Dietary Management of Obesity: A Review of the Evidence


Abstract
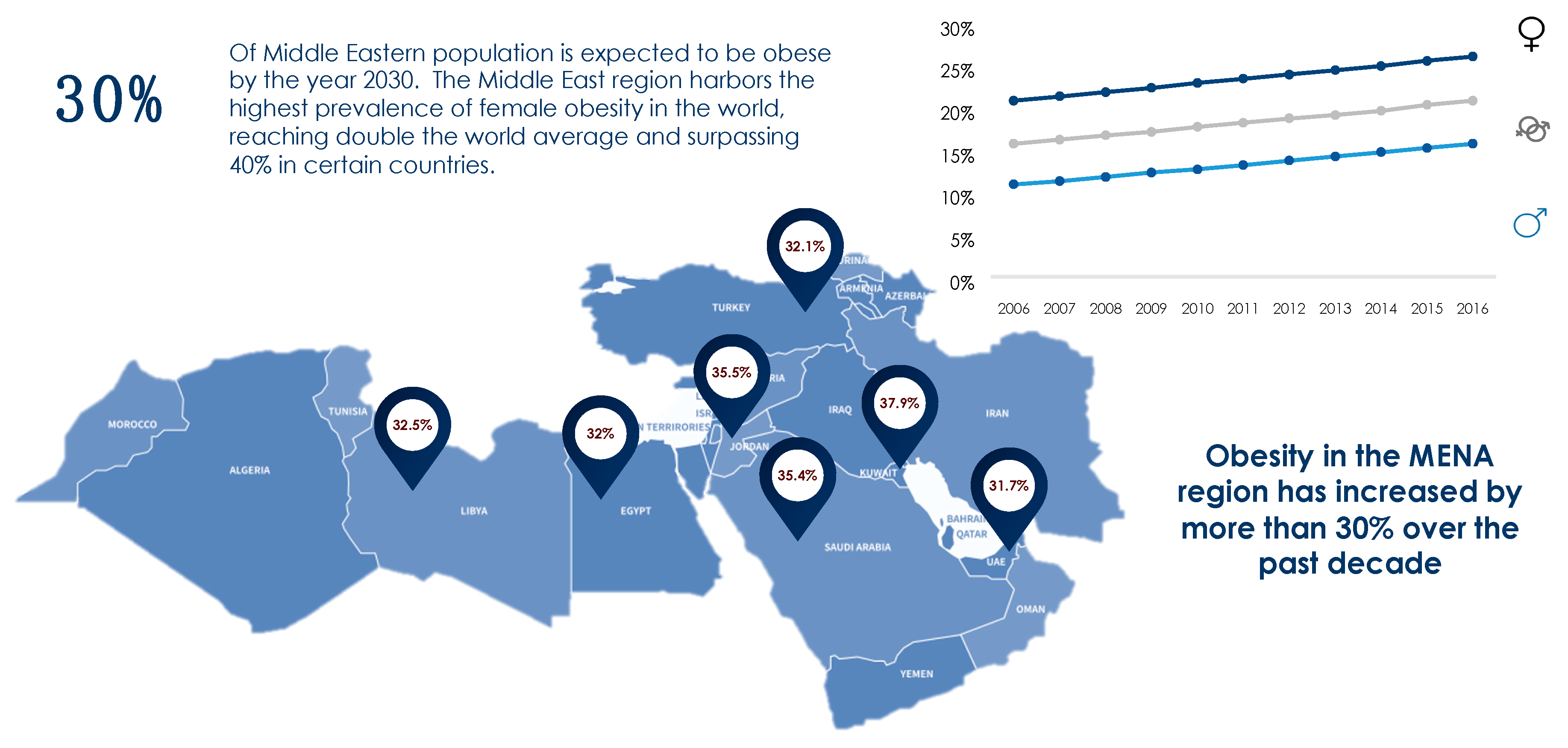
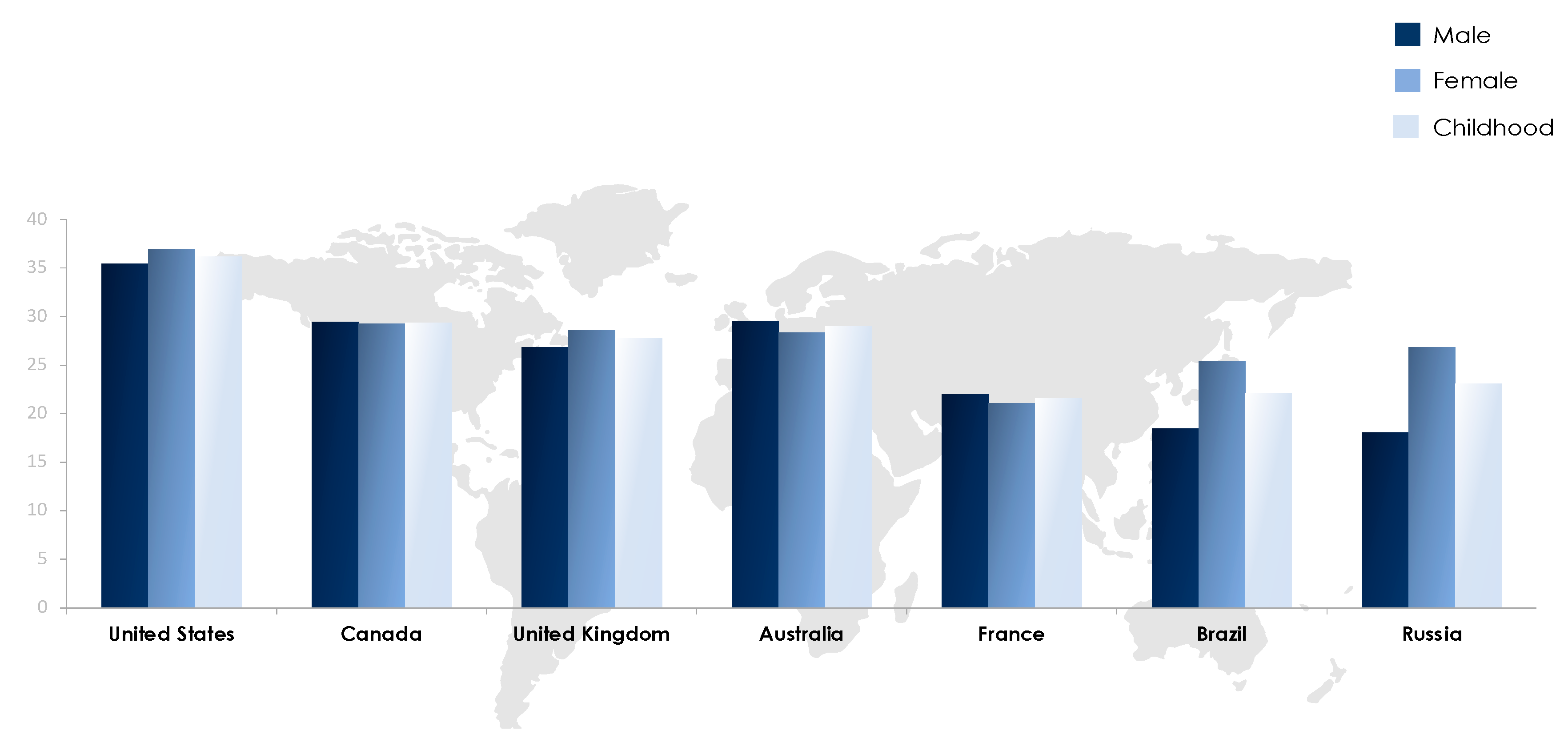
1. Obesity: Prevalence and Magnitude
2. Population-Wide Initiatives to Combat Obesity
3. The Interplay of External and Internal Culprits
3.1. Role of Gene-Environment Interaction
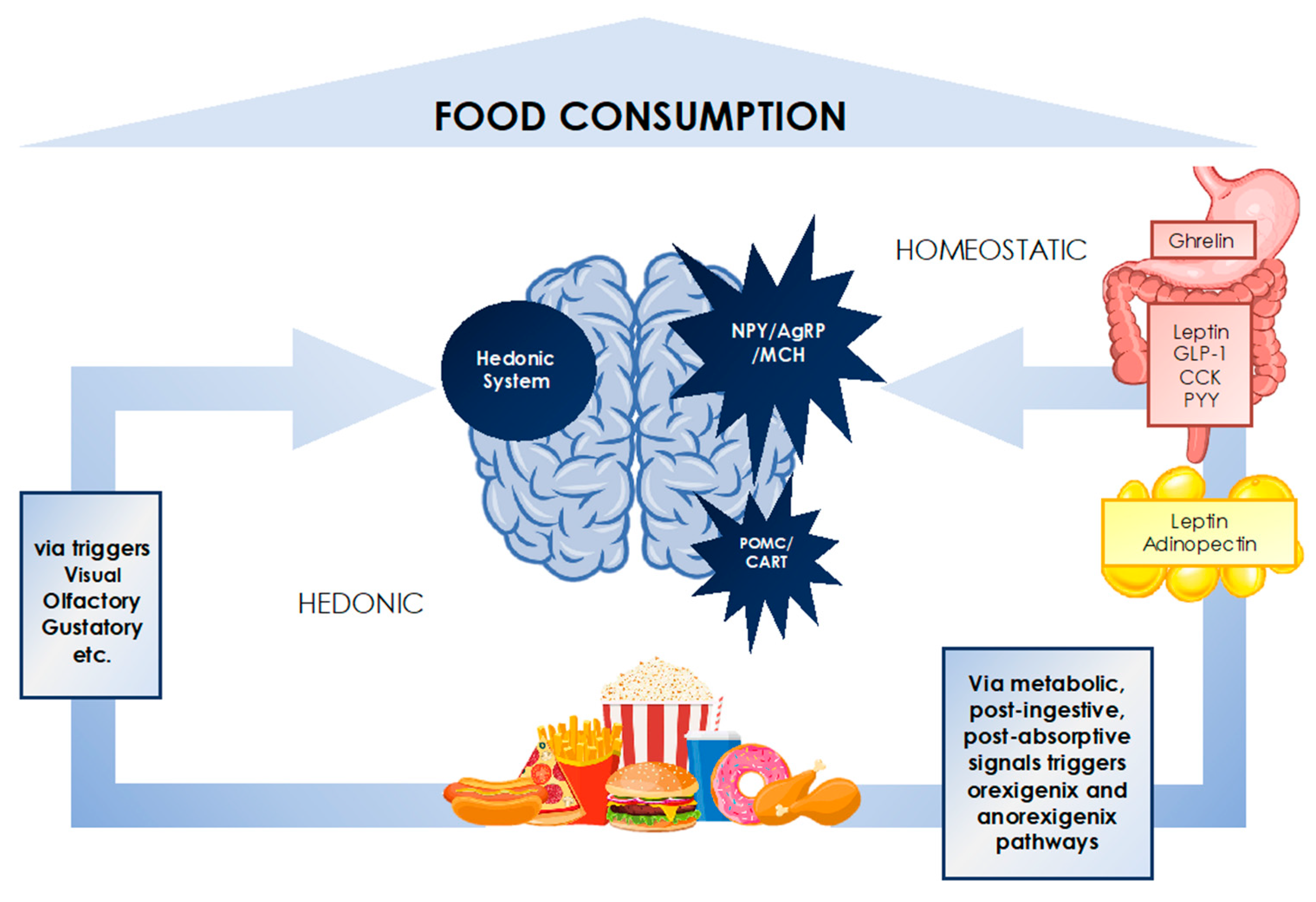
3.2. Regulation of Food Intake—Homeostatic and Hedonic Systems
4. Dietary Interventions in the Management of Obesity
4.1. Macronutrient-Focused Approach
4.1.1. Effects on Weight Loss
4.1.2. Effect of Metabolic Regulation and Adherence
High Protein Diets
Low Carbohydrate Diets
Low Fat Diets
4.2. Dietary Pattern-Focused Approach (The Mediterranean Diet)
4.3. Dietary Timing-Focused Approach (Intermittent Fasting)
5. The Epigenetic Effects of Diet
6. The Way Forward
Author Contributions
Funding
Conflicts of Interest
References
- Null, N. Trends in adult body-mass index in 200 countries from 1975 to 2014: A pooled analysis of 1698 population-based measurement studies with 19·2 million participants. Lancet 2016, 387, 1377–1396. [Google Scholar]
- Global Health Observatory. Prevalence of Obesity among Adults, BMI ≥ 30, Age-Standardized—Estimates by WHO Region [Online]. 2016. Available online: https://apps.who.int/gho/data/view.main.REGION2480A?lang=en (accessed on 19 October 2020).
- International Diabetes Federation. IDF Diabetes Atlas Ninth Edition 2019; International Diabetes Federation: Brussels, Belgium, 2019. [Google Scholar]
- McKinsey Global Institute. Overcoming Obesity: An Initial Economic Analysis; McKinsey & Company: London, UK, 2014; Available online: https://www.mckinsey.com/~/media/mckinsey/business%20functions/economic%20studies%20temp/our%20insights/how%20the%20world%20could%20better%20fight%20obesity/mgi_overcoming_obesity_full_report.ashx (accessed on 19 October 2020).
