrossRef]
- Gabius, H.-J. The sugar code: Why glycans are so important. Biosystems 2018, 164, 102–111. [Google Scholar] [CrossRef]
- Villalobo, A.; Gabius, H. Signaling pathways for transduction of the initial message of the glycocode into cellular responses. Acta Anat. 1998, 161, 110–129. [Google Scholar] [CrossRef]
- Gillmann, K.M.; Temme, J.S.; Marglous, S.; Brown, C.E.; Gildersleeve, J.C. Anti-glycan monoclonal antibodies: Basic research and clinical applications. Curr. Opin. Chem. Biol. 2023, 74, 102281. [Google Scholar] [CrossRef]
- Váradi, C.; Hajdu, V.; Farkas, F.; Gilányi, I.; Oláh, C.; Viskolcz, B. The Analysis of Human Serum N-Glycosylation in Patients with Primary and Metastatic Brain Tumors. Life 2021, 11, 29. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, S.; Liu, J.; Zhang, Y.; Liu, Y.; Shen, S.; Tian, F.; Yan, G.; Gao, Y.; Qin, X. Identification of serum glycobiomarkers for Hepatocellular Carcinoma using lectin microarrays. Front. Immunol. 2022, 13, 973993. [Google Scholar] [CrossRef]
- Haakensen, V.D.; Steinfeld, I.; Saldova, R.; Shehni, A.A.; Kifer, I.; Naume, B.; Rudd, P.M.; Børresen-Dale, A.-L.; Yakhini, Z. Serum N-glycan analysis in breast cancer patients—Relation to tumour biology and clinical outcome. Mol. Oncol. 2016, 10, 59–72. [Google Scholar] [CrossRef]
- Doherty, M.; Theodoratou, E.; Walsh, I.; Adamczyk, B.; Stöckmann, H.; Agakov, F.; Timofeeva, M.; Trbojević-Akmačić, I.; Vučković, F.; Duffy, F.; et al. Plasma N-glycans in colorectal cancer risk. Sci. Rep. 2018, 8, 8655. [Google Scholar] [CrossRef]
- Liang, Y.; Han, P.; Wang, T.; Ren, H.; Gao, L.; Shi, P.; Zhang, S.; Yang, A.; Li, Z.; Chen, M. Stage-associated differences in the serum N- and O-glycan profiles of patients with non-small cell lung cancer. Clin. Proteom. 2019, 16, 20. [Google Scholar] [CrossRef]
- Gebrehiwot, A.G.; Melka, D.S.; Kassaye, Y.M.; Rehan, I.F.; Rangappa, S.; Hinou, H.; Kamiyama, T.; Nishimura, S.-I. Healthy human serum N-glycan profiling reveals the influence of ethnic variation on the identified cancer-relevant glycan biomarkers. PLoS ONE 2018, 13, e0209515. [Google Scholar] [CrossRef]
- Cindrić, A.; Krištić, J.; Martinić Kavur, M.; Pezer, M. Glycosylation and Aging. Adv. Exp. Med. Biol. 2021, 1325, 341–373. [Google Scholar]
- Chircov, C.; Grumezescu, A.M.; Holban, A.M. Magnetic Particles for Advanced Molecular Diagnosis. Materials 2019, 12, 2158. [Google Scholar] [CrossRef]
- Ha, Y. Exploiting the Potential of Magnetic Nanoparticles for Rapid Diagnosis Tests (RDTs): Nanoparticle-Antibody Conjugates and Color Development Strategies. Diagnostics 2023, 13, 3033. [Google Scholar] [CrossRef]
- Halawa, T.; Baeesa, S.; Fadul, M.M.; Badahdah, A.A.; Enani, M.; Fathaddin, A.A.; Kawass, D.; Alkhotani, A.; Bahakeem, B.; Kurdi, M. The Role of Liquid Biopsy in the Diagnosis and Prognosis of WHO Grade 4 Astrocytoma. Cureus 2023, 15, e41221. [Google Scholar] [CrossRef]
- Dave, V.P.; Ngo, T.A.; Pernestig, A.-K.; Tilevik, D.; Kant, K.; Nguyen, T.; Wolff, A.; Bang, D.D. MicroRNA amplification and detection technologies: Opportunities and challenges for point of care diagnostics. Lab. Investig. 2019, 99, 452–469. [Google Scholar] [CrossRef]
- Beaver, J.A.; Tzou, A.; Blumenthal, G.M.; McKee, A.E.; Kim, G.; Pazdur, R.; Philip, R. An FDA Perspective on the Regulatory Implications of Complex Signatures to Predict Response to Targeted Therapies. Clin. Cancer Res. 2017, 23, 1368–1372. [Google Scholar] [CrossRef]
