
Magnetic Resonance Imaging and Spectroscopy Analysis in a Pelizaeus–Merzbacher Disease Rat Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials & Methods
2.1. Animals
2.2. MRI Measurements
2.3. LFB Staining
2.4. Statistical Analysis
3. Results
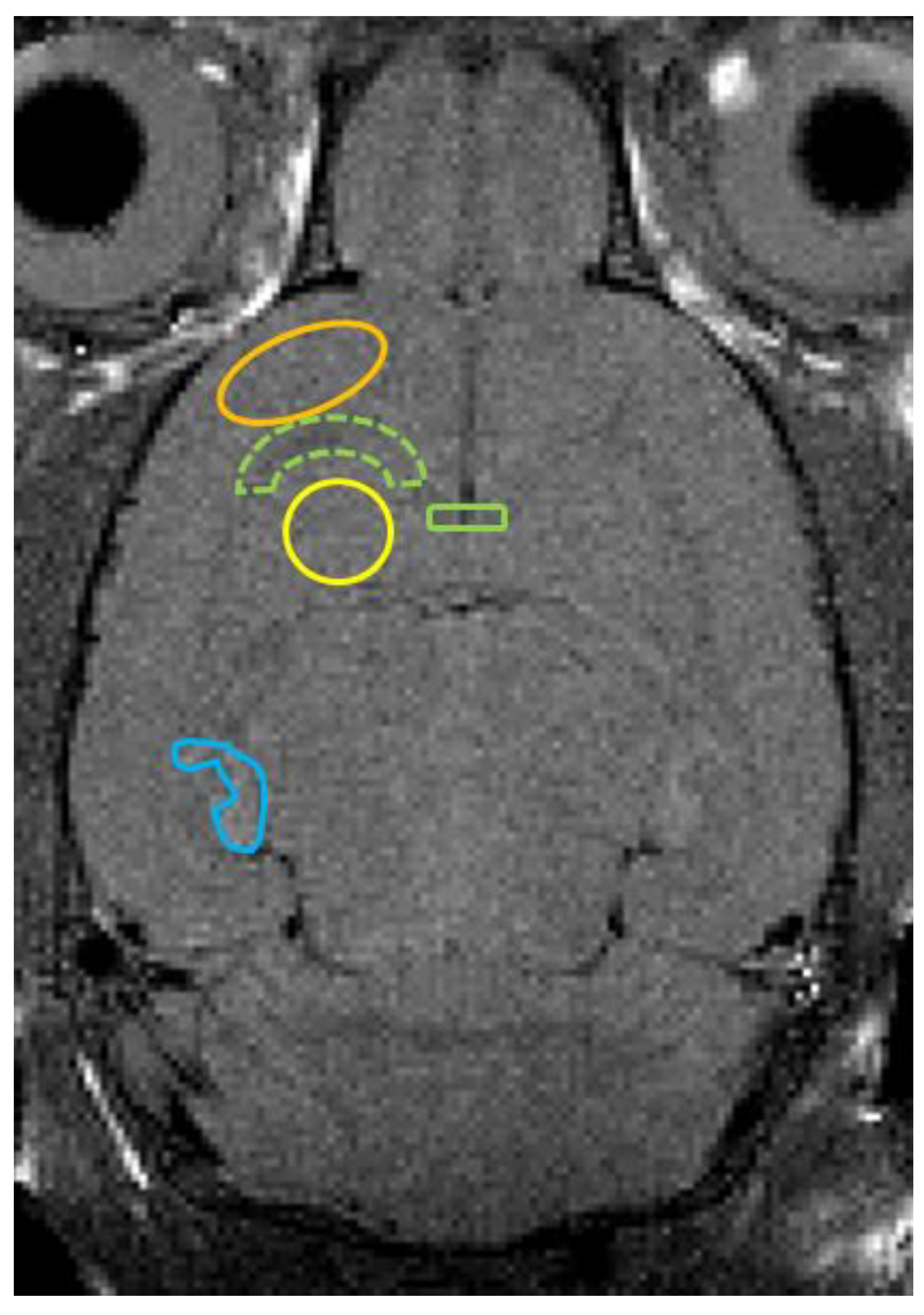
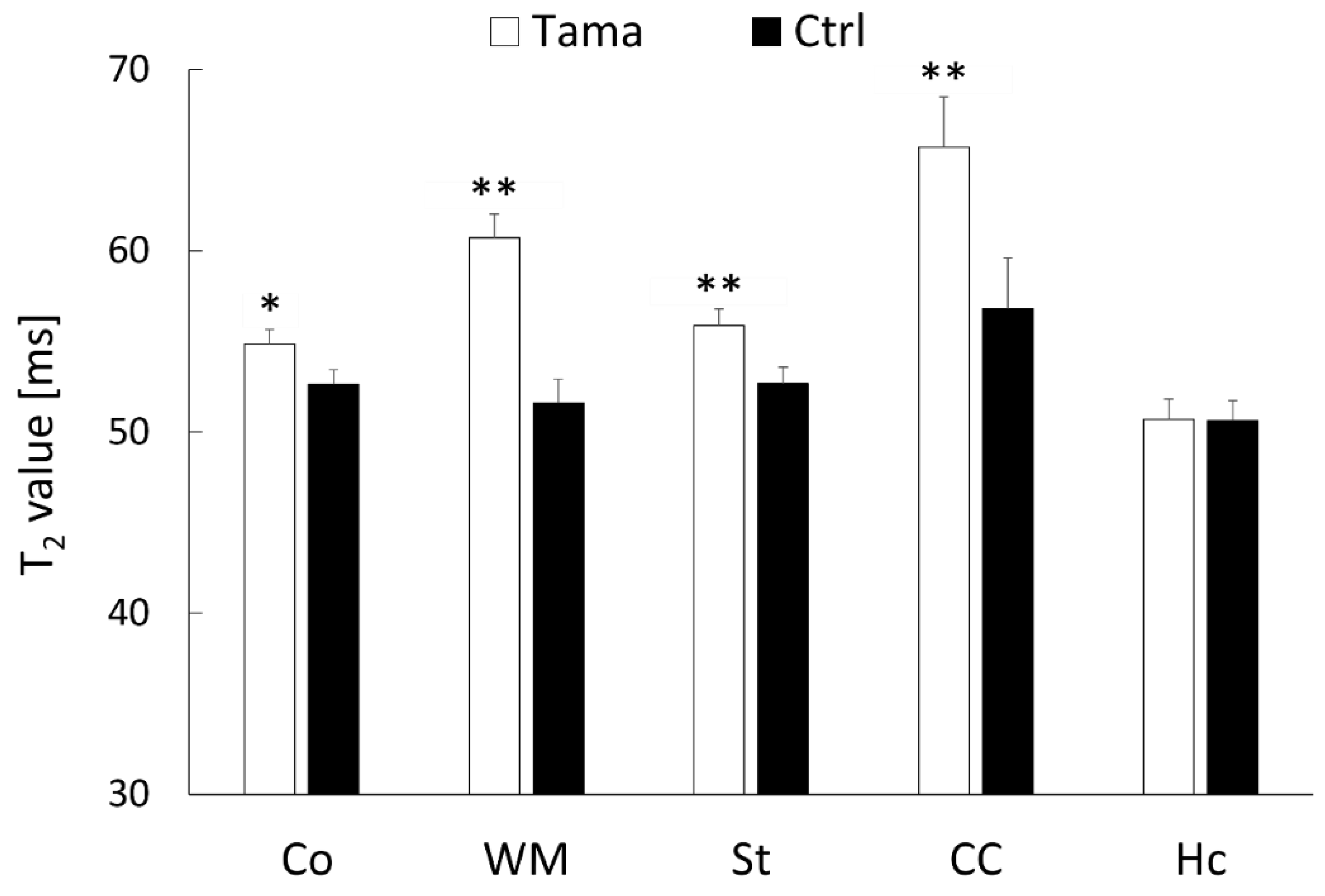
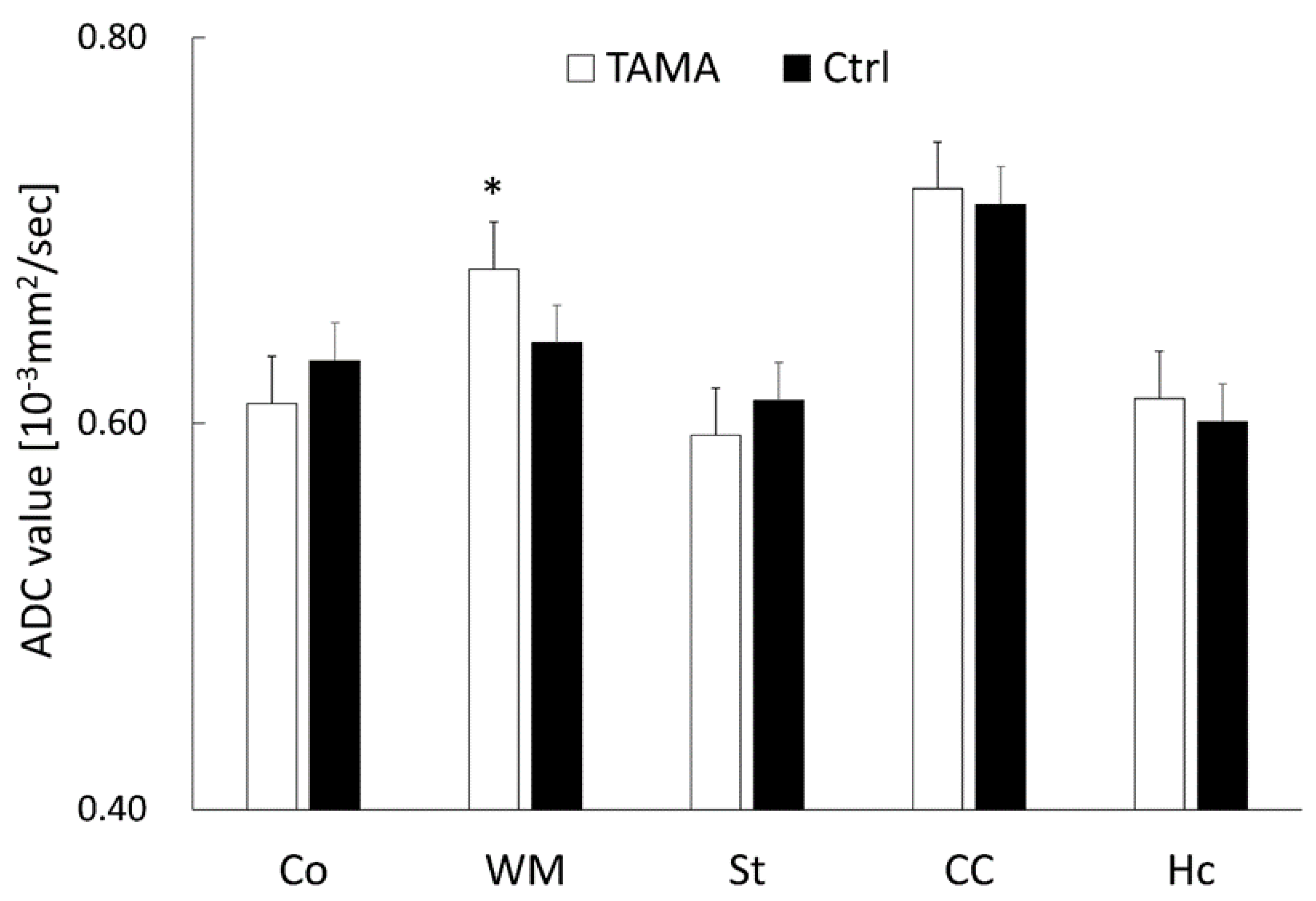
3.1. MRI Findings, T2 Value Measurements, and ADC Value Measurements
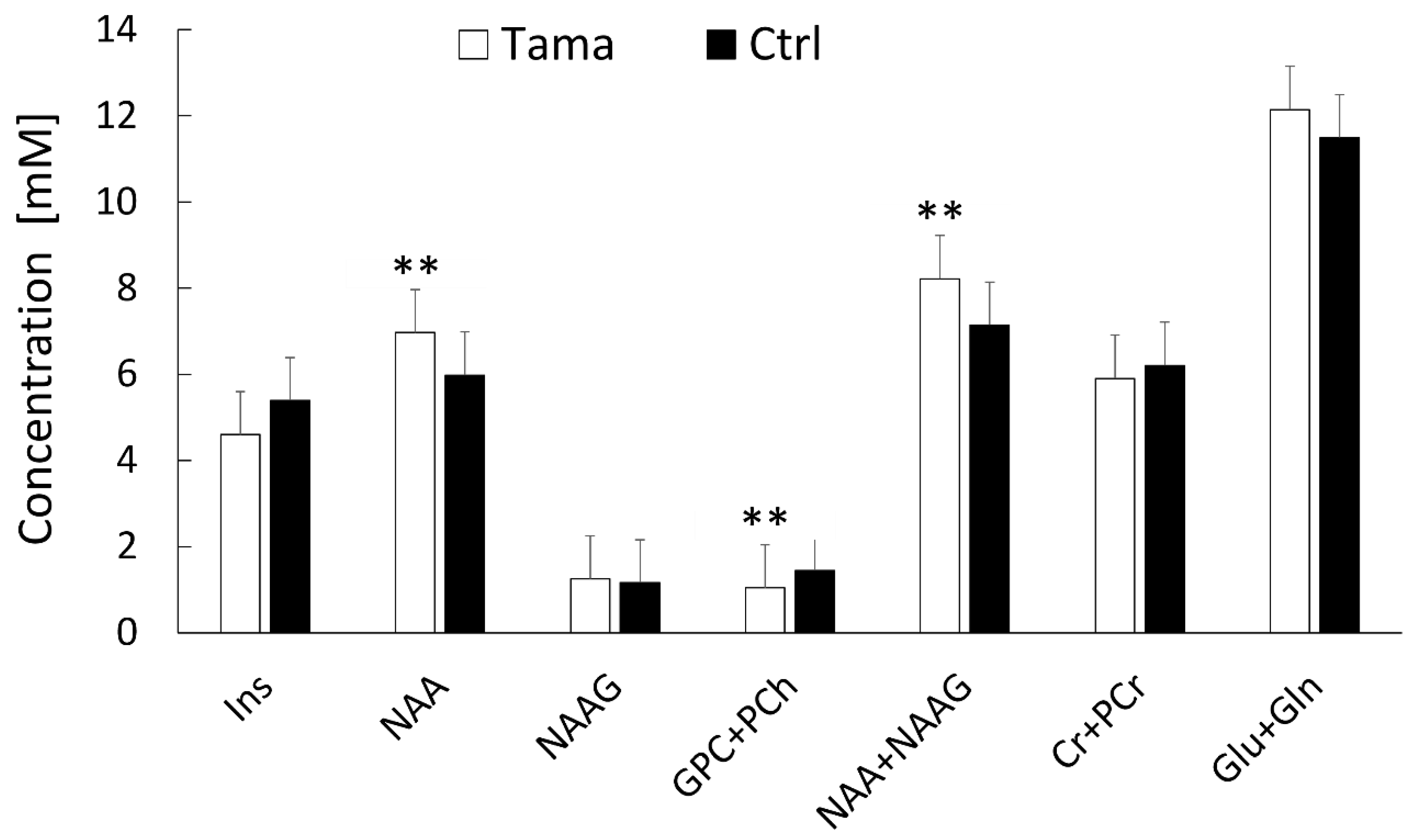
3.2. Brain Metabolites Measured Using 1H-MRS
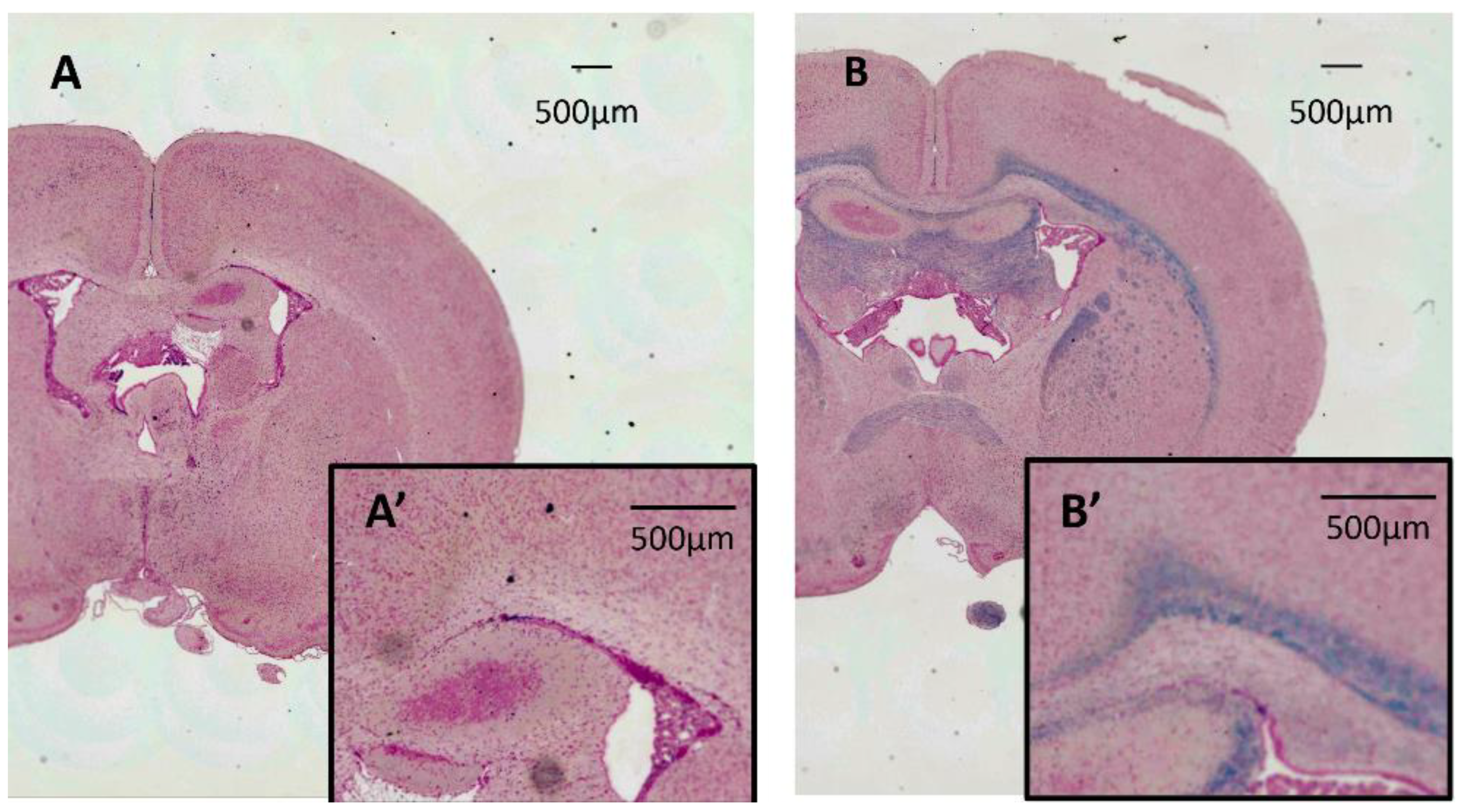
3.3. LFB Staining
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, Y.; Morrison, B.M.; Li, Y.; Lengacher, S.; Farah, M.H.; Hoffman, P.N.; Liu, Y.; Tsingalia, A.; Jin, L.; Zhang, P.W.; et al. Oligodendroglia metabolically support axons and contribute to neurodegeneration. Nature 2012, 487, 443–448. [Google Scholar] [CrossRef]
- Koeppen, A.H.; Robitaille, Y. Pelizaeus-Merzbacher disease. J. Neuropathol. Exp. Neurol. 2002, 61, 747–759. [Google Scholar] [CrossRef] [Green Version]
- Inoue, K.; Tanaka, H.; Scaglia, F.; Araki, A.; Shaffer, L.G.; Lupski, J.R. Compensating for central nervous system dysmyelination: Females with a proteolipid protein gene duplication and sustained clinical improvement. Ann. Neurol. 2001, 50, 747–754. [Google Scholar] [CrossRef]
