Expression of Neurokinin B Receptor in the Gingival Squamous Cell Carcinoma Bone Microenvironment

 , ,
, ,
Abstract
1. Introduction
2. Patients and Methods
2.1. Patients
2.2. Histopathological Analysis
2.3. Immunohistochemical Analysis
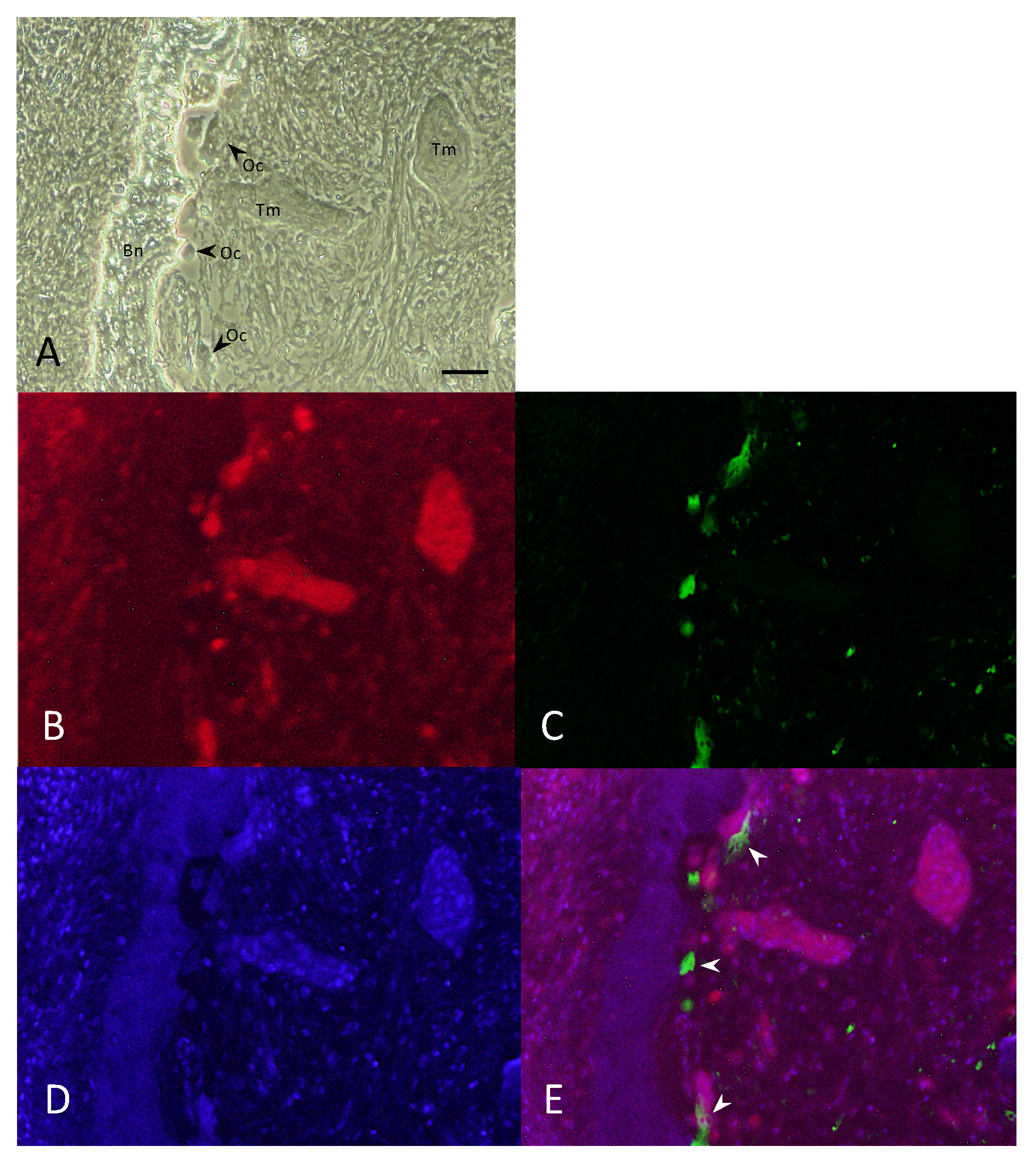
2.4. Double-Fluorescent Immunohistochemical Analysis
2.5. Quantification of NK-3R in Tissue Sections
2.6. Statistical Analysis
3. Results
3.1. Patient Demographics
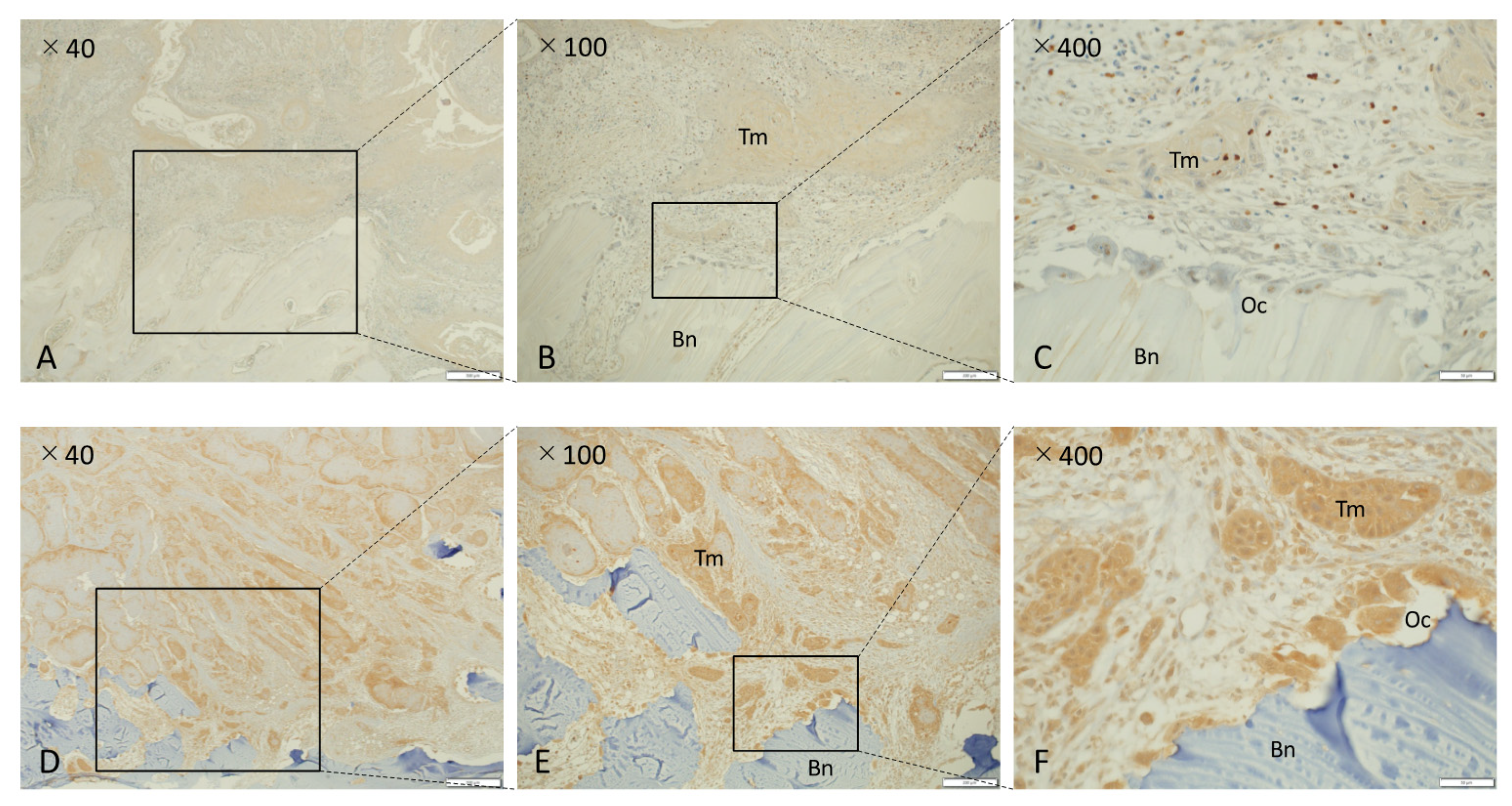
3.2. NK-3R Expression in Tumor Cells and Clinicopathological Characteristics in Human Gingival SCC
3.3. NK-3R Expression in Osteoclasts and Clinicopathological Characteristics in Human Gingival SCC
3.4. NK-3R Expression in Osteolytic Gingival SCC and Osteoclasts
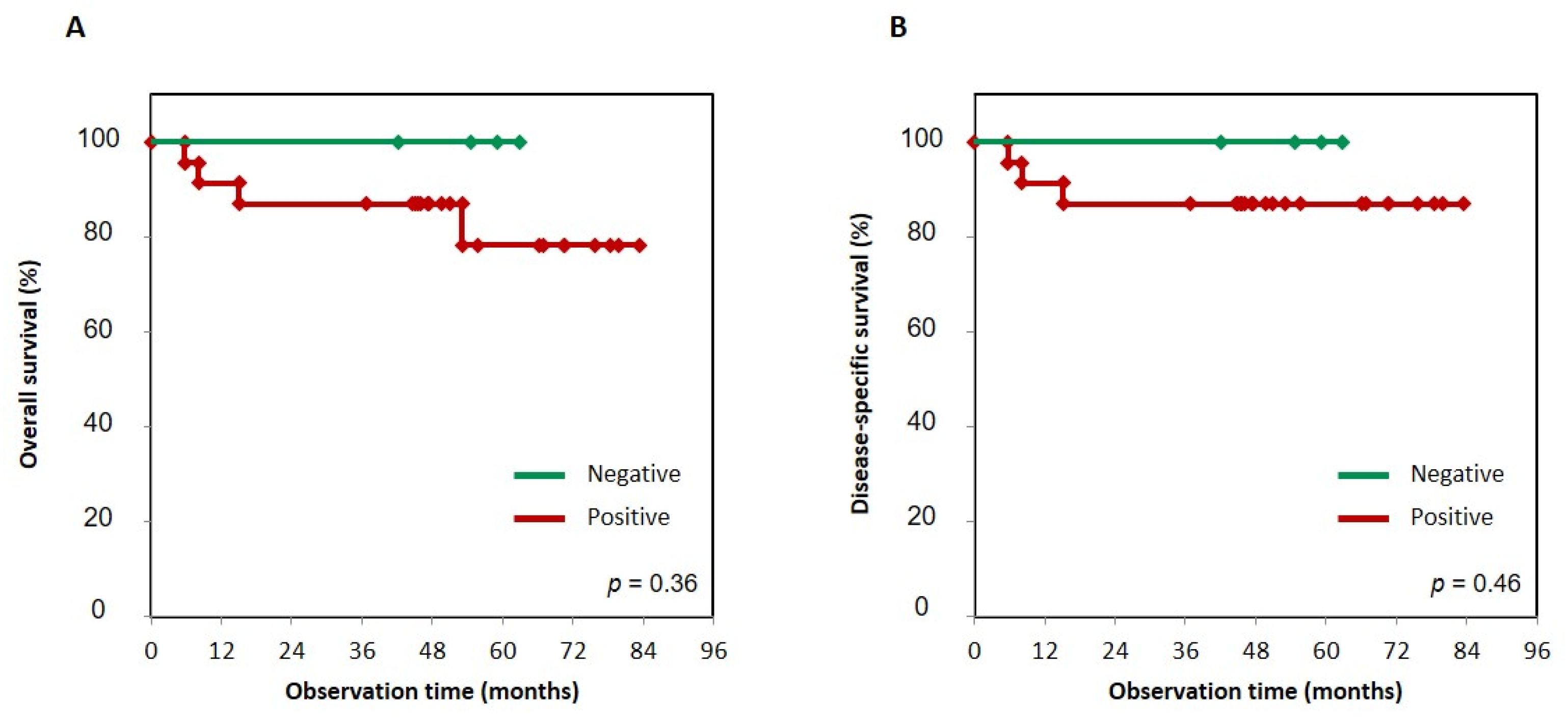
3.5. Survival Analysis
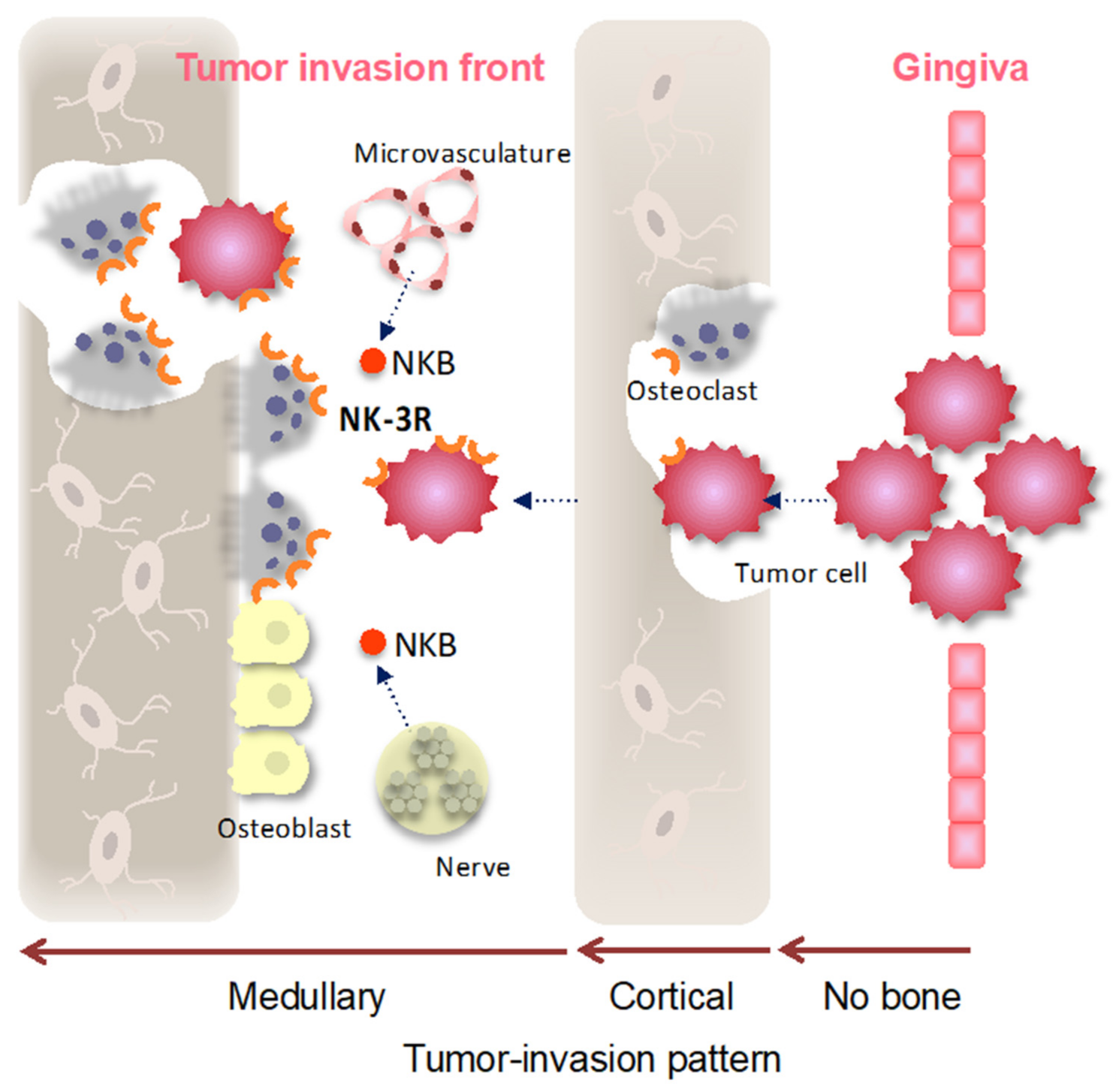
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sloan, P.; Gale, N.; Hunter, K.; Lingen, M.; Nylander, K.; Reibel, J.; Salo, T.; Zain, R.B. Malignant surface epithelial tumours. World Health Organization classification of tumours. In WHO Classification of Head and Neck Tumors, 4th ed.; El-Naggar, A.K., Chan, J.K.C., Grandis, J.R., Tanaka, T., Slootweg, P.J., Eds.; IARC: Lyon, France, 2017; pp. 109–111. [Google Scholar]
- Lee, K.C.; Chuang, S.K.; Philipone, E.M.; Peters, S.M. Which Clinicopathologic Factors Affect the Prognosis of Gingival Squamous Cell Carcinoma: A Population Analysis of 4,345 Cases. J. Oral Maxillofac. Surg. 2019, 77, 986–993. [Google Scholar] [CrossRef]
- Farhood, Z.; Simpson, M.; Ward, G.M.; Walker, R.J.; Osazuwa-Peters, N. Does anatomic subsite influence oral cavity cancer mortality? A SEER database analysis. Laryngoscope 2019, 129, 1400–1406. [Google Scholar] [CrossRef] [PubMed]
- Fives, C.; Nae, A.; Roche, P.; O’Leary, G.; Fitzgerald, B.; Feeley, L.; Sheahan, P. Impact of mandibular invasion on prognosis in oral squamous cell carcinoma four centimeters or less in size. Laryngoscope 2017, 127, 849–854. [Google Scholar] [CrossRef]
- Fried, D.; Mullins, B.; Weissler, M.; Shores, C.; Zanation, A.; Hackman, T.; Shockley, W.; Hayes, N.; Chera, B.S. Prognostic significance of bone invasion for oral cavity squamous cell carcinoma considered T1/T2 by American joint committee on cancer size criteria. Head Neck 2014, 36, 776–781. [Google Scholar] [CrossRef] [PubMed]
- Okura, M.; Yanamoto, S.; Umeda, M.; Otsuru, M.; Ota, Y.; Kurita, H.; Kamata, T.; Kirita, T.; Yamakawa, N.; Yamashita, T.; et al. Prognostic and staging implications of mandibular canal invasion in lower gingival squamous cell carcinoma. Cancer Med. 2016, 5, 3378–3385. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.; Shimo, T.; Murase, Y.; Takabatake, K.; Kishimoto, K.; Ibaragi, S.; Yoshioka, N.; Okui, T.; Nagatsuka, H.; Sasaki, A. The Prognostic Implications of Bone Invasion in Gingival Squamous Cell Carcinoma. Anticancer Res. 2018, 38, 955–962. [Google Scholar] [PubMed]