- Gortmaker, S.L.; Wang, Y.C.; Long, M.W.; Giles, C.M.; Ward, Z.J.; Barrett, J.L.; Kenney, E.L.; Sonneville, K.R.; Afzal, A.S.; Resch, S.C.; et al. Three Interventions That Reduce Childhood Obesity Are Projected To Save More Than They Cost To Implement. Health Aff. 2015, 34, 1932–1939. [Google Scholar] [CrossRef] [PubMed]
- Lavie, C.J.; Laddu, D.; Arena, R.; Ortega, F.B.; Alpert, M.A.; Kushner, R.F. Healthy weight and obesity prevention: JACC health promotion series. J. Am. Coll. Cardiol. 2018, 72, 1506–1531. [Google Scholar] [CrossRef] [PubMed]
- Sisnowski, J.; Street, J.; Merlin, T. Improving food environments and tackling obesity: A realist systematic review of the policy success of regulatory interventions targeting population nutrition. PLoS ONE 2017, 12, e0182581. [Google Scholar] [CrossRef]
- Barquera, S.; Campos, I.; Rivera, J.A. Mexico attempts to tackle obesity: The process, results, push backs and future challenges. Obes. Rev. 2013, 14, 69–78. [Google Scholar] [CrossRef]
- Colchero, M.A.; Popkin, B.M.; Rivera, J.A.; Ng, S.W. Beverage purchases from stores in Mexico under the excise tax on sugar sweetened beverages: Observational study. BMJ 2016, 352, h6704. [Google Scholar] [CrossRef] [PubMed]
- Batis, C.; Rivera, J.A.; Popkin, B.M.; Taillie, L.S. First-Year Evaluation of Mexico’s Tax on Nonessential Energy-Dense Foods: An Observational Study. PLoS Med. 2016, 13, e1002057. [Google Scholar] [CrossRef]
- Colchero, M.A.; Rivera-Dommarco, J.; Popkin, B.M.; Ng, S.W. In Mexico, Evidence Of Sustained Consumer Response Two Years After Implementing A Sugar-Sweetened Beverage Tax. Health Aff. 2017, 36, 564–571. [Google Scholar] [CrossRef]
- Fernandez, M.A.; Raine, K.D. Insights on the Influence of Sugar Taxes on Obesity Prevention Efforts. Curr. Nutr. Rep. 2019, 8, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Osiac, L.; Cofré, C.; Pizarro, T.; Mansilla, C.; Herrera, C.A.; Burrows, J.; Castillo, C. Using evidence-informed policies to tackle overweight and obesity in Chile. Rev. Panam. Salud Pública 2017, 41, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Foster, N.; Thow, A.M.; Unwin, N.; Alvarado, M.; Samuels, T.A. Regulatory measures to fight obesity in Small Island Developing States of the Caribbean and Pacific, 2015–2017. Rev. Panam. Salud Pública 2018, 42, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Thow, A.M.; Downs, S.M.; Mayes, C.; Trevena, H.; Waqanivalu, T.; Cawley, J. Fiscal policy to improve diets and prevent noncommunicable diseases: From recommendations to action. Bull. World Health Organ. 2018, 96, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Samara, A.; Andersen, P.T.; Aro, A.R. Health Promotion and Obesity in the Arab Gulf States: Challenges and Good Practices. J. Obes. 2019, 2019, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Caballero, B. The Global Epidemic of Obesity: An Overview. Epidemiol. Rev. 2007, 29, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Heymsfield, S.B.; Wadden, T.A. Mechanisms, Pathophysiology, and Management of Obesity. N. Engl. J. Med. 2017, 376, 254–266. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Cardel, M.; Donahoo, W.T. Social and Environmental Factors Influencing Obesity. In Endotext [Internet]; MDText. com, Inc.: South Dartmouth, MA, USA, 2019. [Google Scholar]
- Schwartz, M.B.; Just, D.R.; Chriqui, J.F.; Ammerman, A.S. Appetite self-regulation: Environmental and policy influences on eating behaviors. Obesity 2017, 25, S26–S38. [Google Scholar] [CrossRef]
- Whelan, K. Advanced Nutrition and Dietetics in Obesity; John Wiley & Sons: Hoboken, NJ, USA, 2018. [Google Scholar]
- Van Djik, S.; Tellam, R.L.; Morrison, J.L.; Mühlhäusler, B.S.; Molloy, P. Recent developments on the role of epigenetics in obesity and metabolic disease. Clin. Epigenetics 2015, 7, 1–13. [Google Scholar] [CrossRef]
- Gkouskou, K.; Vlastos, I.; Karkalousos, P.; Chaniotis, D.; Sanoudou, D.; Eliopoulos, A.G. The “Virtual Digital Twins” Concept in Precision Nutrition. Adv. Nutr. 2020, 11, 1405–1413. [Google Scholar] [CrossRef]
- Abdul, Q.A.; Yu, B.P.; Chung, H.Y.; Jung, H.A.; Choi, J.S. Epigenetic modifications of gene expression by lifestyle and environment. Arch. Pharmacal Res. 2017, 40, 1219–1237. [Google Scholar] [CrossRef]
- Sulc, J.A.; Mounier, N.; Günther, F.; Winkler, T.W.; Wood, A.R.; Frayling, T.M.; Heid, I.M.; Robinson, M.R.; Kutalik, Z. Quantification of the overall contribution of gene-environment interaction for obesity-related traits. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Singh, A.K.; Bishayee, A.; Pandey, A.K. Targeting Histone Deacetylases with Natural and Synthetic Agents: An Emerging Anticancer Strategy. Nutrients 2018, 10, 731. [Google Scholar] [CrossRef] [PubMed]
- Hall, E.; Volkov, P.; Dayeh, T.; Bacos, K.; Rönn, T.; Nitert, M.D.; Ling, C. Effects of palmitate on genome-wide mRNA expression and DNA methylation patterns in human pancreatic islets. BMC Med. 2014, 12, 103. [Google Scholar] [CrossRef] [PubMed]
- González-Becerra, K.; Ramos-Lopez, O.; Barrón-Cabrera, E.; Riezu-Boj, J.I.; Milagro, F.I.; Martinez-Lopez, E.; Martínez, J.A. Fatty acids, epigenetic mechanisms and chronic diseases: A systematic review. Lipids Health Dis. 2019, 18, 178. [Google Scholar] [CrossRef] [PubMed]
- Bilman, E.; Van Kleef, E.; Van Trijp, H. External cues challenging the internal appetite control system—Overview and practical implications. Crit. Rev. Food Sci. Nutr. 2017, 57, 2825–2834. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, M.; Blundell, J.; Halford, J.; King, N.; Finlayson, G. The Regulation of Food Intake in Humans. In Endotext [Internet]; MDText. com, Inc.: South Dartmouth, MA, USA, 2015. [Google Scholar]
- Moehlecke, M.; Canani, L.H.; Silva, L.O.J.E.; Trindade, M.R.M.; Friedman, R.; Leitão, C.B. Determinants of body weight regulation in humans. Arch. Endocrinol. Metab. 2016, 60, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Sumithran, P.; Prendergast, L.A.; Delbridge, E.; Purcell, K.; Shulkes, A.; Kriketos, A.; Proietto, J. Long-Term Persistence of Hormonal Adaptations to Weight Loss. N. Engl. J. Med. 2011, 365, 1597–1604. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Singh, S.N. Regulation of Food Intake: A Complex Process. Def. Life Sci. J. 2018, 3, 182–189. [Google Scholar] [CrossRef]
