Increased Vascular Adhesion Protein 1 (VAP-1) Levels Are Associated with Alternative M2 Macrophage Activation and Poor Prognosis for Human Gliomas

 ,
,
Abstract
1. Introduction
2. Materials and Methods
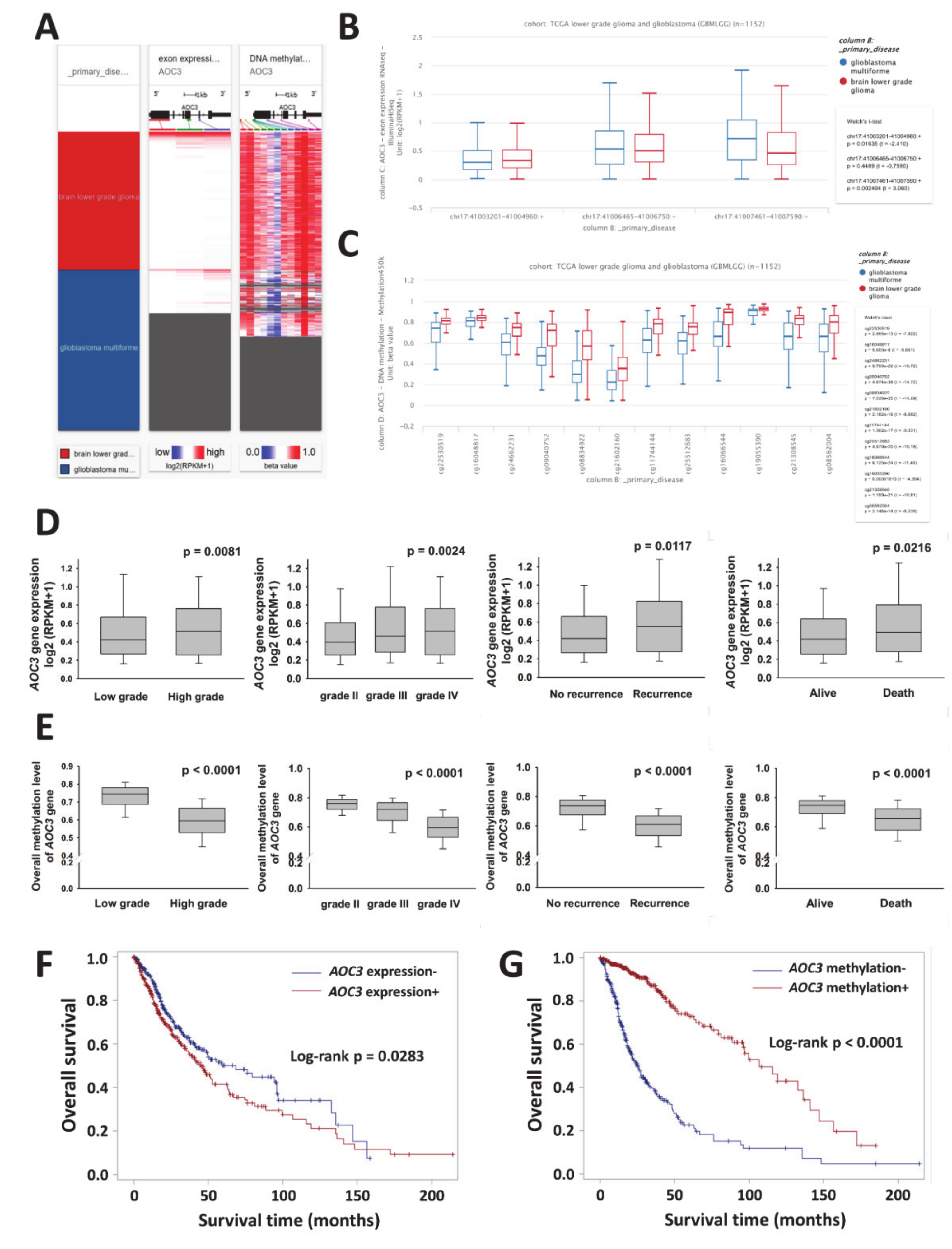
2.1. AOC3 Exon Expression and DNA Methylation Datasets
2.2. Specimens
2.3. Immunohistochemistry (IHC)
2.4. Pathologic Evaluation
2.5. Statistical Analysis
3. Results
3.1. VAP-1(AOC3) Expression as a Potential Biomarker for Prognosis in Patients with Gliomas
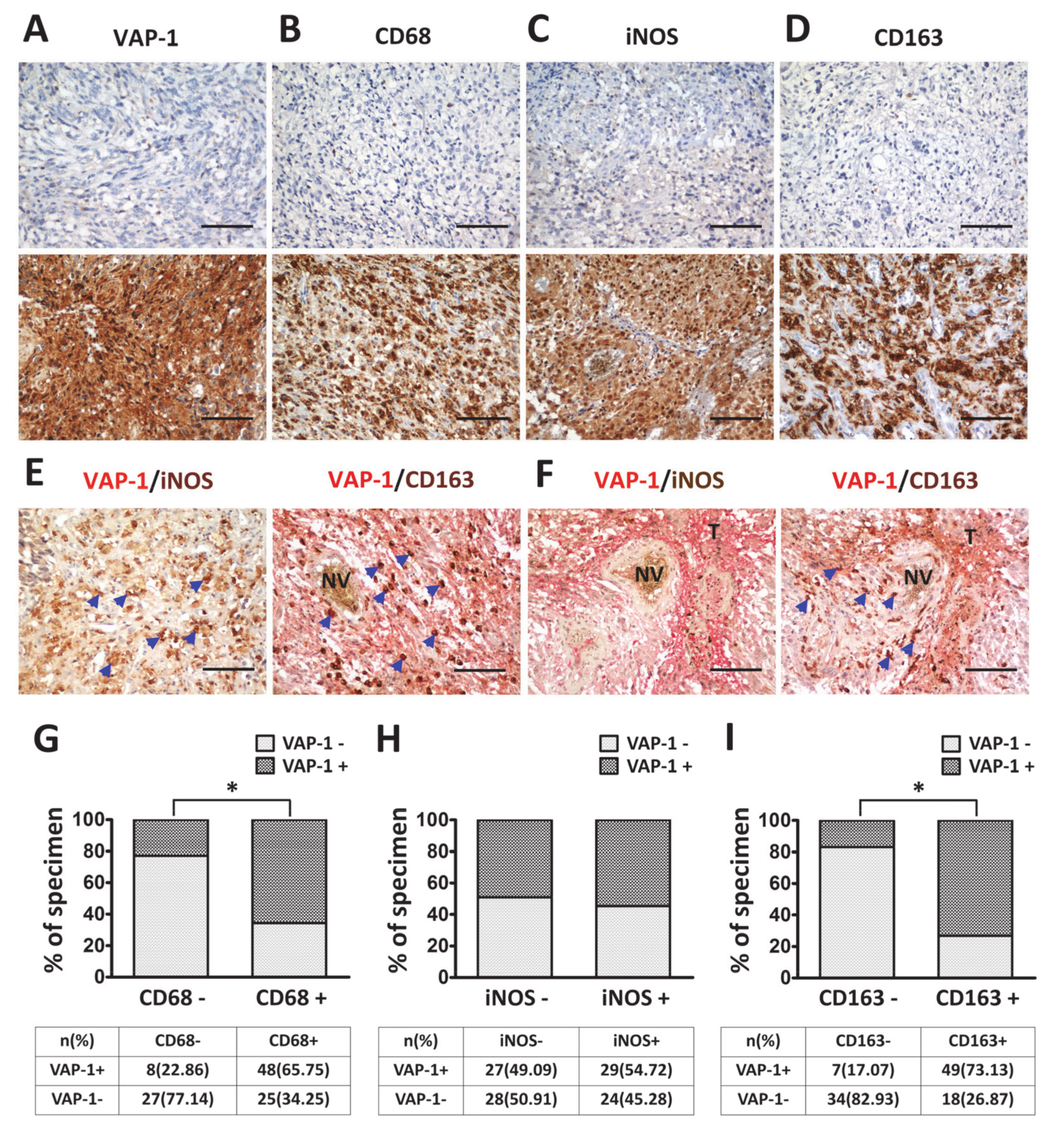
3.2. VAP-1 Expression and Co-Expressed with TAM Biomarkers are Elevated in Malignant Glioma Tissues
3.3. VAP-1 Alone and VAP-1/TAM Coexpression Correlated with Clinicopathological Variables in Glioma Patients
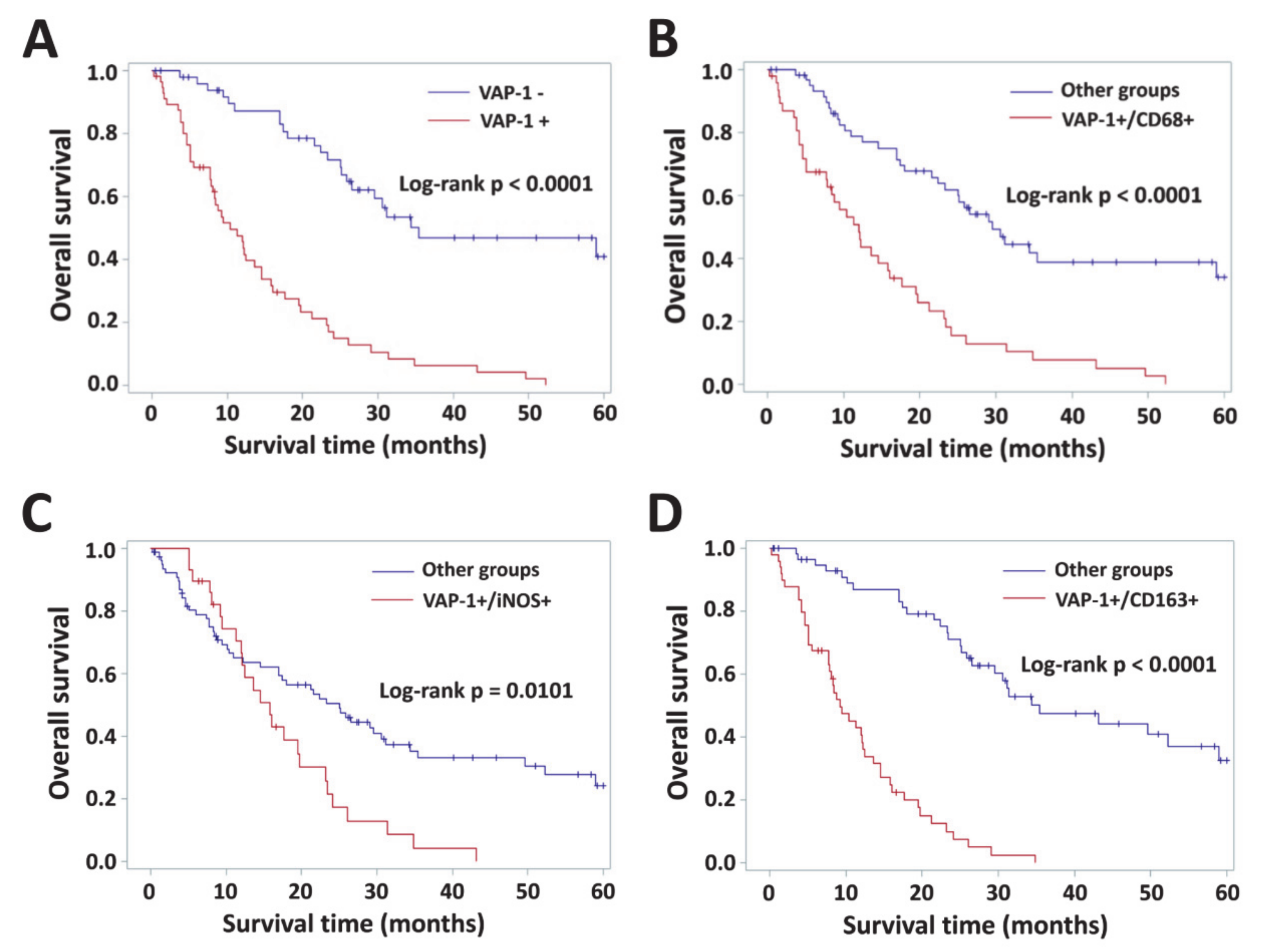
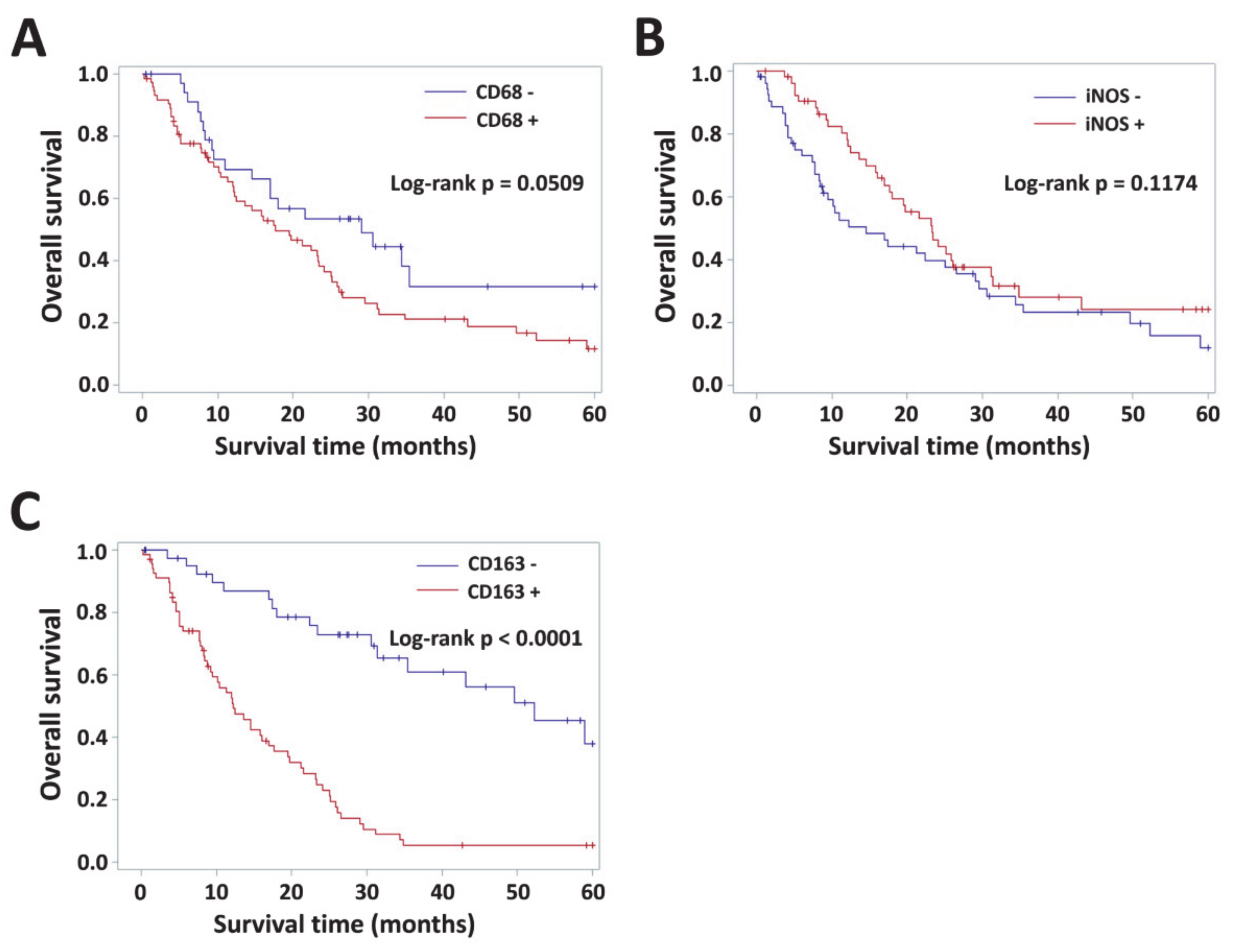
3.4. Impact of VAP-1 and VAP-1/TAM Phenotype on the Survival of Glioma Patients
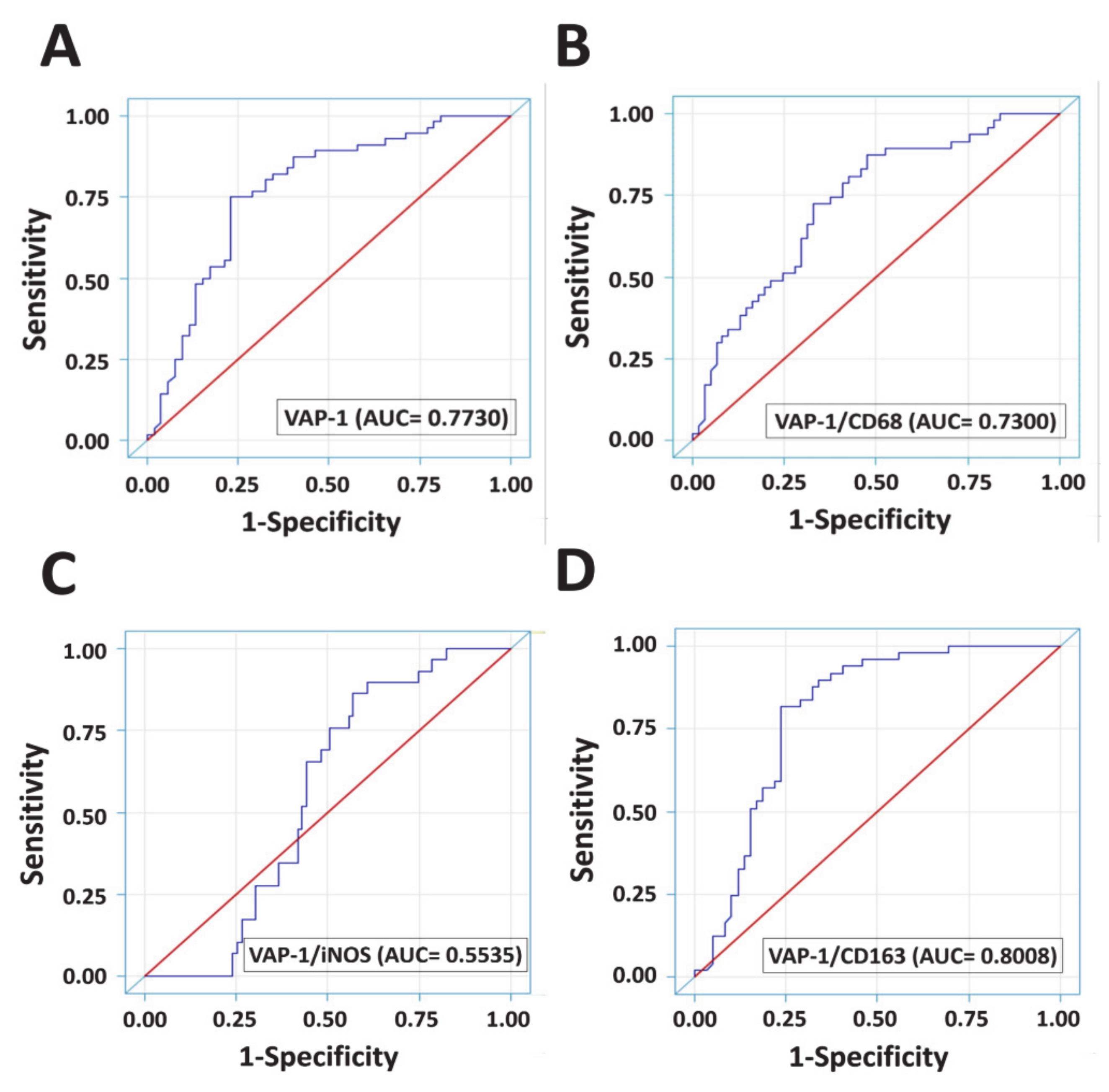
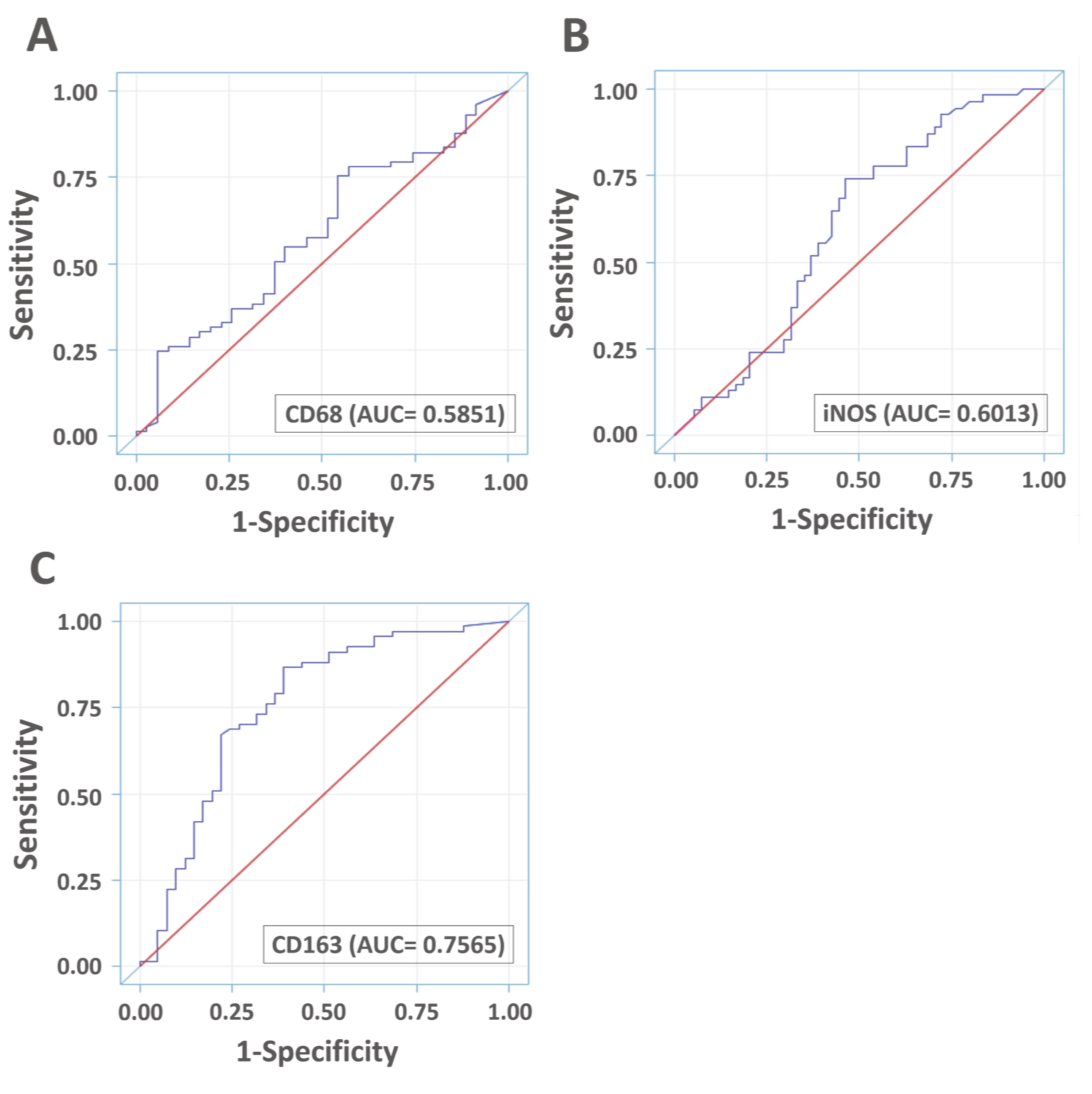
3.5. Diagnostic Accuracies of VAP-1 Expression and VAP-1/TAM Coexpression