- Honami, T.; Shimo, T.; Okui, T.; Kurio, N.; Hassan, N.M.; Iwamoto, M.; Sasaki, A. Sonic hedgehog signaling promotes growth of oral squamous cell carcinoma cells associated with bone destruction. Oral Oncol. 2012, 48, 49–55. [Google Scholar] [CrossRef]
- Shimo, T.; Matsumoto, K.; Takabatake, K.; Aoyama, E.; Takebe, Y.; Ibaragi, S.; Okui, T.; Kurio, N.; Takada, H.; Obata, K.; et al. The Role of Sonic Hedgehog Signaling in Osteoclastogenesis and Jaw Bone Destruction. PLoS ONE 2016, 11, e0151731. [Google Scholar] [CrossRef] [PubMed]
- Navarro, V.M. New insights into the control of pulsatile GnRH release: The role of Kiss1/neurokinin B neurons. Front. Endocrinol. 2012, 3, 48. [Google Scholar] [CrossRef] [PubMed]
- Skorupskaite, K.; George, J.T.; Veldhuis, J.D.; Millar, R.P.; Anderson, R.A. Interactions Between Neurokinin B and Kisspeptin in Mediating Estrogen Feedback in Healthy Women. J. Clin. Endocrinol. Metab. 2016, 101, 4628–4636. [Google Scholar] [CrossRef]
- Pennefather, J.N.; Lecci, A.; Candenas, M.L.; Patak, E.; Pinto, F.M.; Maggi, C.A. Tachykinins and tachykinin receptors: A growing family. Life Sci. 2004, 74, 1445–1463. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Smith, J.T.; Henry, B.; Rao, A.; Pereira, A.; Clarke, I.J. Expression of genes for Kisspeptin (KISS1), Neurokinin B (TAC3), Prodynorphin (PDYN), and gonadotropin inhibitory hormone (RFRP) across natural puberty in ewes. Physiol. Rep. 2020, 8, e14399. [Google Scholar] [CrossRef] [PubMed]
- Young, J.; Bouligand, J.; Francou, B.; Raffin-Sanson, M.L.; Gaillez, S.; Jeanpierre, M.; Grynberg, M.; Kamenicky, P.; Chanson, P.; Brailly-Tabard, S.; et al. TAC3 and TACR3 defects cause hypothalamic congenital hypogonadotropic hypogonadism in humans. J. Clin. Endocrinol. Metab. 2010, 95, 2287–2295. [Google Scholar] [CrossRef] [PubMed]
- Obata, K.; Shimo, T.; Okui, T.; Matsumoto, K.; Takada, H.; Takabatake, K.; Kunisada, Y.; Ibaragi, S.; Nagatsuka, H.; Sasaki, A. Tachykinin Receptor 3 Distribution in Human Oral Squamous Cell Carcinoma. Anticancer Res. 2016, 36, 6335–6341. [Google Scholar] [CrossRef][Green Version]
- Obata, K.; Shimo, T.; Okui, T.; Matsumoto, K.; Takada, H.; Takabatake, K.; Kunisada, Y.; Ibaragi, S.; Yoshioka, N.; Kishimoto, K.; et al. Role of Neurokinin 3 Receptor Signaling in Oral Squamous Cell Carcinoma. Anticancer Res. 2017, 37, 6119–6123. [Google Scholar] [PubMed]
- Brierley, J.D.; Gospodarowicz, M.K. TNM Classification of Malignant Tumours, 8th ed.; Wittekind, C., Ed.; Wiley-Blackwell: New York, NY, USA, 2016. [Google Scholar]