- Zarinshenas, R.; Amini, A.; Mambetsariev, I.; Abuali, T.; Fricke, J.; Ladbury, C.; Salgia, R. Assessment of Barriers and Challenges to Screening, Diagnosis, and Biomarker Testing in Early-Stage Lung Cancer. Cancers 2023, 15, 1595. [Google Scholar] [CrossRef]
- Liu, X.; Papukashvili, D.; Wang, Z.; Liu, Y.; Chen, X.; Li, J.; Li, Z.; Hu, L.; Li, Z.; Rcheulishvili, N.; et al. Potential utility of miRNAs for liquid biopsy in breast cancer. Front. Oncol. 2022, 12, 940314. [Google Scholar] [CrossRef]
- Buhagiar, A.; Seria, E.; Borg, M.; Borg, J.; Ayers, D. Overview of microRNAs as liquid biopsy biomarkers for colorectal cancer sub-type profiling and chemoresistance. Cancer Drug Resist. 2021, 4, 934–945. [Google Scholar] [CrossRef]
- Preethi, K.A.; Selvakumar, S.C.; Ross, K.; Jayaraman, S.; Tusubira, D.; Sekar, D. Liquid biopsy: Exosomal microRNAs as novel diagnostic and prognostic biomarkers in cancer. Mol. Cancer 2022, 21, 54. [Google Scholar] [CrossRef]
- Di Bella, M.A. Overview and Update on Extracellular Vesicles: Considerations on Exosomes and Their Application in Modern Medicine. Biology 2022, 11, 804. [Google Scholar] [CrossRef]
- Bertokova, A.; Svecova, N.; Kozics, K.; Gabelova, A.; Vikartovska, A.; Jane, E.; Hires, M.; Bertok, T.; Tkac, J. Exosomes from prostate cancer cell lines: Isolation optimisation and characterisation. Biomed. Pharmacother. 2022, 151, 113093. [Google Scholar] [CrossRef]
- Verbakel, J.Y.; Steyerberg, E.W.; Uno, H.; De Cock, B.; Wynants, L.; Collins, G.S.; Van Calster, B. ROC curves for clinical prediction models part 1. ROC plots showed no added value above the AUC when evaluating the performance of clinical prediction models. J. Clin. Epidemiol. 2020, 126, 207–216. [Google Scholar] [CrossRef]
- Halligan, S.; Altman, D.G.; Mallett, S. Disadvantages of using the area under the receiver operating characteristic curve to assess imaging tests: A discussion and proposal for an alternative approach. Eur. Radiol. 2015, 25, 932–939. [Google Scholar] [CrossRef]
- Movahedi, F.; Padman, R.; Antaki, J.F. Limitations of receiver operating characteristic curve on imbalanced data: Assist device mortality risk scores. J. Thorac. Cardiovasc. Surg. 2023, 165, 1433–1442.e2. [Google Scholar] [CrossRef]
- Post, M.A.; Lefeber, D.J. Clinical glycomics in the diagnostic laboratory. Ann. Transl. Med. 2019, 7 (Suppl. S6), S220. [Google Scholar] [CrossRef]
- de Haan, N.; Pučić-Baković, M.; Novokmet, M.; Falck, D.; Lageveen-Kammeijer, G.; Razdorov, G.; Vučković, F.; Trbojević-Akmačić, I.; Gornik, O.; Hanić, M.; et al. Developments and perspectives in high-throughput protein glycomics: Enabling the analysis of thousands of samples. Glycobiology 2022, 32, 651–663. [Google Scholar] [CrossRef]
- Lauc, G.; Pezer, M.; Rudan, I.; Campbell, H. Mechanisms of disease: The human N-glycome. Biochim. Biophys. Acta 2016, 1860, 1574–1582. [Google Scholar] [CrossRef]
- Li, R.; Xia, Y.; Chen, X.; Li, X.; Huang, G.; Peng, X.; Liu, K.; Zhang, C.; Li, M.; Lin, Y.; et al. Identification of a three-miRNA panel in serum for bladder cancer diagnosis by a diagnostic test. Transl. Cancer Res. 2022, 11, 1005–1016. [Google Scholar] [CrossRef]
- Habibi, S.; Jamshidian, H.; Kadivar, M.; Eshraghian, M.R.; Javanbakht, M.H.; Derakhshanian, H.; Djalali, M. A study of lipid- and protein- bound sialic acids for the diagnosis of bladder cancer and their relationships with the severity of malignancy. Rep. Biochem. Mol. Biol. 2014, 2, 70–75. [Google Scholar]