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


References
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Cheng, W.; Ren, X.; Wang, Z.; Liu, X.; Li, G.; Han, S.; Jiang, T.; Wu, A. Tumor Purity as an Underlying Key Factor in Glioma. Clin. Cancer Res. 2017, 23, 6279–6291. [Google Scholar] [CrossRef] [PubMed]
- Golebiewska, A.; Bougnaud, S.; Stieber, D.; Brons, N.H.; Vallar, L.; Hertel, F.; Klink, B.; Schrock, E.; Bjerkvig, R.; Niclou, S.P. Side population in human glioblastoma is non-tumorigenic and characterizes brain endothelial cells. Brain 2013, 136, 1462–1475. [Google Scholar] [CrossRef][Green Version]
- Hambardzumyan, D.; Gutmann, D.H.; Kettenmann, H. The role of microglia and macrophages in glioma maintenance and progression. Nat. Neurosci. 2016, 19, 20–27. [Google Scholar] [CrossRef]
- Graeber, M.B.; Scheithauer, B.W.; Kreutzberg, G.W. Microglia in brain tumors. Glia 2002, 40, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Watters, J.J.; Schartner, J.M.; Badie, B. Microglia function in brain tumors. J. Neurosci. Res. 2005, 81, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.Y.; Watabe, K. The