- Shimo, T.; Kubota, S.; Goda, T.; Yoshihama, Y.; Kurio, N.; Nishida, T.; Ng, P.S.; Endo, K.; Takigawa, M.; Sasakii, A. Clinical significance and pathogenic function of connective tissue growth factor (CTGF/CCN2) in osteolytic mandibular squamous cell carcinoma. Anticancer Res. 2008, 28, 2343–2348. [Google Scholar]
- Hoveyda, H.R.; Fraser, G.L.; Roy, M.O.; Dutheuil, G.; Batt, F.; El Bousmaqui, M.; Korac, J.; Lenoir, F.; Lapin, A.; Noel, S.; et al. Discovery and optimization of novel antagonists to the human neurokinin-3 receptor for the treatment of sex-hormone disorders (Part I). J. Med. Chem. 2015, 58, 3060–3082. [Google Scholar] [CrossRef] [PubMed]
- Rajapakshe, R.M.; Pallegama, R.W.; Jayasooriya, P.R.; Siriwardena, B.S.; Attygalla, A.M.; Hewapathirana, S.; Weerasinghe, J.U.; Dias, D.K.; Tilakaratne, W.M. A retrospective analysis to determine factors contributing to the survival of patients with oral squamous cell carcinoma. Cancer Epidemiol. 2015, 39, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Elmusrati, A.A.; Pilborough, A.E.; Khurram, S.A.; Lambert, D.W. Cancer-associated fibroblasts promote bone invasion in oral squamous cell carcinoma. Br. J. Cancer 2017, 117, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Zarrer, J.; Haider, M.T.; Smit, D.J.; Taipaleenmaki, H. Pathological Crosstalk between Metastatic Breast Cancer Cells and the Bone Microenvironment. Biomolecules 2020, 10, 337. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yu, C.; Gao, X.; Welte, T.; Muscarella, A.M.; Tian, L.; Zhao, H.; Zhao, Z.; Du, S.; Tao, J.; et al. The osteogenic niche promotes early-stage bone colonization of disseminated breast cancer cells. Cancer Cell 2015, 27, 193–210. [Google Scholar] [CrossRef] [PubMed]
- Haider, M.T.; Holen, I.; Dear, T.N.; Hunter, K.; Brown, H.K. Modifying the osteoblastic niche with zoledronic acid in vivo-potential implications for breast cancer bone metastasis. Bone 2014, 66, 240–250. [Google Scholar] [CrossRef]
- Mak, K.K.; Bi, Y.; Wan, C.; Chuang, P.T.; Clemens, T.; Young, M.; Yang, Y. Hedgehog signaling in mature osteoblasts regulates bone formation and resorption by controlling PTHrP and RANKL expression. Dev. Cell 2008, 14, 674–688. [Google Scholar] [CrossRef]
- Mundy, G.R.; Yang, X. Hedgehog coordination of postnatal osteoclast and osteoblast activities. Dev. Cell 2008, 14, 637–638. [Google Scholar] [CrossRef][Green Version]