- De Silva, A.; Bloom, S.R. Gut Hormones and Appetite Control: A Focus on PYY and GLP-1 as Therapeutic Targets in Obesity. Gut Liver 2012, 6, 10–20. [Google Scholar] [CrossRef]
- Yu, Y.; Vasselli, J.R.; Zhang, Y.; Mechanick, J.I.; Korner, J.; Peterli, R. Metabolic vs. hedonic obesity: A conceptual distinction and its clinical implications. Obes. Rev. 2015, 16, 234–247. [Google Scholar] [CrossRef]
- Lee, P.C.; Dixon, J.B. Food for Thought: Reward Mechanisms and Hedonic Overeating in Obesity. Curr. Obes. Rep. 2017, 6, 353–361. [Google Scholar] [CrossRef]
- Finlayson, G. Food addiction and obesity: Unnecessary medicalization of hedonic overeating. Nat. Rev. Endocrinol. 2017, 13, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, N.S.; Hinton, E.; Parkinson, J.; Lawrence, A.D. Nucleus accumbens response to food cues predicts subsequent snack consumption in women and increased body mass index in those with reduced self-control. NeuroImage 2012, 63, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Paolini, B.M.; Laurienti, P.J.; Simpson, S.L.; Burdette, J.H.; Lyday, R.G.; Erejeski, W.J. Global integration of the hot-state brain network of appetite predicts short term weight loss in older adult. Front. Aging Neurosci. 2015, 7, 70. [Google Scholar] [CrossRef] [PubMed]
- Murdaugh, D.L.; Cox, J.E.; Cook, E.W., III; Weller, R.E. fMRI reactivity to high-calorie food pictures predicts short-and long-term outcome in a weight-loss program. Neuroimage 2012, 59, 2709–2721. [Google Scholar] [CrossRef] [PubMed]
- Hall, K.D.; Hammond, R.A.; Rahmandad, H. Dynamic Interplay among Homeostatic, Hedonic, and Cognitive Feedback Circuits Regulating Body Weight. Am. J. Public Health 2014, 104, 1169–1175. [Google Scholar] [CrossRef]
- Koliaki, C.; Spinos, T.; Spinou, M.; Brinia, M.-E.; Mitsopoulou, D.; Katsilambros, N. Defining the Optimal Dietary Approach for Safe, Effective and Sustainable Weight Loss in Overweight and Obese Adults. Healthcare 2018, 6, 73. [Google Scholar] [CrossRef]
- Katsilambros, N.L. Nutritional treatment of obesity in adults. What is certain and what uncertain. Arch. Hell. Med./Arheia Ellenikes Iatrikes 2015, 32. [Google Scholar]
- Langeveld, M.; Devries, J.H. The long-term effect of energy restricted diets for treating obesity. Obesity 2015, 23, 1529–1538. [Google Scholar] [CrossRef]
- Anderson, J.W.; Konz, E.C.; Frederich, R.C.; Wood, C.L. Long-term weight-loss maintenance: A meta-analysis of US studies. Am. J. Clin. Nutr. 2001, 74, 579–584. [Google Scholar] [CrossRef]
- Dayan, P.H.; Sforzo, G.; Boisseau, N.; Pereira-Lancha, L.O.; Lancha, A.H., Jr. A new clinical perspective: Treating obesity with nutritional coaching versus energy-restricted diets. Nutrition 2019, 60, 147–151. [Google Scholar] [CrossRef]
- Lam, C.C.B. Obesity Updates: Understanding Obesity as a Disease and Intermittent Fasting. Singap. Fam. Physicians 2019, 10, 15–18. [Google Scholar] [CrossRef]
- Bray, G.A.; Krauss, R.M.; Sacks, F.M.; Qi, L. Lessons Learned from the POUNDS Lost Study: Genetic, Metabolic, and Behavioral Factors Affecting Changes in Body Weight, Body Composition, and Cardiometabolic Risk. Curr. Obes. Rep. 2019, 8, 262–283. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.; Abdelhamid, A.; Moore, H.J.; Douthwaite, W.; Skeaff, C.M.; Summerbell, C.D. Effect of reducing total fat intake on body weight: Systematic review and meta-analysis of randomised controlled trials and cohort studies. BMJ 2012, 345, e7666. [Google Scholar] [CrossRef] [PubMed]
- Wycherley, T.P.; Moran, L.J.; Clifton, P.M.; Noakes, M.; Brinkworth, G.D. Effects of energy-restricted high-protein, low-fat compared with standard-protein, low-fat diets: A meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2012, 96, 1281–1298. [Google Scholar] [CrossRef]
- Ajala, O.; English, P.; Pinkney, J. Systematic review and meta-analysis of different dietary approaches to the management of type 2 diabetes. Am. J. Clin. Nutr. 2013, 97, 505–516. [Google Scholar] [CrossRef]
- Bueno, N.B.; de Melo, I.S.V.; de Oliveira, S.L.; Ataide, T.D.R. Very-low-carbohydrate ketogenic diet v. low-fat diet for long-term weight loss: A meta-analysis of randomised controlled trials. Br. J. Nutr. 2013, 110, 1178–1187. [Google Scholar] [CrossRef]
- Naude, C.E.; Schoonees, A.; Senekal, M.; Young, T.; Garner, P.; Volmink, J. Low Carbohydrate versus Isoenergetic Balanced Diets for Reducing Weight and Cardiovascular Risk: A Systematic Review and Meta-Analysis. PLoS ONE 2014, 9, e100652. [Google Scholar] [CrossRef]
- Clifton, P.M.; Condo, D.; Keogh, J. Long term weight maintenance after advice to consume low carbohydrate, higher protein diets—A systematic review and meta analysis. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 224–235. [Google Scholar] [CrossRef]
- Tobias, D.K.; Chen, M.; Manson, J.E.; Ludwig, D.S.; Willett, W.C.; Hu, F.B. Effect of low-fat diet interventions versus other diet interventions on long-term weight change in adults: A systematic review and meta-analysis. Lancet Diabetes Endocrinol. 2015, 3, 968–979. [Google Scholar] [CrossRef]
- Boaz, O.R.M.; Raz, O. Low Fat vs. Low Carbohydrate Diet Strategies for Weight Reduction: A Meta-Analysis. J. Obes. Weight. Loss Ther. 2015, 5, 2. [Google Scholar] [CrossRef]
- Mansoor, N.; Vinknes, K.J.; Veierød, M.B.; Retterstol, K. Effects of low-carbohydrate dietsv. low-fat diets on body weight and cardiovascular risk factors: A meta-analysis of randomised controlled trials. Br. J. Nutr. 2016, 115, 466–479. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Sadeghirad, B.; Ball, G.D.C.; Da Costa, B.R.; Hitchcock, C.L.; Svendrovski, A.; Kiflen, R.; Quadri, K.; Kwon, H.Y.; Karamouzian, M.; et al. Comparison of dietary macronutrient patterns of 14 popular named dietary programmes for weight and cardiovascular risk factor reduction in adults: Systematic review and network meta-analysis of randomised trials. BMJ 2020, 369, m696. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Qi, Q.; Zhang, C.; Smith, S.R.; Hu, F.B.; Sacks, F.M.; Bray, G.A.; Qi, L. FTO Genotype and 2-Year Change in Body Composition and Fat Distribution in Response to Weight-Loss Diets: The POUNDS LOST Trial. Diabetes 2012, 61, 3005–3011. [Google Scholar] [CrossRef] [PubMed]