- Seitelberger, F. Neuropathology and genetics of Pelizaeus-Merzbacher disease. Brain Pathol. 1995, 5, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Nave, K.A.; Lai, C.; Bloom, F.E.; Milner, R.J. Jimpy mutant mouse: A 74-base deletion in the mRNA for myelin proteolipid protein and evidence for a primary defect in RNA splicing. Proc. Natl. Acad. Sci. USA 1986, 83, 9264–9268. [Google Scholar] [CrossRef] [Green Version]
- Gow, A.; Lazzarini, R.A. A cellular mechanism governing the severity of Pelizaeus-Merzbacher disease. Nat. Genet. 1996, 13, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Dentinger, M.P.; Barron, K.D.; Csiza, C.K. Glial and axonal development in optic nerve of myelin deficient rat mutant. Brain Res. 1985, 344, 255–266. [Google Scholar] [CrossRef]
- Marciano, F.F.; Gocht, A.; Dentinger, M.P.; Hof, L.; Csiza, C.K.; Barron, K.D. Axonal regrowth in the amyelinated optic nerve of the myelin-deficient rat: Ultrastructural observations and effects of ganglioside administration. J. Comp. Neurol. 1990, 295, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Csiza, C.K.; de Lahunta, A. Myelin deficiency (md): A neurologic mutant in the Wistar rat. Am. J. Pathol. 1979, 95, 215–224. [Google Scholar]