- Ohba, S.; Kawaguchi, H.; Kugimiya, F.; Ogasawara, T.; Kawamura, N.; Saito, T.; Ikeda, T.; Fujii, K.; Miyajima, T.; Kuramochi, A.; et al. Patched1 haploinsufficiency increases adult bone mass and modulates Gli3 repressor activity. Dev. Cell 2008, 14, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Tucker, J.A.; Khullar, S.; Samant, R.S.; Shevde, L.A. Hedgehog signaling in tumor cells facilitates osteoblast-enhanced osteolytic metastases. PLoS ONE 2012, 7, e34374. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.B.; Yin, L.; Shi, C.; Li, Q.; Duan, P.; Huang, J.M.; Liu, C.; Wang, F.; Lewis, M.; Wang, Y.; et al. MAOA-Dependent Activation of Shh-IL6-RANKL Signaling Network Promotes Prostate Cancer Metastasis by Engaging Tumor-Stromal Cell Interactions. Cancer Cell 2017, 31, 368–382. [Google Scholar] [CrossRef]
- Li, X.; Jie, Q.; Zhang, H.; Zhao, Y.; Lin, Y.; Du, J.; Shi, J.; Wang, L.; Guo, K.; Li, Y.; et al. Disturbed MEK/ERK signaling increases osteoclast activity via the Hedgehog-Gli pathway in postmenopausal osteoporosis. Prog. Biophys. Mol. Biol. 2016, 122, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Amano, K.; Densmore, M.J.; Lanske, B. Conditional Deletion of Indian Hedgehog in Limb Mesenchyme Results in Complete Loss of Growth Plate Formation but Allows Mature Osteoblast Differentiation. J. Bone Miner. Res. 2015, 30, 2262–2272. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, Y.; Liao, Z.; Liu, Q.; Lei, X.; Li, M.; Saijilafu; Zhang, Z.; Hong, D.; Zhu, M.; et al. Genetic and pharmacological activation of Hedgehog signaling inhibits osteoclastogenesis and attenuates titanium particle-induced osteolysis partly through suppressing the JNK/c-Fos-NFATc1 cascade. Theranostics 2020, 10, 6638–6660. [Google Scholar] [CrossRef] [PubMed]
- Ichiki, T.; Kuroishi, K.N.; Gunjigake, K.K.; Kobayashi, S.; Goto, T. Neurokinin B activates the formation and bone resorption activity of rat osteoclasts. Neuropeptides 2011, 45, 239–244. [Google Scholar] [CrossRef]
- Abaci, A.; Catli, G.; Anik, A.; Kume, T.; Calan, O.G.; Dundar, B.N.; Bober, E. Significance of serum neurokinin B and kisspeptin levels in the differential diagnosis of premature thelarche and idiopathic central precocious puberty. Peptides 2015, 64, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Salman, H.; Shah, M.; Ali, A.; Aziz, A.; Vitale, S.G. Assessment of Relationship of Serum Neurokinin-B Level in the Pathophysiology of Pre-eclampsia: A Case-Control Study. Adv. Ther. 2018, 35, 1114–1121. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Zhu, Y.; Cai, H.; Zhang, Y.; Hou, J. Impact of lymphovascular invasion in oral squamous cell carcinoma: A meta-analysis. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2020, 131, 319–328.e1. [Google Scholar] [CrossRef]
- Pal, S.; Wu, J.; Murray, J.K.; Gellman, S.H.; Wozniak, M.A.; Keely, P.J.; Boyer, M.E.; Gomez, T.M.; Hasso, S.M.; Fallon, J.F.; et al. An antiangiogenic neurokinin-B/thromboxane A2 regulatory axis. J. Cell Biol. 2006, 174, 1047–1058. [Google Scholar] [CrossRef]