- Carreiro, A.; Dhillon, J.; Gordon, S.; Higgins, K.A.; Jacobs, A.G.; McArthur, B.M.; Redan, B.W.; Rivera, R.L.; Schmidt, L.R.; Mattes, R.D. The Macronutrients, Appetite, and Energy Intake. Annu. Rev. Nutr. 2016, 36, 73–103. [Google Scholar] [CrossRef]
- Vogels, N.; Diepvens, K.; Westerterp-Plantenga, M.S. Predictors of Long-term Weight Maintenance. Obes. Res. 2005, 13, 2162–2168. [Google Scholar] [CrossRef]
- Drummen, M.; Tischmann, L.; Gatta-Cherifi, B.; Adam, T.; Westerterp-Plantenga, M. Dietary Protein and Energy Balance in Relation to Obesity and Co-morbidities. Front. Endocrinol. 2018, 9, 443. [Google Scholar] [CrossRef]
- Morell, P.; Fiszman, S. Revisiting the role of protein-induced satiation and satiety. Food Hydrocoll. 2017, 68, 199–210. [Google Scholar] [CrossRef]
- Pesta, D.H.; Samuel, V.T. A high-protein diet for reducing body fat: Mechanisms and possible caveats. Nutr. Metab. 2014, 11, 1–8. [Google Scholar] [CrossRef]
- Batterham, R.L.; Heffron, H.; Kapoor, S.; Chivers, J.E.; Chandarana, K.; Herzog, H.; Le Roux, C.W.; Thomas, E.L.; Bell, J.D.; Withers, D.J. Critical role for peptide YY in protein-mediated satiation and body-weight regulation. Cell Metab. 2006, 4, 223–233. [Google Scholar] [CrossRef]
- Hall, W.L.; Millward, D.J.; Long, S.J.; Morgan, L.M. Casein and whey exert different effects on plasma amino acid profiles, gastrointestinal hormone secretion and appetite. Br. J. Nutr. 2003, 89, 239–248. [Google Scholar] [CrossRef]
- Lejeune, M.; Westerterp, K.R.; Adam, T.C.M.; Luscombe-Marsh, N.D.; Westerterp-Plantenga, M.S. Ghrelin and glucagon-like peptide 1 concentrations, 24-h satiety, and energy and substrate metabolism during a high-protein diet and measured in a respiration chamber. Am. J. Clin. Nutr. 2006, 83, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Smeets, A.J.; Soenen, S.; Luscombe-Marsh, N.D.; Ueland, Ø.; Westerterp-Plantenga, M.S. Energy Expenditure, Satiety, and Plasma Ghrelin, Glucagon-Like Peptide 1, and Peptide Tyrosine-Tyrosine Concentrations following a Single High-Protein Lunch. J. Nutr. 2008, 138, 698–702. [Google Scholar] [CrossRef] [PubMed]
- Veldhorst, M.A.; Nieuwenhuizen, A.G.; Hochstenbach-Waelen, A.; Westerterp, K.R.; Engelen, M.P.K.J.; Brummer, R.-J.M.; Deutz, N.E.P.; Westerterp-Plantenga, M.S. Comparison of the effects of a high- and normal-casein breakfast on satiety, ‘satiety’ hormones, plasma amino acids and subsequent energy intake. Br. J. Nutr. 2009, 101, 295–303. [Google Scholar] [CrossRef]
- Tomé, D.; Chaumontet, C.; Even, P.C.; Darcel, N.; Azzout-Marniche, D. Protein status modulates the rewarding value of foods and meals to maintain an adequate protein intake. Physiol. Behav. 2019, 206, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Vu, J.P.; Luong, L.; Parsons, W.F.; Oh, S.; Sanford, D.; Gabalski, A.; Lighton, J.R.; Pisegna, J.R.; Germano, P. Long-term intake of a high-protein diet affects body phenotype, metabolism, and plasma hormones in mice. J. Nutr. 2017, 147, 2243–2251. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, P.B.; Toubro, S.; Astrup, A. Effect of fat-reduced diets on 24-h energy expenditure: Comparisons between animal protein, vegetable protein, and carbohydrate. Am. J. Clin. Nutr. 2000, 72, 1135–1141. [Google Scholar] [CrossRef] [PubMed]
- Azzout-Marniche, D.; Gaudichon, C.; Blouet, C.; Bos, C.; Mathé, V.; Huneau, J.-F.; Tomé, D. Liver glyconeogenesis: A pathway to cope with postprandial amino acid excess in high-protein fed rats? Am. J. Physiol. Integr. Comp. Physiol. 2007, 292, R1400–R1407. [Google Scholar] [CrossRef] [PubMed]
- Campos-Nonato, I.; Hernandez, L.; Barquera, S. Effect of a High-Protein Diet versus Standard-Protein Diet on Weight Loss and Biomarkers of Metabolic Syndrome: A Randomized Clinical Trial. Obes. Facts 2017, 10, 238–251. [Google Scholar] [CrossRef]
- Verreijen, A.M.; Engberink, M.F.; Memelink, R.G.; Van Der Plas, S.E.; Visser, M.; Weijs, P.J.M. Effect of a high protein diet and/or resistance exercise on the preservation of fat free mass during weight loss in overweight and obese older adults: A randomized controlled trial. Nutr. J. 2017, 16, 1–8. [Google Scholar] [CrossRef]
- Noakes, M.; Keogh, J.B.; Foster, P.R.; Clifton, P.M. Effect of an energy-restricted, high-protein, low-fat diet relative to a conventional high-carbohydrate, low-fat diet on weight loss, body composition, nutritional status, and markers of cardiovascular health in obese women. Am. J. Clin. Nutr. 2005, 81, 1298–1306. [Google Scholar] [CrossRef]
- Johnstone, A.M.; Horgan, G.W.; Murison, S.D.; Bremner, D.M.; Lobley, G.E. Effects of a high-protein ketogenic diet on hunger, appetite, and weight loss in obese men feeding ad libitum. Am. J. Clin. Nutr. 2008, 87, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, A.M.; Lobley, G.E.; Horgan, G.W.; Bremner, D.M.; Fyfe, C.L.; Morrice, P.C.; Duthie, G.G. Effects of a high-protein, low-carbohydrate v. high-protein, moderate-carbohydrate weight-loss diet on antioxidant status, endothelial markers and plasma indices of the cardiometabolic profile. Br. J. Nutr. 2011, 106, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Hwalla, N.C.; Sawaya, S.; Torbay, N.; Habbal, Z.; Azar, S.; Hashim, S.A.; Habbal, M.Z. High protein vs high carbohydrate hypoenergetic diet for the treatment of obese hyperinsulinemic subjects. Int. J. Obes. 1999, 23, 1202–1206. [Google Scholar] [CrossRef] [PubMed]
- Gibson, A.A.; Seimon, R.V.; Lee, C.M.Y.; Ayre, J.; Franklin, J.; Markovic, T.P.; Caterson, I.D.; Sainsbury, A. Do ketogenic diets really suppress appetite? A systematic review and meta-analysis. Obes. Rev. 2014, 16, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Sumithran, P.; Prendergast, L.A.; Delbridge, E.; Purcell, K.; Shulkes, A.; Kriketos, A.D.; Proietto, J. Ketosis and appetite-mediating nutrients and hormones after weight loss. Eur. J. Clin. Nutr. 2013, 67, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Paoli, A.; Bosco, G.; Camporesi, E.M.; Mangar, D. Ketosis, ketogenic diet and food intake control: A complex relationship. Front. Psychol. 2015, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Deemer, S.E.; Plaisance, E.P.; Martins, C. Impact of ketosis on appetite regulation—A review. Nutr. Res. 2020, 77, 1–11. [Google Scholar] [CrossRef]