roles of microglia/macrophages in tumor progression of brain cancer and metastatic disease. Front Biosci. (Landmark Ed) 2017, 22, 1805–1829. [Google Scholar]
- Qian, B.Z.; Pollard, J.W. Macrophage diversity enhances tumor progression and metastasis. Cell 2010, 141, 39–51. [Google Scholar] [CrossRef]
- Ruffell, B.; Affara, N.I.; Coussens, L.M. Differential macrophage programming in the tumor microenvironment. Trends Immunol. 2012, 33, 119–126. [Google Scholar] [CrossRef]
- Lewis, C.E.; Harney, A.S.; Pollard, J.W. The Multifaceted Role of Perivascular Macrophages in Tumors. Cancer Cell 2016, 30, 18–25. [Google Scholar] [CrossRef]
- Tan, H.Y.; Wang, N.; Li, S.; Hong, M.; Wang, X.; Feng, Y. The Reactive Oxygen Species in Macrophage Polarization: Reflecting Its Dual Role in Progression and Treatment of Human Diseases. Oxidative Med. Cell. Longev. 2016, 2016, 2795090. [Google Scholar] [CrossRef]
- Jarosz-Biej, M.; Kaminska, N.; Matuszczak, S.; Cichon, T.; Pamula-Pilat, J.; Czapla, J.; Smolarczyk, R.; Skwarzynska, D.; Kulik, K.; Szala, S. M1-like macrophages change tumor blood vessels and microenvironment in murine melanoma. PLoS ONE 2018, 13, e0191012. [Google Scholar] [CrossRef]
