
Aberrant Whole Blood Gene Expression in the Lumen of Human Intracranial Aneurysms

Abstract
1. Introduction
2. Methods
2.1. Study Participants
2.2. Intra-Aneurysmal Blood Collection
2.3. Sample Processing and RNA Extraction
2.4. Quantitative Polymerase Chain Reaction (qPCR) Analysis
2.5. Correlation with IA Size and Instability
2.6. Statistical Analysis
3. Results
3.1. Study Participants
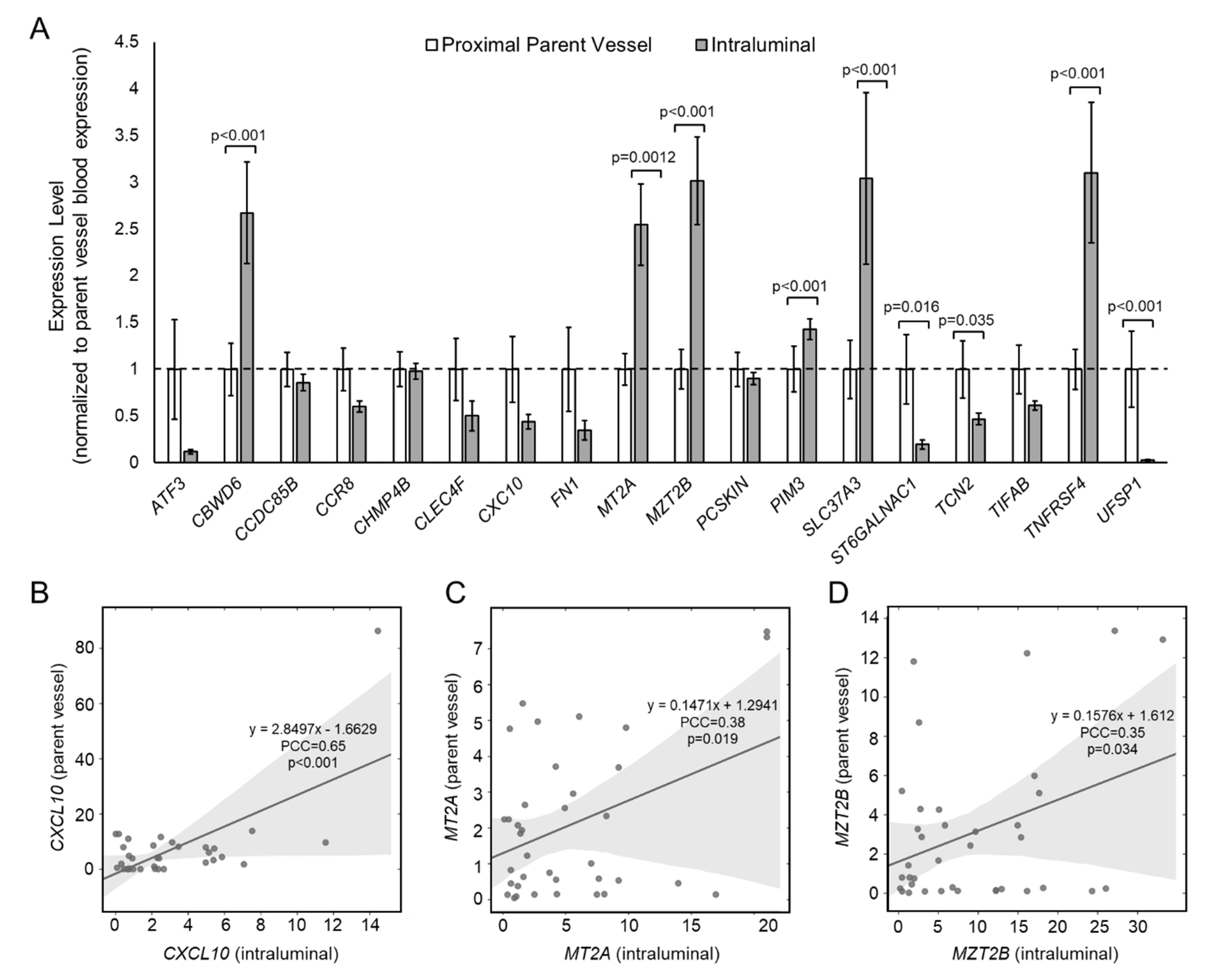
3.2. Differential Expression in the IA Sac versus the Proximal Parent Vessel
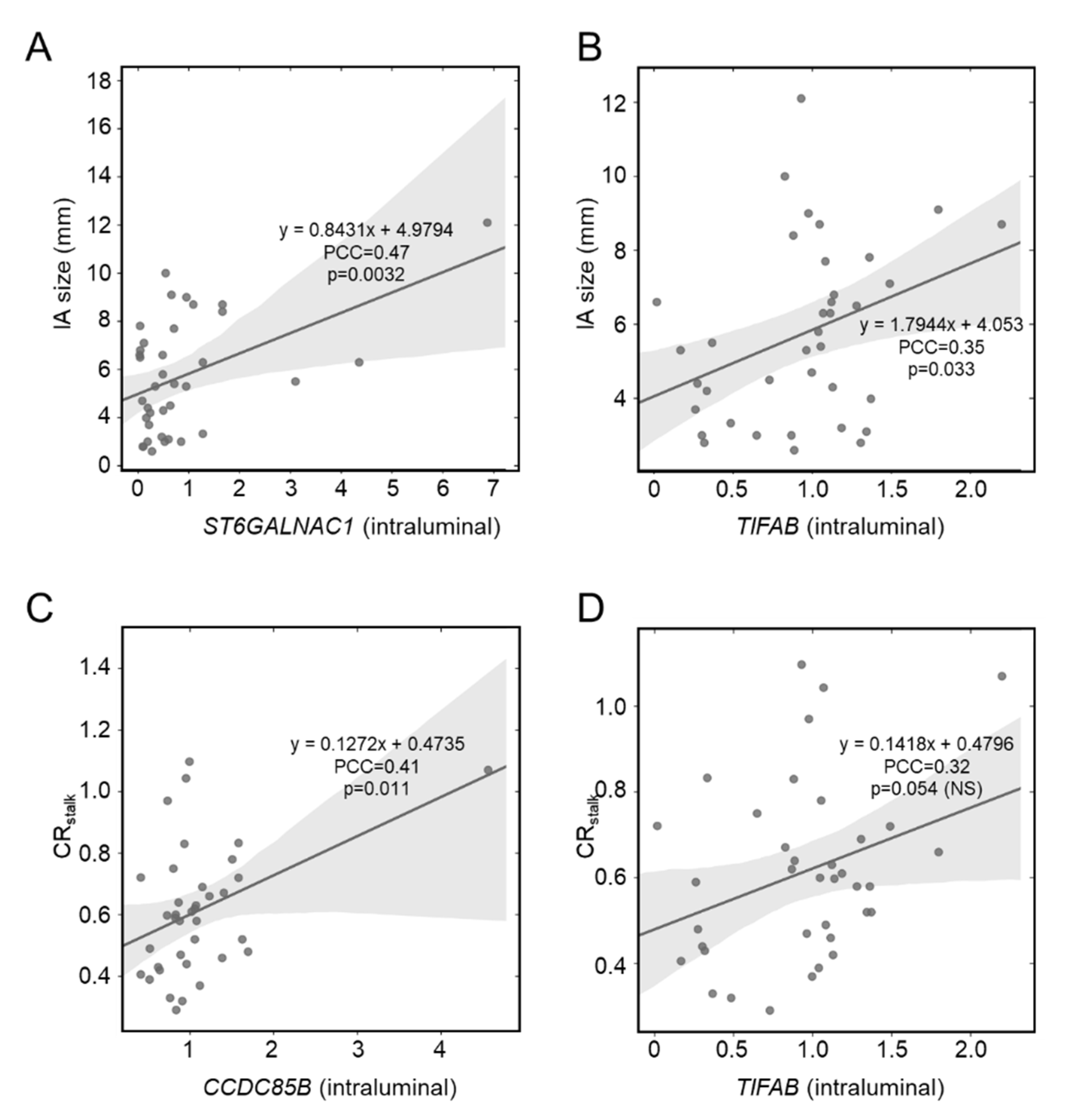
3.3. Correlation between IA Size and Intraluminal Gene Expression
3.4. Correlation between Aneurysmal VWE and Intraluminal Gene Expression
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hop, J.W.; Rinkel, G.J.; Algra, A.; Van Gijn, J. Case-Fatality Rates and Functional Outcome After Subarachnoid Hemorrhage. Stroke 1997, 28, 660–664. [Google Scholar] [CrossRef] [PubMed]
- Nieuwkamp, D.J.; Setz, L.E.; Algra, A.; Linn, F.H.; de Rooij, N.K.; Rinkel, G.J. Changes in case fatality of aneurysmal subarachnoid haemorrhage over time, according to age, sex, and region: A meta-analysis. Lancet Neurol. 2009, 8, 635–642. [Google Scholar] [CrossRef]
- Olafsson, E.; Hauser, W.A.; Gudmundsson, G. A population-based study of prognosis of ruptured cerebral aneurysm: Mortality and recurrence of subarachnoid hemorrhage. Neurology 1997, 48, 1191–1195. [Google Scholar] [CrossRef]
- Findlay, J.M.; Deagle, G.M. Causes of morbidity and mortality following intracranial aneurysm rupture. Can. J. Neurol. Sci. J. Can. Sci. Neurol. 1998, 25, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Ropper, A.H.; Zervas, N.T. Outcome 1 year after SAH from cerebral aneurysm. J. Neurosurg. 1984, 60, 909–915. [Google Scholar] [CrossRef]
- Proust, F.; Hannequin, D.; Langlois, O.; Freger, P.; Creissard, P. Causes of morbidity and mortality after ruptured aneurysm surgery in a series of 230 patients. The importance of control angiography. Stroke 1995, 26, 1553–1557. [Google Scholar] [CrossRef]
- Twitchell, S.; Abou-Al-Shaar, H.; Reese, J.; Karsy, M.; Eli, I.M.; Guan, J.; Taussky, P.; Couldwell, W.T. Analysis of cerebrovascular aneurysm treatment cost: Retrospective cohort comparison of clipping, coiling, and flow diversion. Neurosurg. Focus 2018, 44, E3. [Google Scholar] [CrossRef] [PubMed]