- Jaillard, T.; Roger, M.; Galinier, A.; Guillou, P.; Benani, A.; Leloup, C.; Casteilla, L.; Pénicaud, L.; Lorsignol, A. Hypothalamic Reactive Oxygen Species Are Required for Insulin-Induced Food Intake Inhibition: An NADPH Oxidase–Dependent Mechanism. Diabetes 2009, 58, 1544–1549. [Google Scholar] [CrossRef]
- Frigolet, M.-E.; Barragán, V.-E.R.; González, M.T. Low-Carbohydrate Diets: A Matter of Love or Hate. Ann. Nutr. Metab. 2011, 58, 320–334. [Google Scholar] [CrossRef]
- Retterstøl, K.; Svendsen, M.; Narverud, I.; Holven, K.B. Effect of low carbohydrate high fat diet on LDL cholesterol and gene expression in normal-weight, young adults: A randomized controlled study. Atherosclerosis 2018, 279, 52–61. [Google Scholar] [CrossRef]
- Brouns, F. Overweight and diabetes prevention: Is a low-carbohydrate–high-fat diet recommendable? Eur. J. Nutr. 2018, 57, 1301–1312. [Google Scholar] [CrossRef]
- Webster, C.C.; Murphy, T.E.; Larmuth, K.M.; Noakes, T.D.O.; Smith, J.A.H. Diet, Diabetes Status, and Personal Experiences of Individuals with Type 2 diabetes Who Self-Selected and Followed a Low Carbohydrate High Fat diet. Diabetes Metab. Syndr. Obes. Targets Ther. 2019, 12, 2567–2582. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.R.; Bellamkonda, S.; Zilbermint, M.; Wang, J.; Kalyani, R.R. Effects of the low carbohydrate, high fat diet on glycemic control and body weight in patients with type 2 diabetes: Experience from a community-based cohort. BMJ Open Diabetes Res. Care 2020, 8, e000980. [Google Scholar] [CrossRef] [PubMed]
- Zong, G.; Gao, A.; Hu, F.B.; Sun, Q. Whole grain intake and mortality from all causes, cardiovascular disease, and cancer: A meta-analysis of prospective cohort studies. Circulation 2016, 133, 2370–2380. [Google Scholar] [CrossRef] [PubMed]
- Brinkworth, G.D.; Noakes, M.; Clifton, P.M.; Bird, A.R. Comparative effects of very low-carbohydrate, high-fat and high-carbohydrate, low-fat weight-loss diets on bowel habit and faecal short-chain fatty acids and bacterial populations. Br. J. Nutr. 2009, 101, 1493–1502. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, E.T.; Bowman, S.A.; Spence, J.T.; Freedman, M.; King, J. Popular diets: Correlation to health, nutrition, and obesity. J. Am. Diet. Assoc. 2001, 101, 411–420. [Google Scholar] [CrossRef]
- Hession, M.; Rolland, C.; Kulkarni, U.; Wise, A.; Broom, J. Systematic review of randomized controlled trials of low-carbohydrate vs. low-fat/low-calorie diets in the management of obesity and its comorbidities. Obes. Rev. 2009, 10, 36–50. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.L.; Esteves, S.S.; da Costa Pereira, A.; Yancy, W.S., Jr.; Nunes, J.P. Systematic review and meta-analysis of clinical trials of the effects of low carbohydrate diets on cardiovascular risk factors. Obes. Rev. 2012, 13, 1048–1066. [Google Scholar] [CrossRef]
- Sackner-Bernstein, J.; Kanter, D.; Kaul, S. Dietary Intervention for Overweight and Obese Adults: Comparison of Low-Carbohydrate and Low-Fat Diets. A Meta-Analysis. PLoS ONE 2015, 10, e0139817. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Fukuda, T.; Oyabu, C.; Tanaka, M.; Asano, M.; Yamazaki, M.; Fukui, M. Impact of low-carbohydrate diet on body composition: Meta-analysis of randomized controlled studies. Obes. Rev. 2016, 17, 499–509. [Google Scholar] [CrossRef]
- Hu, T.; Mills, K.T.; Yao, L.; Demanelis, K.; Eloustaz, M.; Yancy, W.S., Jr.; Kelly, T.N.; He, J.; Bazzano, L.A. Effects of low-carbohydrate diets versus low-fat diets on metabolic risk factors: A meta-analysis of randomized controlled clinical trials. Am. J. Epidemiol. 2012, 176 (Suppl. 7), S44–S54. [Google Scholar] [CrossRef] [PubMed]
- Johnston, B.C.; Kanters, S.; Bandayrel, K.; Wu, P.; Naji, F.; Siemieniuk, R.A.; Ball, G.D.; Busse, J.W.; Thorlund, K.; Guyatt, G.; et al. Comparison of weight loss among named diet programs in overweight and obese adults: A meta-analysis. JAMA 2014, 312, 923–933. [Google Scholar] [CrossRef] [PubMed]
- Hall, K.D.; Chen, K.Y.; Guo, J.; Lam, Y.Y.; Leibel, R.L.; Mayer, L.E.; Reitman, M.L.; Rosenbaum, M.; Smith, S.R.; Walsh, B.T.; et al. Energy expenditure and body composition changes after an isocaloric ketogenic diet in overweight and obese men. Am. J. Clin. Nutr. 2016, 104, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Noto, H.; Goto, A.; Tsujimoto, T.; Noda, M. Low-Carbohydrate Diets and All-Cause Mortality: A Systematic Review and Meta-Analysis of Observational Studies. PLoS ONE 2013, 8, e55030, Erratum in 2019, 14, e0212203. [Google Scholar] [CrossRef] [PubMed]
- Claessens, M.; Van Baak, M.A.; Monsheimer, S.; Saris, W.H.M. The effect of a low-fat, high-protein or high-carbohydrate ad libitum diet on weight loss maintenance and metabolic risk factors. Int. J. Obes. 2009, 33, 296–304. [Google Scholar] [CrossRef]
- Tessitore, A.; Mastroiaco, V.; Vetuschi, A.; Sferra, R.; Pompili, S.; Cicciarelli, G.; Barnabei, R.; Capece, D.; Zazzeroni, F.; Capalbo, C.; et al. Development of hepatocellular cancer induced by long term low fat-high carbohydrate diet in a NAFLD/NASH mouse model. Oncotarget 2017, 8, 53482–53494. [Google Scholar] [CrossRef]
- Hu, T.; Yao, L.; Reynolds, K.; Niu, T.; Li, S.; Whelton, P.K.; He, J.; Bazzano, L.A. The effects of a low-carbohydrate diet on appetite: A randomized controlled trial. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 476–488. [Google Scholar] [CrossRef]
- Boden, G.; Sargrad, K.; Homko, C.; Mozzoli, M.; Stein, T.P. Effect of a Low-Carbohydrate Diet on Appetite, Blood Glucose Levels, and Insulin Resistance in Obese Patients with Type 2 Diabetes. Ann. Intern. Med. 2005, 142, 403–411. [Google Scholar] [CrossRef]
- Essah, P.A.; Levy, J.R.; Sistrun, S.N.; Kelly, S.M.; Nestler, J.E. Effect of Macronutrient Composition on Postprandial Peptide YY Levels. J. Clin. Endocrinol. Metab. 2007, 92, 4052–4055. [Google Scholar] [CrossRef]
- Howard, B.V.; Van Horn, L.; Hsia, J.; Manson, J.E.; Stefanick, M.L.; Wassertheil-Smoller, S.; Kuller, L.H.; LaCroix, A.Z.; Langer, R.D.; Lasser, N.L.; et al. Low-fat dietary pattern and risk of cardiovascular disease: The Women’s Health Initiative Randomized Controlled Dietary Modification Trial. JAMA 2006, 295, 655–666. [Google Scholar] [CrossRef]