- Lu, G.; Zhang, R.; Geng, S.; Peng, L.; Jayaraman, P.; Chen, C.; Xu, F.; Yang, J.; Li, Q.; Zheng, H.; et al. Myeloid cell-derived inducible nitric oxide synthase suppresses M1 macrophage polarization. Nat. Commun. 2015, 6, 6676. [Google Scholar] [CrossRef] [PubMed]
- Ley, K. M1 Means Kill; M2 Means Heal. J. Immunol. 2017, 199, 2191–2193. [Google Scholar] [CrossRef]
- Kerkar, S.P.; Restifo, N.P. Cellular constituents of immune escape within the tumor microenvironment. Cancer Res. 2012, 72, 3125–3130. [Google Scholar] [CrossRef]
- Heusinkveld, M.; van der Burg, S.H. Identification and manipulation of tumor associated macrophages in human cancers. J. Transl. Med. 2011, 9, 216. [Google Scholar] [CrossRef]
- Jayasingam, S.D.; Citartan, M.; Thang, T.H.; Mat Zin, A.A.; Ang, K.C.; Ch’ng, E.S. Evaluating the Polarization of Tumor-Associated Macrophages Into M1 and M2 Phenotypes in Human Cancer Tissue: Technicalities and Challenges in Routine Clinical Practice. Front. Oncol. 2019, 9, 1512. [Google Scholar] [CrossRef]