- Greving, J.P.; Rinkel, G.J.; Buskens, E.; Algra, A. Cost-effectiveness of preventive treatment of intracranial aneurysms: New data and uncertainties. Neurology 2009, 73, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Bederson, J.B.; Connolly, E.S.; Batjer, H.H.; Dacey, R.G.; Dion, J.E.; Diringer, M.; Duldner, J.E.; Harbaugh, R.; Patel, A.B.; Rosenwasser, R.H. Guidelines for the Management of Aneurysmal Subarachnoid Hemorrhage. Stroke 2009, 40, 994–1025. [Google Scholar] [CrossRef] [PubMed]
- Tutino, V.M.; Poppenberg, K.; Li, L.; Shallwani, H.; Jiang, K.; Jarvis, J.N.; Sun, Y.; Snyder, K.V.; Levy, E.I.; Siddiqui, A.H.; et al. Biomarkers from circulating neutrophil transcriptomes have potential to detect unruptured intracranial aneurysms. J. Transl. Med. 2018, 16, 1–16. [Google Scholar] [CrossRef]
- Tutino, V.M.; Poppenberg, K.; Jiang, K.; Jarvis, J.N.; Sun, Y.; Sonig, A.; Siddiqui, A.H.; Snyder, K.V.; Levy, E.I.; Kolega, J.; et al. Circulating neutrophil transcriptome may reveal intracranial aneurysm signature. PLoS ONE 2018, 13, e0191407. [Google Scholar] [CrossRef] [PubMed]
- Poppenberg, K.E.; Tutino, V.M.; Li, L.; Waqas, M.; June, A.; Chaves, L.; Jiang, K.; Jarvis, J.N.; Sun, Y.; Snyder, K.V.; et al. Classification models using circulating neutrophil transcripts can detect unruptured intracranial aneurysm. J. Transl. Med. 2020, 18, 392. [Google Scholar] [CrossRef]
- Tutino, V.M.; Poppenberg, K.E.; Damiano, R.J.; Patel, T.R.; Waqas, M.; Dmytriw, A.A.; Snyder, K.V.; Siddiqui, A.H.; Jarvis, J.N. Characterization of Long Non-coding RNA Signatures of Intracranial Aneurysm in Circulating Whole Blood. Mol. Diagn. Ther. 2020, 24, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Poppenberg, K.E.; Li, L.; Waqas, M.; Paliwal, N.; Jiang, K.; Jarvis, J.N.; Sun, Y.; Snyder, K.V.; Levy, E.I.; Siddiqui, A.H.; et al. Whole blood transcriptome biomarkers of unruptured intracranial aneurysm. PLoS ONE 2020, 15, e0241838. [Google Scholar] [CrossRef] [PubMed]
- Chalouhi, N.; Points, L.; Pierce, G.L.; Ballas, Z.; Jabbour, P.; Hasan, D. Localized increase of chemokines in the lumen of human cerebral aneurysms. Stroke 2013, 44, 2594–2597. [Google Scholar] [CrossRef] [PubMed]
- Wiebers, D.O. Unruptured intracranial aneurysms: Natural history, clinical outcome, and risks of surgical and endovascular treatment. Lancet 2003, 362, 103–110. [Google Scholar] [CrossRef]