- Fujiyoshi, K.; Hikishima, K.; Nakahara, J.; Tsuji, O.; Hata, J.; Konomi, T.; Nagai, T.; Shibata, S.; Kaneko, S.; Iwanami, A.; et al. Application of q-Space Diffusion MRI for the Visualization of White Matter. J. Neurosci. 2016, 36, 2796–2808. [Google Scholar] [CrossRef] [Green Version]
- Bar-Shir, A.; Duncan, I.D.; Cohen, Y. QSI and DTI of excised brains of the myelin-deficient rat. Neuroimage 2009, 48, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Takanashi, J.; Saito, S.; Aoki, I.; Barkovich, A.J.; Ito, Y.; Inoue, K. Increased N-acetylaspartate in model mouse of Pelizaeus-Merzbacher disease. J. Magn. Reson. Imaging 2012, 35, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.S.; Marques, J.P. Multi-compartment relaxometry and diffusion informed myelin water imaging—Promises and challenges of new gradient echo myelin water imaging methods. Neuroimage 2020, 221, 117159. [Google Scholar] [CrossRef] [PubMed]
- Marcuard, S.P.; Ehrinpreis, M.N.; Fitter, W.F. Gold induced ulcerative proctitis: Report and review of the literature. J. Rheumatol. 1987, 14, 142–144. [Google Scholar]
- O’Brien, J.S. Stability of the Myelin Membrane. Science 1965, 147, 1099–1107. [Google Scholar] [CrossRef]
- Simons, M.; Nave, K.A. Oligodendrocytes: Myelination and Axonal Support. Cold Spring Harb. Perspect. Biol. 2015, 8, a020479. [Google Scholar] [CrossRef]