- Borsos, B.N.; Páhi, Z.G.; Ujfaludi, Z.; Sükösd, F.; Nikolényi, A.; Bankó, S.; Pankotai-Bodó, G.; Oláh-Németh, O.; Pankotai, T. BC-miR: Monitoring Breast Cancer-Related miRNA Profile in Blood Sera—A Prosperous Approach for Tumor Detection. Cells 2022, 11, 2721. [Google Scholar] [CrossRef]
- Alley, W.R., Jr.; Novotny, M.V. Glycomic analysis of sialic acid linkages in glycans derived from blood serum glycoproteins. J. Proteome Res. 2010, 9, 3062–3072. [Google Scholar] [CrossRef]
- Yoshikawa, Y.; Fukunaga, M.; Takahashi, J.; Shimizu, D.; Masuda, T.; Mizushima, T.; Yamada, K.; Mori, M.; Eguchi, H.; Doki, Y.; et al. Identification of the Minimum Combination of Serum microRNAs to Predict the Recurrence of Colorectal Cancer Cases. Ann. Surg. Oncol. 2023, 30, 233–243. [Google Scholar] [CrossRef]
- Moran, A.B.; Elgood-Hunt, G.; van der Burgt, Y.E.M.; Wuhrer, M.; Mesker, W.E.; Tollenaar, R.; Spencer, D.I.R.; Lageveen-Kammeijer, G.S.M. Serum N-Glycosylation RPLC-FD-MS Assay to Assess Colorectal Cancer Surgical Interventions. Biomolecules 2023, 13, 896. [Google Scholar] [CrossRef]
- Jiang, H.G.; Dai, C.H.; Xu, Y.P.; Jiang, Q.; Xia, X.B.; Shu, Y.; Li, J. Four plasma miRNAs act as biomarkers for diagnosis and prognosis of non-small cell lung cancer. Oncol. Lett. 2021, 22, 792. [Google Scholar] [CrossRef]
- Arnold, J.N.; Saldova, R.; Galligan, M.C.; Murphy, T.B.; Mimura-Kimura, Y.; Telford, J.E.; Godwin, A.K.; Rudd, P.M. Novel Glycan Biomarkers for the Detection of Lung Cancer. J. Proteome Res. 2011, 10, 1755–1764. [Google Scholar] [CrossRef]
- Fogli, S.; Polini, B.; Carpi, S.; Pardini, B.; Naccarati, A.; Dubbini, N.; Lanza, M.; Breschi, M.C.; Romanini, A.; Nieri, P. Identification of plasma microRNAs as new potential biomarkers with high diagnostic power in human cutaneous melanoma. Tumour Biol. 2017, 39, 1010428317701646. [Google Scholar] [CrossRef]
- Shewell, L.K.; Day, C.J.; Hippolite, T.; De Bisscop, X.; Paton, J.C.; Paton, A.W.; Jennings, M.P. Serum Neu5Gc biomarkers are elevated in primary cutaneous melanoma. Biochem. Biophys. Res. Commun. 2023, 642, 162–166. [Google Scholar] [CrossRef]
- Gahlawat, A.W.; Witte, T.; Haarhuis, L.; Schott, S. A novel circulating miRNA panel for non-invasive ovarian cancer diagnosis and prognosis. Br. J. Cancer 2022, 127, 1550–1556. [Google Scholar] [CrossRef]
- Dědová, T.; Braicu, E.I.; Sehouli, J.; Blanchard, V. Sialic Acid Linkage Analysis Refines the Diagnosis of Ovarian Cancer. Front. Oncol. 2019, 9, 261. [Google Scholar] [CrossRef]
- Khan, I.A.; Rashid, S.; Singh, N.; Rashid, S.; Singh, V.; Gunjan, D.; Das, P.; Dash, N.R.; Pandey, R.M.; Chauhan, S.S.; et al. Panel of serum miRNAs as potential non-invasive biomarkers for pancreatic ductal adenocarcinoma. Sci. Rep. 2021, 11, 2824. [Google Scholar] [CrossRef]
- Vreeker, G.C.M.; Hanna-Sawires, R.G.; Mohammed, Y.; Bladergroen, M.R.; Nicolardi, S.; Dotz, V.; Nouta, J.; Bonsing, B.A.; Mesker, W.E.; van der Burgt, Y.E.M.; et al. Serum N-Glycome analysis reveals pancreatic cancer disease signatures. Cancer Med. 2020, 9, 8519–8529. [Google Scholar] [CrossRef]
- Lyu, J.; Zhao, L.; Wang, F.; Ji, J.; Cao, Z.; Xu, H.; Shi, X.; Zhu, Y.; Zhang, C.; Guo, F.; et al. Discovery and Validation of Serum MicroRNAs as Early Diagnostic Biomarkers for Prostate Cancer in Chinese Population. BioMed Res. Int. 2019, 2019, 9306803. [Google Scholar] [CrossRef]