- Look AHEAD Research Group. Cardiovascular effects of intensive lifestyle intervention in type 2 diabetes. N. Engl. J. Med. 2013, 369, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.-I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary Prevention of Cardiovascular Disease with a Mediterranean Diet Supplemented with Extra-Virgin Olive Oil or Nuts. N. Engl. J. Med. 2018, 378, e34. [Google Scholar] [CrossRef] [PubMed]
- Tosti, V.; Bertozzi, B.; Fontana, L. Health Benefits of the Mediterranean Diet: Metabolic and Molecular Mechanisms. J. Gerontol. Ser. A Boil. Sci. Med. Sci. 2018, 73, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Schwingshackl, L.; Schwedhelm, C.; Galbete, C.; Hoffmann, G. Adherence to Mediterranean Diet and Risk of Cancer: An Updated Systematic Review and Meta-Analysis. Nutrients 2017, 9, 1063. [Google Scholar] [CrossRef]
- Soltani, S.; Jayedi, A.; Shab-Bidar, S.; Becerra-Tomás, N.; Salas-Salvadó, J. Adherence to the Mediterranean Diet in Relation to All-Cause Mortality: A Systematic Review and Dose-Response Meta-Analysis of Prospective Cohort Studies. Adv. Nutr. 2019, 10, 1029–1039. [Google Scholar] [CrossRef]
- Mancini, J.G.; Filion, K.B.; Atallah, R.; Eisenberg, M.J. Systematic Review of the Mediterranean Diet for Long-Term Weight Loss. Am. J. Med. 2016, 129, 407–415. [Google Scholar] [CrossRef]
- Cani, P.D.; Delzenne, P.D.C.A.N.M. The Role of the Gut Microbiota in Energy Metabolism and Metabolic Disease. Curr. Pharm. Des. 2009, 15, 1546–1558. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Christiansen, C.B.; Gabe, M.B.N.; Svendsen, B.; Dragsted, L.O.; Rosenkilde, M.M.; Holst, J.J. The impact of short-chain fatty acids on GLP-1 and PYY secretion from the isolated perfused rat colon. Am. J. Physiol. Liver Physiol. 2018, 315, G53–G65. [Google Scholar] [CrossRef]
- De Vadder, F.; Kovatcheva-Datchary, P.; Goncalves, D.; Vinera, J.; Zitoun, C.; Duchampt, A.; Bäckhed, F.; Mithieux, G. Microbiota-Generated Metabolites Promote Metabolic Benefits via Gut-Brain Neural Circuits. Cell 2014, 156, 84–96. [Google Scholar] [CrossRef]
- Li, Z.; Yi, C.-X.; Katiraei, S.; Kooijman, S.; Zhou, E.; Chung, C.K.; Gao, Y.; Heuvel, J.K.V.D.; Meijer, O.C.; Berbée, J.F.P.; et al. Butyrate reduces appetite and activates brown adipose tissue via the gut-brain neural circuit. Gut 2017, 67, 1269–1279. [Google Scholar] [CrossRef] [PubMed]
- Torres-Fuentes, C.; Golubeva, A.V.; Zhdanov, A.V.; Wallace, S.; Arboleya, S.; Papkovsky, D.B.; El Aidy, S.; Ross, P.; Roy, B.L.; Stanton, C.; et al. Short-chain fatty acids and microbiota metabolites attenuate ghrelin receptor signaling. FASEB J. 2019, 33, 13546–13559. [Google Scholar] [CrossRef] [PubMed]
- Barrea, L.; Muscogiuri, G.; Annunziata, G.; Laudisio, D.; Pugliese, G.; Salzano, C.; Colao, A.; Savastano, S. From gut microbiota dysfunction to obesity: Could short-chain fatty acids stop this dangerous course? Hormones 2019, 18, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; Van Der Beek, C.M.; Jocken, J.W.E.; Goossens, G.H.; Holst, J.J.; Damink, S.W.M.O.; Lenaerts, K.; DeJong, C.H.C.; Blaak, E.E. Colonic infusions of short-chain fatty acid mixtures promote energy metabolism in overweight/obese men: A randomized crossover trial. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Harris, L.; Hamilton, S.; Azevedo, L.B.; Olajide, J.; De Brún, C.; Waller, G.; Whittaker, V.; Sharp, T.; Lean, M.; Hankey, C.; et al. Intermittent fasting interventions for treatment of overweight and obesity in adults: A systematic review and meta-analysis. JBI Database Syst. Rev. Implement. Rep. 2018, 16, 507–547. [Google Scholar] [CrossRef] [PubMed]
- Patterson, R.E.; Laughlin, G.A.; LaCroix, A.Z.; Hartman, S.J.; Natarajan, L.; Senger, C.M.; Martínez, M.E.; Villaseñor, A.; Sears, D.D.; Marinac, C.R.; et al. Intermittent fasting and human metabolic health. J. Acad. Nutr. Diet. 2015, 115, 1203–1212. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P.; Moehl, K.; Ghena, N.; Schmaedick, M.; Cheng, A. Intermittent metabolic switching, neuroplasticity and brain health. Nat. Rev. Neurosci. 2018, 19, 81–94. [Google Scholar] [CrossRef]
- Freire, R.H. Scientific evidence of diets for weight loss: Different macronutrient composition, intermittent fasting, and popular diets. Nutrition 2020, 69, 110549. [Google Scholar] [CrossRef]
- Gabbianelli, R.; Bordoni, L.; Morano, S.; Agius, J.C.; Lalor, J. Nutri-Epigenetics and Gut Microbiota: How Birth Care, Bonding and Breastfeeding Can Influence and Be Influenced? Int. J. Mol. Sci. 2020, 21, 5032. [Google Scholar] [CrossRef]
- Heijmans, B.T.; Tobi, E.W.; Stein, A.D.; Putter, H.; Blauw, G.J.; Susser, E.S.; Slagboom, P.E.; Lumey, L.H. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 17046–17049. [Google Scholar] [CrossRef]
- Shen, L.; Li, C.; Wang, Z.; Zhang, R.; Shen, Y.; Miles, T.; Wei, J.; Zou, Z. Early-life exposure to severe famine is associated with higher methylation level in the IGF2 gene and higher total cholesterol in late adulthood: The Genomic Research of the Chinese Famine (GRECF) study. Clin. Epigenetics 2019, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Li, Y. Epigenetic Mechanisms Link Maternal Diets and Gut Microbiome to Obesity in the Offspring. Front. Genet. 2018, 9, 342. [Google Scholar] [CrossRef] [PubMed]
- Milagro, F.; Mansego, M.; De Miguel, C.; Martinez, J.A. Dietary factors, epigenetic modifications and obesity outcomes: Progresses and perspectives. Mol. Asp. Med. 2013, 34, 782–812. [Google Scholar] [CrossRef] [PubMed]
- Keleher, M.R.; Zaidi, R.; Hicks, L.; Shah, S.; Xing, X.; Li, D.; Wang, T.; Cheverud, J. A high-fat diet alters genome-wide DNA methylation and gene expression in SM/J mice. BMC Genom. 2018, 19, 888. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Tsujimoto, K.; Hashimoto, K.; Kawahori, K.; Hanzawa, N.; Hamaguchi, M.; Seki, T.; Nawa, M.; Ehara, T.; Kitamura, Y. Epigenetic modulation of Fgf21 in the perinatal mouse liver ameliorates diet-induced obesity in adulthood. Nat. Commun. 2018, 9, 636. [Google Scholar] [CrossRef]