- Bruvo, M.; Mahmood, F. Apparent diffusion coefficient measurement of the parotid gland parenchyma. Quant. Imaging Med. Surg. 2021, 11, 3812–3829. [Google Scholar] [CrossRef]
- Chung, S.H.; Yu, H.; Su, M.Y.; Cerussi, A.E.; Tromberg, B.J. Molecular imaging of water binding state and diffusion in breast cancer using diffuse optical spectroscopy and diffusion weighted MRI. J. Biomed. Opt. 2012, 17, 071304. [Google Scholar] [CrossRef] [Green Version]
- Saito, S.; Takahashi, Y.; Ohki, A.; Shintani, Y.; Higuchi, T. Early detection of elevated lactate levels in a mitochondrial disease model using chemical exchange saturation transfer (CEST) and magnetic resonance spectroscopy (MRS) at 7T-MRI. Radiol. Phys. Technol. 2019, 12, 46–54. [Google Scholar] [CrossRef]
- Ohki, A.; Saito, S.; Hata, J.; Okano, H.J.; Higuchi, T.; Fukuchi, K. Neurite orientation dispersion and density imaging for evaluating the severity of neonatal hypoxic-ischemic encephalopathy in rats. Magn. Reson. Imaging 2019, 62, 214–219. [Google Scholar] [CrossRef]
- Onishi, R.; Sawaya, R.; Tsuji, K.; Arihara, N.; Ohki, A.; Ueda, J.; Hata, J.; Saito, S. Evaluation of Temozolomide Treatment for Glioblastoma Using Amide Proton Transfer Imaging and Diffusion MRI. Cancers 2022, 14, 1907. [Google Scholar] [CrossRef]
- Provencher, S.W. Automatic quantitation of localized in vivo 1H spectra with LCModel. NMR Biomed. 2001, 14, 260–264. [Google Scholar] [CrossRef]