- Shabo, I.; Svanvik, J. Expression of macrophage antigens by tumor cells. Adv. Exp. Med. Biol. 2011, 714, 141–150. [Google Scholar] [CrossRef]
- Allavena, P.; Sica, A.; Solinas, G.; Porta, C.; Mantovani, A. The inflammatory micro-environment in tumor progression: The role of tumor-associated macrophages. Crit. Rev. Oncol./Hematol. 2008, 66, 1–9. [Google Scholar] [CrossRef]
- Roszer, T. Understanding the Mysterious M2 Macrophage through Activation Markers and Effector Mechanisms. Mediat. Inflamm. 2015, 2015, 816460. [Google Scholar] [CrossRef]
- Komohara, Y.; Hirahara, J.; Horikawa, T.; Kawamura, K.; Kiyota, E.; Sakashita, N.; Araki, N.; Takeya, M. AM-3K, an anti-macrophage antibody, recognizes CD163, a molecule associated with an anti-inflammatory macrophage phenotype. J. Histochem. Cytochem.: Off. J. Histochem. Soc. 2006, 54, 763–771. [Google Scholar] [CrossRef]
- Sica, A.; Schioppa, T.; Mantovani, A.; Allavena, P. Tumour-associated macrophages are a distinct M2 polarised population promoting tumour progression: Potential targets