- Madkour, M.I.; El-Serafi, A.T.; Jahrami, H.A.; Sherif, N.M.; Hassan, R.E.; Awadallah, S.; Faris, M.A.-I.E. Ramadan diurnal intermittent fasting modulates SOD2, TFAM, Nrf2, and sirtuins (SIRT1, SIRT3) gene expressions in subjects with overweight and obesity. Diabetes Res. Clin. Pr. 2019, 155, 107801. [Google Scholar] [CrossRef]
- Lowe, M.R.; Benson, L.; Singh, S. Individual differences in within-subject weight variability: There’s a signal in the noise. Physiol. Behav. 2020, 226, 113112. [Google Scholar] [CrossRef]
- Bagnall, A.-M.; Radley, D.; Jones, R.; Gately, P.; Nobles, J.; Van Dijk, M.; Blackshaw, J.; Montel, S.; Sahota, P. Whole systems approaches to obesity and other complex public health challenges: A systematic review. BMC Public Health 2019, 19, 8. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Targeting Individual Behaviors |
| Mass media campaigns |
| Nutrition counselling and education in healthcare settings |
| Targeting Food and Built Environment |
| Restaurant menu labelling |
| Dietary guidelines/standards |
| Advertisement restrictions |
| Interventions targeting schools and universities |
| Regulations/policies |
| Fiscal measures including taxes and subsidies |
| Reference | Sample Size | Duration | Comparison | Result |
|---|---|---|---|---|
| Hooper et al. (2012) [49] | 73,589 adults with normal weight, overweight or obesity | 6 m–7.5 years | Low fat (>30%) vs. Normal Fat | Low fat diets produced greater weight loss by 1.57 kg (95% CI −2.0 to −1.2 kg) |
| Wycherley et al. (2012) [50] | 1063 adults, BMI status not specified | >12 weeks | High protein (27–35%) vs. Low fat | High protein diets produced greater weight loss by 0.79 kg (95% CI −1.5 to −0.08 kg) |
| Ajala et al. (2013) [51] | 3073 adults with normal weight, overweight or obesity with or without diabetes | >6 months | Low carbohydrate vs. Mediterranean diet vs. High protein | Mediterranean diet and low carbohydrate diets produced greater weight loss than high protein by 1.84 and 0.69 kg respectively |
| Bueno et al. (2013) [52] | 1415 adults, BMI status not specified | ≥12 months | Very low carbohydrate (<50 g) vs. low fat (<30%, energy restricted) | Very low carbohydrate diets achieved greater weight loss by 0.91 (95% CI −1.65 to −0.17 kg) |
| Naude et al. (2014) [53] | 3209 adults with overweight or obesity with or without diabetes | >12 weeks | Low carbohydrate vs. isocaloric balanced diets | No difference |
| Clifton et al. (2014) [54] | 3492 adults with overweight or obesity | >12 months | High protein (>25%) vs. usual diet | High protein diets produced greater weight loss by 1 kg |
| Tobias et al. (2015) [55] | 68,128 adults, BMI status not specified | ≥12 months | Low fat vs. usual diets, low carbohydrate, high fat | Low carbohydrate diets produced greater weight loss by 1.15 kg (95% CI −0.52 to −1.79) |
| Boaz et al. (2015) [56] | 1161 adults with normal weight, overweight or obesity | >12 weeks | Low carbohydrate vs. Low fat | No difference |
| Mansoor et al. (2016) [57] | 1369 adults, BMI status not specified | ≥6 months | Low fat vs. low carbohydrates | Low carbohydrate diets produced greater weight loss by 2.17 kg (95% CI −0.99 to −3.36 kg) |
| Ge et al. (2020) [58] | 21,942 adults with overweight or obesity | 6–12 m | 14 popular diets including low fat, low carbohydrate, Mediterranean Diet | Low carbohydrate and low-fat diets produced similar weight loss, greater than Mediterranean diet |
| Nutritional Factors | Potential Epigenetic Mechanisms | Associated Metabolic Outcomes |
|---|---|---|
| Caloric restriction/undernutrition | DNA methylation Histone modifications miRNA | Increased risk of obesity and metabolic disorders (including insulin resistance and diabetes) in adulthood |
| Fatty Acids | ||
| Unsaturated FA (MUFA, n-3 PUFA) | DNA methylation Histone modifications miRNA | Prevention of metabolic derangements (lipid metabolism disturbances, inflammation, and insulin resistance) or chronic diseases (obesity, diabetes, non-alcoholic fatty liver disease, CVD risk and some types of cancer) (Becerra, 2019) |
| n-6 PUFA, Saturated and trans FA | DNA methylation Histone modifications miRNA | Associated with the presence or development of obesity, diabetes, inflammatory profile, atherosclerosis, hyperglycemia, insulin resistance, lipid alterations, lipotoxicity, dysregulation of lipid metabolism, and abnormal lipid accumulation (Becerra, 2019) |
| SCFA | DNA methylation Histone modifications Interaction with Gut Microbiome | Mixed effects |
| Methyl donors (e.g., folate, methionine, choline) * | DNA methylation | Increased risk of metabolic disorders including insulin resistance, diabetes and obesity |
| Probiotics/Fibers | Production of SCFA (same as SCFA) | Same as SCFA |
| Vitamins and minerals (e.g., retinol, ascorbate, tocopherol) | DNA methylation Histone modifications | Antioxidant effects |
| Polyphenols and other plant compounds | Inhibition of DNA hypermethylation Histone modification miRNA | May protect against carcinogenesis and obesity on adulthood |
| Dietary Patterns | ||
| High fat diet/Western diet | DNA methylation of obesity related genes leading to changes in genes expression and appetite regulation | - Imbalance in fatty acids causing oxidative stress and inflammatory state - Increased risk of obesity and metabolic disorders (including insulin resistance and diabetes) in adulthood |
| Mediterranean diet | DNA methylation | Protective against CVD risk and cancer |
| Intermittent Fasting | Improve the expression of anti-oxidant and anti-inflammatory genes | May be protective against oxidative stress |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwalla, N.; Jaafar, Z. Dietary Management of Obesity: A Review of the Evidence. Diagnostics 2021, 11, 24. https://doi.org/10.3390/diagnostics11010024
Hwalla N, Jaafar Z. Dietary Management of Obesity: A Review of the Evidence. Diagnostics. 2021; 11(1):24. https://doi.org/10.3390/diagnostics11010024
Chicago/Turabian StyleHwalla, Nahla, and Zeinab Jaafar. 2021. "Dietary Management of Obesity: A Review of the Evidence" Diagnostics 11, no. 1: 24. https://doi.org/10.3390/diagnostics11010024
APA StyleHwalla, N., & Jaafar, Z. (2021). Dietary Management of Obesity: A Review of the Evidence. Diagnostics, 11(1), 24. https://doi.org/10.3390/diagnostics11010024