- Niessen, H.G.; Angenstein, F.; Vielhaber, S.; Frisch, C.; Kudin, A.; Elger, C.E.; Heinze, H.J.; Scheich, H.; Kunz, W.S. Volumetric magnetic resonance imaging of functionally relevant structural alterations in chronic epilepsy after pilocarpine-induced status epilepticus in rats. Epilepsia 2005, 46, 1021–1026. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Paus, T.; Zijdenbos, A.; Worsley, K.; Collins, D.L.; Blumenthal, J.; Giedd, J.N.; Rapoport, J.L.; Evans, A.C. Structural maturation of neural pathways in children and adolescents: In vivo study. Science 1999, 283, 1908–1911. [Google Scholar] [CrossRef] [PubMed]
- Wiggins, R.C. Myelin development and nutritional insufficiency. Brain Res. 1982, 257, 151–175. [Google Scholar] [CrossRef]
- Downes, N.; Mullins, P. The development of myelin in the brain of the juvenile rat. Toxicol. Pathol. 2014, 42, 913–922. [Google Scholar] [CrossRef] [Green Version]
- Ono, J.; Kodaka, R.; Imai, K.; Itagaki, Y.; Tanaka, J.; Inui, K.; Nagai, T.; Sakurai, K.; Harada, K.; Okada, S. Evaluation of myelination by means of the T2 value on magnetic resonance imaging. Brain Dev. 1993, 15, 433–438. [Google Scholar] [CrossRef]
- De Stefano, N.; Filippi, M. MR spectroscopy in multiple sclerosis. J. Neuroimaging 2007, 17 (Suppl. 1), 31S–35S. [Google Scholar] [CrossRef]