- Wang, T.; Chen, S.; Wang, S.; Shi, L.; Wang, C.; Zhang, J.; Gao, Y.; Li, G.; Qi, Y.; An, X.; et al. Targeting neurokinin-3 receptor: A novel anti-angiogenesis strategy for cancer treatment. Oncotarget 2017, 8, 40713–40723. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.D.; Li, S.Y.; Zhang, S.J.; Gupta, A.; Zhang, C.P.; Wang, L. The neural system regulates bone homeostasis via mesenchymal stem cells: A translational approach. Theranostics 2020, 10, 4839–4850. [Google Scholar] [CrossRef]
- Chen, S.H.; Zhang, B.Y.; Zhou, B.; Zhu, C.Z.; Sun, L.Q.; Feng, Y.J. Perineural invasion of cancer: A complex crosstalk between cells and molecules in the perineural niche. Am. J. Cancer Res. 2019, 9, 1–21. [Google Scholar] [PubMed]
- Moore, A.M.; Coolen, L.M.; Lehman, M.N. Kisspeptin/Neurokinin B/Dynorphin (KNDy) cells as integrators of diverse internal and external cues: Evidence from viral-based monosynaptic tract-tracing in mice. Sci. Rep. 2019, 9, 14768. [Google Scholar] [CrossRef]
- Li, X.; Ma, G.; Ma, Q.; Li, W.; Liu, J.; Han, L.; Duan, W.; Xu, Q.; Liu, H.; Wang, Z.; et al. Neurotransmitter substance P mediates pancreatic cancer perineural invasion via NK-1R in cancer cells. Mol. Cancer Res. 2013, 11, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Alkhadar, H.; Macluskey, M.; White, S.; Ellis, I. Perineural invasion in oral squamous cell carcinoma: Incidence, prognostic impact and molecular insight. J. Oral Pathol. Med. 2020, 49, 994–1003. [Google Scholar] [CrossRef] [PubMed]
- Bakst, R.L.; Glastonbury, C.M.; Parvathaneni, U.; Katabi, N.; Hu, K.S.; Yom, S.S. Perineural Invasion and Perineural Tumor Spread in Head and Neck Cancer. Int. J. Radiat. Oncol. Biol. Phys. 2019, 103, 1109–1124. [Google Scholar] [CrossRef] [PubMed]
- Michalek, J.; Brychtova, S.; Pink, R.; Dvorak, Z. Prognostic and predictive markers for perineural and bone invasion of oral squamous cell carcinoma. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc 2019, 163, 302–308. [Google Scholar] [CrossRef]
- Ge, C.; Huang, H.; Huang, F.; Yang, T.; Zhang, T.; Wu, H.; Zhou, H.; Chen, Q.; Shi, Y.; Sun, Y.; et al. Neurokinin-1 receptor is an effective target for treating leukemia by inducing oxidative stress through mitochondrial calcium overload. Proc. Natl. Acad. Sci. USA 2019, 116, 19635–19645. [Google Scholar] [CrossRef] [PubMed]
- Munoz, M.; Covenas, R. The Neurokinin-1 Receptor Antagonist Aprepitant, a New Drug for the Treatment of Hematological Malignancies: Focus on Acute Myeloid Leukemia. J. Clin. Med. 2020, 9, 1659. [Google Scholar] [CrossRef] [PubMed]
- Munoz, M.; Rosso, M.; Covenas, R. The NK-1 receptor antagonist L-732,138 induces apoptosis in human gastrointestinal cancer cell lines. Pharmacol. Rep. 2017, 69, 696–701. [Google Scholar] [CrossRef]