- Roa, J.; Zanaty, M.; Osorno, C.; Ishii, D.; Bathla, G.; Ortega-Gutierrez, S.; Hasan, D.M.; Samaniego, E.A. Objective quantification of contrast enhancement of unruptured intracranial aneurysms: A high-resolution vessel wall imaging validation study. J. Neurosurg. 2021, 134, 862–869. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, D.; Zanaty, M.; Hudson, J.; Teferi, N.; Ishii, D.; Allan, L.; Jabbour, P.; Ortega-Gutierrez, S.; Samaniego, E.A.; Hasan, D.M. Plasma Soluble Human Elastin Fragments as an Intra-Aneurysmal Localized Biomarker for Ruptured Intracranial Aneurysm. J. Am. Hear. Assoc. 2018, 7, e010051. [Google Scholar] [CrossRef] [PubMed]
- Chalouhi, N.; Jabbour, P.; Zanaty, M.; Starke, R.M.; Torner, J.; Nakagawa, D.; Hasan, D.M. Sex Differential in 15-Hydroxyprostaglandin Dehydrogenase Levels in the Lumen of Human Intracranial Aneurysms. J. Am. Hear. Assoc. 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Mallona, I.; Weiss, J.; Marcos, E.-C. pcrEfficiency: A Web tool for PCR amplification efficiency prediction. BMC Bioinform. 2011, 12, 404. [Google Scholar] [CrossRef] [PubMed]
- Omodaka, S.; Endo, H.; Niizuma, K.; Fujimura, M.; Inoue, T.; Sato, K.; Sugiyama, S.I.; Tominaga, T. Quantitative Assessment of Circumferential Enhancement along the Wall of Cerebral Aneurysms Using MR Imaging. Am. J. Neuroradiol. 2016, 37, 1262–1266. [Google Scholar] [CrossRef] [PubMed]
- Tutino, V.; Zebraski, H.; Rajabzadeh-Oghaz, H.; Waqas, M.; Jarvis, J.; Bach, K.; Mokin, M.; Snyder, K.; Siddiqui, A.; Poppenberg, K. Identification of Circulating Gene Expression Signatures of Intracranial Aneurysm in Peripheral Blood Mononuclear Cells. Diagnostics 2021, 11, 1092. [Google Scholar] [CrossRef] [PubMed]
- Wuensch, K.L.; Evans, J.D. Straightforward Statistics for the Behavioral Sciences. J. Am. Stat. Assoc. 1996, 91, 1750. [Google Scholar] [CrossRef]
- Ishii, D.; Zanaty, M.; Roa, J.A.; Li, L.; Lu, Y.; Sabotin, R.; Allan, L.; Samaniego, E.A.; Hasan, D.M. Concentration of Lp(a) (Lipoprotein[a]) in Aneurysm Sac Is Associated With Wall Enhancement of Unruptured Intracranial Aneurysm. Stroke 2021, 52, 1465–1468. [Google Scholar] [CrossRef] [PubMed]
- Larsen, N.; Von Der Brelie, C.; Trick, D.; Riedel, C.; Lindner, T.; Madjidyar, J.; Jansen, O.; Synowitz, M.; Flüh, C. Vessel Wall Enhancement in Unruptured Intracranial Aneurysms: An Indicator for Higher Risk of Rupture? High-Resolution MR Imaging and Correlated Histologic Findings. Am. J. Neuroradiol. 2018, 39, 1617–1621. [Google Scholar] [CrossRef]