- Sener, R.N. Pelizaeus-Merzbacher disease: Diffusion MR imaging and proton MR spectroscopy findings. J. Neuroradiol. 2004, 31, 138–141. [Google Scholar] [CrossRef]
- Cercignani, M.; Iannucci, G.; Filippi, M. Diffusion-weighted imaging in multiple sclerosis. Ital. J. Neurol. Sci. 1999, 20, S246–S249. [Google Scholar] [CrossRef]
- Phuttharak, W.; Galassi, W.; Laopaiboon, V.; Laopaiboon, M.; Hesselink, J.R. ADC measurements in various patterns of multiple sclerosis lesions. J. Med. Assoc. Thai. 2006, 89, 196–204. [Google Scholar]
- Degaonkar, M.N.; Jayasundar, R.; Jagannathan, N.R. Sequential diffusion-weighted magnetic resonance imaging study of lysophosphatidyl choline-induced experimental demyelinating lesion: An animal model of multiple sclerosis. J. Magn. Reson. Imaging 2002, 16, 153–159. [Google Scholar] [CrossRef]
- Takanashi, J. Neurochemistry of Hypomyelination Investigated with MR Spectroscopy. Magn. Reson. Med. Sci. 2015, 14, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Gow, A.; Southwood, C.M.; Lazzarini, R.A. Disrupted proteolipid protein trafficking results in oligodendrocyte apoptosis in an animal model of Pelizaeus-Merzbacher disease. J. Cell Biol. 1998, 140, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Hobson, G.M.; Garbern, J.Y. Pelizaeus-Merzbacher disease, Pelizaeus-Merzbacher-like disease 1, and related hypomyelinating disorders. Semin. Neurol. 