- Munoz, M.; Rosso, M.; Covenas, R. Neurokinin-1 Receptor Antagonists against Hepatoblastoma. Cancers 2019, 11, 1258. [Google Scholar] [CrossRef]
- Steinhoff, M.S.; von Mentzer, B.; Geppetti, P.; Pothoulakis, C.; Bunnett, N.W. Tachykinins and their receptors: Contributions to physiological control and the mechanisms of disease. Physiol. Rev. 2014, 94, 265–301. [Google Scholar] [CrossRef] [PubMed]
- McCarson, K.E.; Krause, J.E. NK-1 and NK-3 type tachykinin receptor mRNA expression in the rat spinal cord dorsal horn is increased during adjuvant or formalin-induced nociception. J. Neurosci. 1994, 14, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Roca, H.; McCauley, L.K. Inflammation and skeletal metastasis. Bonekey Rep. 2015, 4, 706. [Google Scholar] [CrossRef]
- Wu, M.Y.; Li, C.J.; Yiang, G.T.; Cheng, Y.L.; Tsai, A.P.; Hou, Y.T.; Ho, Y.C.; Hou, M.F.; Chu, P.Y. Molecular Regulation of Bone Metastasis Pathogenesis. Cell. Physiol. Biochem. 2018, 46, 1423–1438. [Google Scholar] [CrossRef] [PubMed]
- Novack, D.V. Editorial: Inflammatory Osteoclasts: A Different Breed of Bone Eaters? Arthritis Rheumatol. 2016, 68, 2834–2836. [Google Scholar] [CrossRef] [PubMed]
- Pettit, A.R.; Ji, H.; Von Stechow, D.; Muller, R.; Goldring, S.R.; Choi, Y.; Benoist, C.; Gravallese, E.M. TRANCE/RANKL knockout mice are protected from bone erosion in a serum transfer model of arthritis. Am. J. Pathol. 2001, 159, 1689–1699. [Google Scholar] [CrossRef]
- Parchim, N.F.; Wang, W.; Iriyama, T.; Ashimi, O.A.; Siddiqui, A.H.; Blackwell, S.; Sibai, B.; Kellems, R.E.; Xia, Y. Neurokinin 3 receptor and phosphocholine transferase: Missing factors for pathogenesis of C-reactive protein in preeclampsia. Hypertension 2015, 65, 430–439. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Parchim, N.F.; Wang, W.; Iriyama, T.; Ashimi, O.A.; Liu, C.; Siddiqui, A.H.; Blackwell, S.; Sibai, B.; Kellems, R.E.; Xia, Y. Elevated C-reactive protein contributes to preeclampsia via kinin signaling pathways. In Pregnancy Hypertension: An International Journal of Women’s Cardiovascular Health; Elsevier: Amsterdam, The Netherlands, 2015; Volume 5, p. 87. [Google Scholar]
- Metgud, R.; Bajaj, S. Altered serum and salivary C-reactive protein levels in patients with oral premalignant lesions and oral squamous cell carcinoma. Biotech. Histochem. 2016, 91, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Tai, S.F.; Chien, H.T.; Young, C.K.; Tsao, C.K.; De Pablo, A.; Fan, K.H.; Liao, C.T.; Wang, H.M.; Kang, C.J.; Chang, J.T.; et al. Roles of preoperative C-reactive protein are more relevant in buccal cancer than other subsites. World J. Surg. Oncol. 2017, 15, 47. [Google Scholar] [CrossRef] [PubMed]