- Quan, K.; Song, J.; Yang, Z.; Wang, D.; An, Q.; Huang, L.; Liu, P.; Li, P.; Tian, Y.; Zhou, L.; et al. Validation of Wall Enhancement as a New Imaging Biomarker of Unruptured Cerebral Aneurysm. Stroke 2019, 50, 1570–1573. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; Kataoka, H.; Shimamura, M.; Nakagami, H.; Wakayama, K.; Moriwaki, T.; Ishibashi, R.; Nozaki, K.; Morishita, R.; Hashimoto, N. NF-κB Is a Key Mediator of Cerebral Aneurysm Formation. Circulation 2007, 116, 2830. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T. The Nuclear Factor NF-κB Pathway in Inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [PubMed]
- Chalouhi, N.; Ali, M.S.; Jabbour, P.M.; Tjoumakaris, S.I.; Gonzalez, L.F.; Rosenwasser, R.H.; Koch, W.J.; Dumont, A.S. Biology of Intracranial Aneurysms: Role of Inflammation. Br. J. Pharmacol. 2012, 32, 1659–1676. [Google Scholar] [CrossRef] [PubMed]
- Chyatte, D.; Bruno, G.; Desai, S.; Todor, D.R. Inflammation and Intracranial Aneurysms. Neurosurgery 1999, 45, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Medel, R.; Valle, E.; Amenta, P.S.; Dumont, A.S. Inflammation and intracranial aneurysms: Mechanisms of initiation, growth, and rupture. Neuroimmunol. Neuroinflamm. 2015, 2, 68–76. [Google Scholar] [CrossRef][Green Version]
- Meng, H.; Tutino, V.; Xiang, J.; Siddiqui, A. High WSS or Low WSS? Complex Interactions of Hemodynamics with Intracranial Aneurysm Initiation, Growth, and Rupture: Toward a Unifying Hypothesis. Am. J. Neuroradiol. 2013, 35, 1254–1262. [Google Scholar] [CrossRef]
- Zhang, X.; Ares, W.J.; Taussky, P.; Ducruet, A.F.; Grandhi, R. Role of matrix metalloproteinases in the pathogenesis of intracranial aneurysms. Neurosurg. Focus 2019, 47, E4. [Google Scholar] [CrossRef]
- Aoki, T.; Nishimura, M.; Kataoka, H.; Ishibashi, R.; Nozaki, K.; Hashimoto, N. Reactive oxygen species modulate growth of cerebral aneurysms: A study using the free radical scavenger edaravone and p47phox−/− mice. Lab. Investig. 2009, 89, 730–741. [Google Scholar] [CrossRef] [PubMed]
- Starke, R.M.; Chalouhi, N.; Ali, M.S.; Jabbour, P.M.; Tjoumakaris, S.I.; Gonzalez, L.F.; Rosenwasser, R.H.; Koch, W.J.; Dumont, A.S. The Role of Oxidative Stress in Cerebral Aneurysm Formation and Rupture. Curr. Neurovasc. Res. 2013, 10, 247–255. [Google Scholar] [CrossRef]
- Poppenberg, K.E.; Zebraski, H.R.; Avasthi, N.; Waqas, M.; Siddiqui, A.H.; Jarvis, J.N.; Tutino, V.M. Epigenetic landscapes of intracranial aneurysm risk haplotypes implicate enhancer function of endothelial cells and fibroblasts in dysregulated gene expression. BMC Med Genom. 2021, 14, 162. [Google Scholar] [CrossRef]