2012, 32, 62–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanefeld, F.A.; Brockmann, K.; Pouwels, P.J.; Wilken, B.; Frahm, J.; Dechent, P. Quantitative proton MRS of Pelizaeus-Merzbacher disease: Evidence of dys- and hypomyelination. Neurology 2005, 65, 701–706. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishikawa, M.; Sawaya, R.; Hirayama, M.; Ueda, J.; Saito, S. Magnetic Resonance Imaging and Spectroscopy Analysis in a Pelizaeus–Merzbacher Disease Rat Model. Diagnostics 2022, 12, 1864. https://doi.org/10.3390/diagnostics12081864
Ishikawa M, Sawaya R, Hirayama M, Ueda J, Saito S. Magnetic Resonance Imaging and Spectroscopy Analysis in a Pelizaeus–Merzbacher Disease Rat Model. Diagnostics. 2022; 12(8):1864. https://doi.org/10.3390/diagnostics12081864
Chicago/Turabian StyleIshikawa, Maho, Reika Sawaya, Miki Hirayama, Junpei Ueda, and Shigeyoshi Saito. 2022. "Magnetic Resonance Imaging and Spectroscopy Analysis in a Pelizaeus–Merzbacher Disease Rat Model" Diagnostics 12, no. 8: 1864. https://doi.org/10.3390/diagnostics12081864
APA StyleIshikawa, M., Sawaya, R., Hirayama, M., Ueda, J., & Saito, S. (2022). Magnetic Resonance Imaging and Spectroscopy Analysis in a Pelizaeus–Merzbacher Disease Rat Model. Diagnostics, 12(8), 1864. https://doi.org/10.3390/diagnostics12081864