- Khandavilli, S.D.; Ceallaigh, P.O.; Lloyd, C.J.; Whitaker, R. Serum C-reactive protein as a prognostic indicator in patients with oral squamous cell carcinoma. Oral Oncol. 2009, 45, 912–914. [Google Scholar] [CrossRef]
- Blatt, S.; Schon, H.; Sagheb, K.; Kammerer, P.W.; Al-Nawas, B.; Schiegnitz, E. Hemoglobin, C-reactive protein and ferritin in patients with oral carcinoma and their clinical significance—A prospective clinical study. J. Craniomaxillofac. Surg. 2018, 46, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Tampa, M.; Mitran, M.I.; Mitran, C.I.; Sarbu, M.I.; Matei, C.; Nicolae, I.; Caruntu, A.; Tocut, S.M.; Popa, M.I.; Caruntu, C.; et al. Mediators of Inflammation—A Potential Source of Biomarkers in Oral Squamous Cell Carcinoma. J. Immunol. Res. 2018, 2018. [Google Scholar] [CrossRef]
- Li, F.X.; Xu, F.; Lin, X.; Wu, F.; Zhong, J.Y.; Wang, Y.; Guo, B.; Zheng, M.H.; Shan, S.K.; Yuan, L.Q. The Role of Substance P in the Regulation of Bone and Cartilage Metabolic Activity. Front. Endocrinol. 2020, 11, 77. [Google Scholar] [CrossRef] [PubMed]
- Tatullo, M.; Spagnuolo, G.; Codispoti, B.; Zamparini, F.; Zhang, A.; Degli Esposti, M.; Aparicio, C.; Rengo, C.; Nuzzolese, M.; Manzoli, L.; et al. PLA-Based Mineral-Doped Scaffolds Seeded with Human Periapical Cyst-Derived MSCs: A Promising Tool for Regenerative Healing in Dentistry. Materials 2019, 12, 597. [Google Scholar] [CrossRef] [PubMed]
- Marrelli, M.; Codispoti, B.; Shelton, R.M.; Scheven, B.A.; Cooper, P.R.; Tatullo, M.; Paduano, F. Dental Pulp Stem Cell Mechanoresponsiveness: Effects of Mechanical Stimuli on Dental Pulp Stem Cell Behavior. Front. Physiol. 2018, 9, 1685. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Number (N = 27) | % | |
|---|---|---|---|
| Age (years) | 39–89 (mean 72.8) | ||
| Gender | Male | 10 | 37.0 |
| Female | 17 | 63.0 | |
| Tumor site | Upper gingiva | 10 | 37.0 |
| Lower gingiva | 17 | 63.0 | |
| Tumor size | <2 cm | 4 | 14.8 |
| ≥2 cm, <4 cm | 17 | 63.0 | |
| ≥4 cm | 6 | 22.2 | |
| Pathological N stage | N0 | 22 | 81.5 |
| N1 | 3 | 11.1 | |
| N2 | 2 | 7.4 | |
| Pathological TNM stage | I | 3 | 11.1 |
| II | 13 | 48.1 | |
| III | 1 | 3.7 | |
| IVA | 9 | 33.3 | |
| IVB | 1 | 3.7 | |
| Differentiation | Well | 15 | 55.6 |
| Moderate | 9 | 33.3 | |
| Poor | 3 | 11.1 |
| Parameter | NK-3R Expression in Tumor Cells | p-Value | ||
|---|---|---|---|---|
| Positive (N = 23) | Negative (N = 4) | |||
| Age | <70 | 7 | 2 | 0.41 |
| ≥70 | 16 | 2 | ||
| Gender | Male | 7 | 3 | 0.13 |
| Female | 16 | 1 | ||
| Tumor size | <2 cm | 2 | 2 | 0.08 |
| ≥2 cm, <4 cm | 16 | 1 | ||
| ≥4 cm | 5 | 1 | ||
| N stage | N0 | 18 | 4 | 0.59 |
| N1 | 3 | 0 | ||
| N2 | 2 | 0 | ||
| TNM Stage | I, II | 13 | 3 | 0.49 |
| III, IV | 10 | 1 | ||
| Differentiation | Well | 11 | 4 | 0.08 |
| Moderate–poor | 12 | 0 | ||