- Poppenberg, K.; Jiang, K.; Tso, M.K.; Snyder, K.V.; Siddiqui, A.H.; Kolega, J.; Jarvis, J.N.; Meng, H.; Tutino, V.M. Epigenetic landscapes suggest that genetic risk for intracranial aneurysm operates on the endothelium. BMC Med. Genom. 2019, 12, 149. [Google Scholar] [CrossRef] [PubMed]
- Rovin, B.H.; Dickerson, J.A.; Tan, L.C.; Hebert, C.A. Activation of nuclear factor-kappa B correlates with MCP-1 expression by human mesangial cells. Kidney Int. 1995, 48, 1263–1271. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kim, H.K.; Park, H.R.; Sul, K.H.; Chung, H.Y.; Chung, J. Induction of RANTES and CCR5 through NF-κB Activation via MAPK Pathway in Aged Rat Gingival Tissues. Biotechnol. Lett. 2006, 28, 17–23. [Google Scholar] [CrossRef]
- Bunting, K.; Rao, S.; Hardy, K.; Woltring, D.; Denyer, G.S.; Wang, J.; Gerondakis, S.; Shannon, M.F. Genome-Wide Analysis of Gene Expression in T Cells to Identify Targets of the NF-κB Transcription Factor c-Rel. J. Immunol. 2007, 178, 7097–7109. [Google Scholar] [CrossRef]
- Matsukura, S.; Stellato, C.; Plitt, J.R.; Bickel, C.; Miura, K.; Georas, S.N.; Casolaro, V.; Schleimer, R.P. Activation of Eotaxin Gene Transcription by NF-κB and STAT6 in Human Airway Epithelial Cells. J. Immunol. 1999, 163, 6876. [Google Scholar] [PubMed]
- Elliott, C.; Allport, V.; Loudon, J.; Wu, G.; Bennett, P. Nuclear factor-kappa B is essential for up-regulation of interleukin-8 expression in human amnion and cervical epithelial cells. Mol. Hum. Reprod. 2001, 7, 787–790. [Google Scholar] [CrossRef] [PubMed]
- Hata, K.; Andoh, A.; Shimada, M.; Fujino, S.; Bamba, S.; Araki, Y.; Okuno, T.; Fujiyama, Y.; Bamba, T. IL-17 stimulates inflammatory responses via NF-κB and MAP kinase pathways in human colonic myofibroblasts. Am. J. Physiol. Liver Physiol. 2002, 282, G1035–G1044. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, J.; Wang, J. A database and functional annotation of NF-kB target genes. Int. J. Clin. Exp. Med. 2016, 9, 7986–7995. [Google Scholar]
- Webb, G.J.; Hirschfield, G.; Lane, P.J.L. OX40, OX40L and Autoimmunity: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2015, 50, 312–332. [Google Scholar] [CrossRef]
- Mashimo, Y.; Suzuki, Y.; Hatori, K.; Tabara, Y.; Miki, T.; Tokunaga, K.; Katsuya, T.; Ogihara, T.; Yamada, M.; Takahashi, N.; et al. Association of TNFRSF4 gene polymorphisms with essential hypertension. J. Hypertens. 2008, 26, 902–913. [Google Scholar] [CrossRef] [PubMed]