- Ferrer-Batallé, M.; Llop, E.; Ramírez, M.; Aleixandre, R.N.; Saez, M.; Comet, J.; De Llorens, R.; Peracaula, R. Comparative Study of Blood-Based Biomarkers, α2,3-Sialic Acid PSA and PHI, for High-Risk Prostate Cancer Detection. Int. J. Mol. Sci. 2017, 18, 845. [Google Scholar] [CrossRef]
- So, J.B.Y.; Kapoor, R.; Zhu, F.; Koh, C.; Zhou, L.; Zou, R.; Tang, Y.C.; Goo, P.C.K.; Rha, S.Y.; Chung, H.C.; et al. Development and validation of a serum microRNA biomarker panel for detecting gastric cancer in a high-risk population. Gut 2021, 70, 829–837. [Google Scholar] [CrossRef]
- Zhao, J.; Qin, R.; Chen, H.; Yang, Y.; Qin, W.; Han, J.; Wang, X.; Ren, S.; Sun, Y.; Gu, J. A nomogram based on glycomic biomarkers in serum and clinicopathological characteristics for evaluating the risk of peritoneal metastasis in gastric cancer. Clin. Proteom. 2020, 17, 34. [Google Scholar] [CrossRef]
- Dieckmann, K.P.; Radtke, A.; Geczi, L.; Matthies, C.; Anheuser, P.; Eckardt, U.; Sommer, J.; Zengerling, F.; Trenti, E.; Pichler, R.; et al. Serum Levels of MicroRNA-371a-3p (M371 Test) as a New Biomarker of Testicular Germ Cell Tumors: Results of a Prospective Multicentric Study. J. Clin. Oncol. 2019, 37, 1412–1423. [Google Scholar] [CrossRef]
- Narita, T.; Hatakeyama, S.; Yoneyama, T.; Narita, S.; Yamashita, S.; Mitsuzuka, K.; Sakurai, T.; Kawamura, S.; Tochigi, T.; Takahashi, I.; et al. Clinical implications of serum N-glycan profiling as a diagnostic and prognostic biomarker in germ-cell tumors. Cancer Med. 2017, 6, 739–748. [Google Scholar] [CrossRef]
- Pinkeova, A.; Tomikova, A.; Bertokova, A.; Fabinyova, E.; Bartova, R.; Jane, E.; Hroncekova, S.; Sievert, K.-D.; Sokol, R.; Jirasko, M. Glycoprofiling of proteins as prostate cancer biomarkers: A multinational population study. PLoS ONE 2024, 19, e0300430. [Google Scholar] [CrossRef]
- Cole, C.B.; Morelli, M.P.; Fantini, M.; Miettinen, M.; Fetsch, P.; Peer, C.; Figg, W.D.; Yin, T.; Houston, N.; McCoy, A.; et al. First-in-human phase 1 clinical trial of anti-core 1 O-glycans targeting monoclonal antibody NEO-201 in treatment-refractory solid tumors. J. Exp. Clin. Cancer Res. 2023, 42, 76. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinkeova, A.; Kosutova, N.; Jane, E.; Lorencova, L.; Bertokova, A.; Bertok, T.; Tkac, J. Medical Relevance, State-of-the-Art and Perspectives of “Sweet Metacode” in Liquid Biopsy Approaches. Diagnostics 2024, 14, 713. https://doi.org/10.3390/diagnostics14070713
Pinkeova A, Kosutova N, Jane E, Lorencova L, Bertokova A, Bertok T, Tkac J. Medical Relevance, State-of-the-Art and Perspectives of “Sweet Metacode” in Liquid Biopsy Approaches. Diagnostics. 2024; 14(7):713. https://doi.org/10.3390/diagnostics14070713
Chicago/Turabian StylePinkeova, Andrea, Natalia Kosutova, Eduard Jane, Lenka Lorencova, Aniko Bertokova, Tomas Bertok, and Jan Tkac. 2024. "Medical Relevance, State-of-the-Art and Perspectives of “Sweet Metacode” in Liquid Biopsy Approaches" Diagnostics 14, no. 7: 713. https://doi.org/10.3390/diagnostics14070713
APA StylePinkeova, A., Kosutova, N., Jane, E., Lorencova, L., Bertokova, A., Bertok, T., & Tkac, J. (2024). Medical Relevance, State-of-the-Art and Perspectives of “Sweet Metacode” in Liquid Biopsy Approaches. Diagnostics, 14(7), 713. https://doi.org/10.3390/diagnostics14070713