| Bone invasion | No bone invasion | 7 | 4 | 0.03 |
| Cortical | 8 | 0 | ||
| Medullary | 8 | 0 | ||
| Lymphovascular invasion | Absent | 19 | 4 | 0.37 |
| Present | 4 | 0 | ||
| Perineural invasion | Absent | 20 | 4 | 0.44 |
| Present | 3 | 0 | ||
| Primary recurrence | Absent | 17 | 3 | 0.96 |
| Present | 6 | 1 | ||
| Secondary cervical lymph nodal metastasis | Absent | 20 | 4 | 0.61 |
| Present | 3 | 0 | ||
| Parameter | ‘NK-3R-Positive’ Osteoclast Number | p-Value | ||
|---|---|---|---|---|
| Large (N = 9) | Small (N = 16) | |||
| Age | <70 | 2 | 7 | 0.28 |
| ≥70 | 7 | 9 | ||
| Gender | Male | 1 | 7 | 0.09 |
| Female | 8 | 9 | ||
| Tumor size | <2 cm | 2 | 2 | 0.49 |
| ≥2 cm, <4 cm | 4 | 11 | ||
| ≥4 cm | 3 | 3 | ||
| N stage | N0 | 6 | 15 | 0.12 |
| N1 | 1 | 1 | ||
| N2 | 2 | 0 | ||
| TNM Stage | I, II | 3 | 13 | 0.016 |
| III, IV | 6 | 3 | ||
| Differentiation | Well | 5 | 9 | 0.97 |
| Moderate–poor | 4 | 7 | ||
| Bone invasion | No bone invasion | 0 | 11 | <0.001 |
| Cortical | 3 | 5 | ||
| Medullary | 6 | 0 | ||
| Lymphovascular invasion | Absent | 6 | 15 | 0.07 |
| Present | 3 | 1 | ||
| Perineural invasion | Absent | 7 | 15 | 0.23 |
| Present | 2 | 1 | ||
| Primary recurrence | Absent | 6 | 12 | 0.71 |
| Present | 3 | 4 | ||
| Secondary cervical lymph nodal metastasis | Absent | 8 | 14 | 0.91 |
| Present | 1 | 2 | ||
| NK-3R expression in tumor cells | Positive | 9 | 12 | 0.1 |
| Negative | 0 | 4 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshida, S.; Shimo, T.; Takabatake, K.; Murase, Y.; Obata, K.; Okui, T.; Kunisada, Y.; Ibaragi, S.; Nagatsuka, H.; Sasaki, A. Expression of Neurokinin B Receptor in the Gingival Squamous Cell Carcinoma Bone Microenvironment. Diagnostics 2021, 11, 1044. https://doi.org/10.3390/diagnostics11061044
Yoshida S, Shimo T, Takabatake K, Murase Y, Obata K, Okui T, Kunisada Y, Ibaragi S, Nagatsuka H, Sasaki A. Expression of Neurokinin B Receptor in the Gingival Squamous Cell Carcinoma Bone Microenvironment. Diagnostics. 2021; 11(6):1044. https://doi.org/10.3390/diagnostics11061044
Chicago/Turabian StyleYoshida, Shoko, Tsuyoshi Shimo, Kiyofumi Takabatake, Yurika Murase, Kyoichi Obata, Tatsuo Okui, Yuki Kunisada, Soichiro Ibaragi, Hitoshi Nagatsuka, and Akira Sasaki. 2021. "Expression of Neurokinin B Receptor in the Gingival Squamous Cell Carcinoma Bone Microenvironment" Diagnostics 11, no. 6: 1044. https://doi.org/10.3390/diagnostics11061044
APA StyleYoshida, S., Shimo, T., Takabatake, K., Murase, Y., Obata, K., Okui, T., Kunisada, Y., Ibaragi, S., Nagatsuka, H., & Sasaki, A. (2021). Expression of Neurokinin B Receptor in the Gingival Squamous Cell Carcinoma Bone Microenvironment. Diagnostics, 11(6), 1044. https://doi.org/10.3390/diagnostics11061044