- Van Wanrooij, E.J.; Van Puijvelde, G.H.; De Vos, P.; Yagita, H.; Van Berkel, T.J.; Kuiper, J. Interruption of the Tnfrsf4/Tnfsf4 (OX40/OX40L) Pathway Attenuates Atherogenesis in Low-Density Lipoprotein Receptor-Deficient Mice. Arter. Thromb. Vasc. Biol. 2007, 27, 204–210. [Google Scholar] [CrossRef]
- Jin, H.; Zhang, C.; Sun, C.; Zhao, X.; Tian, D.; Shi, W.; Tian, Y.; Liu, K.; Sun, G.; Xu, H.; et al. OX40 expression in neutrophils promotes hepatic ischemia/reperfusion injury. JCI Insight 2019, 4. [Google Scholar] [CrossRef]
- Ueno, H.; Blanco, P. OX40/OX40L axis: Not a friend in autoimmunity. Oncotarget 2015, 6, 21779–21780. [Google Scholar] [CrossRef]
- Willoughby, J.; Griffiths, J.; Tews, I.; Cragg, M.S. OX40: Structure and function—What questions remain? Mol. Immunol. 2017, 83, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Kurata, I.; Matsumoto, I.; Ohyama, A.; Osada, A.; Ebe, H.; Kawaguchi, H.; Kaneko, S.; Kondo, Y.; Tsuboi, H.; Tomioka, A.; et al. Potential involvement of OX40 in the regulation of autoantibody sialylation in arthritis. Ann. Rheum. Dis. 2019, 78, 1488–1496. [Google Scholar] [CrossRef]
- Lv, Y.-W.; Chen, Y.; Lv, H.-T.; Li, X.; Tang, Y.-J.; Qian, W.-G.; Xu, Q.-Q.; Sun, L.; Qian, G.-H.; Ding, Y.-Y.; et al. Kawasaki disease OX40–OX40L axis acts as an upstream regulator of NFAT signaling pathway. Pediatr. Res. 2019, 85, 835–840. [Google Scholar] [CrossRef]
- Pan, Y.; Huang, J.; Xing, R.; Yin, X.; Cui, J.; Li, W.; Yu, J.; Lu, Y. Metallothionein 2A inhibits NF-κB pathway activation and predicts clinical outcome segregated with TNM stage in gastric cancer patients following radical resection. J. Transl. Med. 2013, 11, 173. [Google Scholar] [CrossRef] [PubMed]
- Leng, W.; Lei, X.; Meng, H.; Ouyang, X.; Liang, Z. EOLA1 Inhibits Lipopolysaccharide-Induced Vascular Cell Adhesion Molecule-1 Expression by Association with MT2A in ECV304 Cells. Int. J. Inflamm. 2015, 2015, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lavorgna, A.; Harhaj, E.W. EBV LMP1: New and shared pathways to NF-κB activation. Proc. Natl. Acad. Sci. USA 2012, 109, 2188–2189. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, C.; Senba, M.; Hashimoto, T.; Imaizumi, A.; Mori, N. Expression and significance of Pim-3 kinase in adult T-cell leukemia. Eur. J. Haematol. 2017, 99, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Iwaya, T.; Sawada, G.; Amano, S.; Kume, K.; Ito, C.; Endo, F.; Konosu, M.; Shioi, Y.; Akiyama, Y.; Takahara, T.; et al. Downregulation of ST6GALNAC1 is associated with esophageal squamous cell carcinoma development. Int. J. Oncol. 2016, 50, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-Y.; Cao, Y.-X.; Zhou, X.; Wei, B.; Zhan, L.; Sun, S.-Y. Stimulative role of ST6GALNAC1 in proliferation, migration and invasion of ovarian cancer stem cells via the Akt signaling pathway. Cancer Cell Int. 2019, 19, 86. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Hirohashi, Y.; Murai, A.; Nishidate, T.; Okita, K.; Wang, L.; Ikehara, Y.; Satoyoshi, T.; Usui, A.; Kubo, T.; et al. ST6GALNAC1 plays important roles in enhancing cancer stem phenotypes of colorectal cancer via the Akt pathway. Oncotarget 2017, 8, 112550–112564. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.-C.; Korean Kawasaki Disease Genetics Consortium; Kim, J.-J.; Yu, J.J.; Yun, S.W.; Yoon, K.L.; Lee, K.-Y.; Kil, H.-R.; Kim, G.B.; Han, M.-K.; et al. Identification of the TIFAB Gene as a Susceptibility Locus for Coronary Artery Aneurysm in Patients with Kawasaki Disease. Pediatr. Cardiol. 2018, 40, 483–488. [Google Scholar] [CrossRef]
- Niederkorn, M.; Agarwal, P.; Starczynowski, D.T. TIFA and TIFAB: FHA-domain proteins involved in inflammation, hematopoiesis, and disease. Exp. Hematol. 2020, 90, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Gao, Y.; Yu, J.; Jiang, G.; Zhang, X.; Lin, X.; Han, Q.; Rong, X.; Xu, H.-T.; Li, Q.; et al. CCDC85B promotes non-small cell lung cancer cell proliferation and invasion. Mol. Carcinog. 2018, 58, 126–134. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Characteristic | Value |
|---|---|
| Age (average years ± SD) | 63.0 ± 11.7 |
| Female gender (n/ntotal) | 25/31 (80.6%) |
| Smoking (n/ntotal) | 19/31 (61.3%) |
| Hypertension (n/ntotal) | 24/31 (77.4%) |
| Hyperlipidemia (n/ntotal) | 14/31 (45.2%) |
| Diabetes mellitus (n/ntotal) | 2/31 (6.5%) |
| Patients with multiple IAs (n/ntotal) | 4/31 (12.9%) |
| Total number of IAs (n/ntotal) | 37/31 |
| IA location (n/ntotal_IA) | |
| ACA | 1/37 (2.7%) |
| ACom | 7/37 (18.9%) |
| BT | 5/37 (13.5%) |
| ICA | 14/37 (37.8%) |
| MCA | 6/37 (16.2%) |
| PCom | 2/37 (5.4%) |
| PICA | 1/37 (2.7%) |
| VA | 1/37 (2.7%) |
| Pt. ID | IA ID | PV ID | Location | Max. D | CRstalk |
|---|---|---|---|---|---|
| 1 | IA1 | PV1 | R MCA | 7.8 | 0.58 |
| 2 | IA2 | PV2 | R ICA (term.) | 8.7 | 1.07 |
| 3 | IA3 | PV3 | R ACA | 4.0 | 0.52 |
| 4 | IA4 | PV4 | BT | 7.1 | 0.72 |
| 5 | IA5 | PV5/6 | R ICA (paraop.) | 6.5 | 0.58 |
| 5 | IA6 | PV5/6 | R ICA (paraop.) | 6.3 | 0.46 |
| 5 | IA7 | PV7 | L ICA (Op.) | 2.8 | 0.69 |
| 6 | IA8 | PV8 | R ICA (term.) | 3.1 | 0.52 |
| 7 | IA9 | PV9 | ACom | 5.8 | 0.39 |
| 8 | IA10 | PV10 | L PCom | 8.4 | 0.83 |
| 9 | IA11 | PV11 | L ICA (term.) | 6.6 | 0.63 |
| 10 | IA12 | PV12 | ACom | 5.3 | 0.47 |
| 11 | IA13 | PV13 | ACom | 9.0 | 0.97 |
| 12 | IA14 | PV14 | BT | 3.3 | 0.32 |
| 13 | IA15 | PV15 | ACom | 3.2 | 0.61 |
| 14 | IA16 | PV16 | R ICA | 5.4 | 0.78 |
| 15 | IA17 | PV17 | L ICA (op.) | 3.7 | 0.59 |
| 16 | IA18 | PV18 | L ICA (op.) | 4.7 | 0.37 |
| 17 | IA19 | PV19/20 | L ICA | 9.1 | 0.66 |
| 17 | IA20 | PV19/20 | L MCA | 4.4 | 0.48 |
| 18 | IA21 | PV21 | PICA | 4.3 | 0.42 |
| 19 | IA22 | PV22 | R MCA | 5.5 | 0.33 |
| 20 | IA23 | PV23 | R PCom | 7.7 | 0.49 |
| 21 | IA24 | PV24 | R ICA (paraop.) | 3.0 | 0.62 |
| 22 | IA25 | PV25 | ACom | 2.8 | 0.43 |
| 23 | IA26 | PV26 | ACom | 8.7 | 0.60 |
| 24 | IA27 | PV27/28/29 | R MCA | 2.6 | 0.64 |
| 24 | IA28 | PV27/28/29 | R MCA | 3.0 | 0.44 |
| 24 | IA29 | PV27/28/29 | R ICA | 3.0 | 0.75 |
| 25 | IA30 | PV30 | BT | 12.1 | 1.10 |
| 26 | IA31 | PV31 | BT | 5.3 | 0.41 |
| 27 | IA32 | PV32 | R MCA | 6.6 | 0.72 |
| 28 | IA33 | PV33 | ACom | 4.5 | 0.29 |
| 29 | IA34 | PV34 | BT | 10.0 | 0.67 |
| 30 | IA35 | PV35 | R ICA (paraop.) | 4.2 | 0.83 |
| 31 | IA36 | PV36 | R ICA (paraop.) | 6.8 | 0.60 |
| 31 | IA37 | PV37 | L. VA | 6.3 | 1.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tutino, V.M.; Lu, Y.; Ishii, D.; Poppenberg, K.E.; Rajabzadeh-Oghaz, H.; Siddiqui, A.H.; Hasan, D.M. Aberrant Whole Blood Gene Expression in the Lumen of Human Intracranial Aneurysms. Diagnostics 2021, 11, 1442. https://doi.org/10.3390/diagnostics11081442
Tutino VM, Lu Y, Ishii D, Poppenberg KE, Rajabzadeh-Oghaz H, Siddiqui AH, Hasan DM. Aberrant Whole Blood Gene Expression in the Lumen of Human Intracranial Aneurysms. Diagnostics. 2021; 11(8):1442. https://doi.org/10.3390/diagnostics11081442
Chicago/Turabian StyleTutino, Vincent M., Yongjun Lu, Daizo Ishii, Kerry E. Poppenberg, Hamidreza Rajabzadeh-Oghaz, Adnan H. Siddiqui, and David M. Hasan. 2021. "Aberrant Whole Blood Gene Expression in the Lumen of Human Intracranial Aneurysms" Diagnostics 11, no. 8: 1442. https://doi.org/10.3390/diagnostics11081442
APA StyleTutino, V. M., Lu, Y., Ishii, D., Poppenberg, K. E., Rajabzadeh-Oghaz, H., Siddiqui, A. H., & Hasan, D. M. (2021). Aberrant Whole Blood Gene Expression in the Lumen of Human Intracranial Aneurysms. Diagnostics, 11(8), 1442. https://doi.org/10.3390/diagnostics11081442