of anti-cancer therapy. Eur. J. Cancer 2006, 42, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Strilic, B.; Offermanns, S. Intravascular Survival and Extravasation of Tumor Cells. Cancer Cell 2017, 32, 282–293. [Google Scholar] [CrossRef] [PubMed]
- Reymond, N.; d’Agua, B.B.; Ridley, A.J. Crossing the endothelial barrier during metastasis. Nat. Rev. Cancer 2013, 13, 858–870. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Manaenko, A.; Khatibi, N.H.; Chen, W.; Zhang, J.H.; Tang, J. Vascular adhesion protein-1 inhibition provides antiinflammatory protection after an intracerebral hemorrhagic stroke in mice. J. Cereb. Blood Flow Metab. 2011, 31, 881–893. [Google Scholar] [CrossRef] [PubMed]
- Elo, P.; Tadayon, S.; Liljenback, H.; Teuho, J.; Kakela, M.; Koskensalo, K.; Saunavaara, V.; Virta, J.; Veres, T.Z.; Kiviniemi, A.; et al. Vascular adhesion protein-1 is actively involved in the development of inflammatory lesions in rat models of multiple sclerosis. J. Neuroinflammation 2018, 15, 128. [Google Scholar] [CrossRef]
- Sole, M.; Esteban-Lopez, M.; Taltavull, B.; Fabregas, C.; Fado, R.; Casals, N.; Rodriguez-Alvarez, J.; Minano-Molina, A.J.; Unzeta, M. Blood-brain barrier dysfunction underlying Alzheimer’s disease is induced by an SSAO/VAP-1-dependent cerebrovascular activation with enhanced Abeta deposition. Biochim. Biophys Acta Mol. Basis. Dis. 2019. [Google Scholar] [CrossRef]
- Jalkanen, S.; Karikoski, M.; Mercier, N.; Koskinen, K.; Henttinen, T.; Elima, K.; Salmivirta, K.; Salmi, M. The oxidase activity of vascular adhesion protein-1 (VAP-1) induces endothelial E- and P-selectins and leukocyte binding. Blood 2007, 110, 1864–1870. [Google Scholar] [CrossRef]
- Salmi, M.; Jalkanen, S. Vascular Adhesion Protein-1: A Cell Surface Amine Oxidase in Translation. Antioxid. Redox Signal. 2017. [Google Scholar] [CrossRef]
- Salmi, M.; Jalkanen, S. Cell-surface enzymes in control of leukocyte trafficking. Nat. Rev. Immunol. 2005, 5, 760–771. [Google Scholar] [CrossRef]
- Marttila-Ichihara, F.; Auvinen, K.; Elima, K.; Jalkanen, S.; Salmi, M. Vascular adhesion protein-1 enhances tumor growth by supporting recruitment of Gr-1+CD11b+ myeloid cells into tumors. Cancer Res. 2009, 69, 7875–7883. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Li, H.; Luo, H.J.; Lin, Z.X.; Jiang, Z.W.; Luo, W.H. SSAO inhibitors suppress hepatocellular tumor growth in mice. Cell Immunol. 2013, 283, 61–69. [Google Scholar] [CrossRef]
- Ferjancic, S.; Gil-Bernabe, A.M.; Hill, S.A.; Allen, P.D.; Richardson, P.; Sparey, T.; Savory, E.; McGuffog, J.; Muschel, R.J. VCAM-1 and VAP-1 recruit myeloid cells that promote pulmonary metastasis in mice. Blood 2013, 121, 3289–3297. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Mazzieri, R.; Yang, T.; Gobe, G.C. Translational Significance for Tumor Metastasis of Tumor-Associated Macrophages and Epithelial-Mesenchymal Transition. Front. Immunol. 2017, 8, 1106. [Google Scholar] [CrossRef] [PubMed]
- Mukaida, N.; Nosaka, T.; Nakamoto, Y.; Baba, T. Lung Macrophages: Multifunctional Regulator Cells for Metastatic Cells. Int. J. Mol. Sci. 2018. [Google Scholar] [CrossRef]
- Nakao, S.; Noda, K.; Zandi, S.; Sun, D.; Taher, M.; Schering, A.; Xie, F.; Mashima, Y.; Hafezi-Moghadam, A. VAP-1-mediated M2 macrophage infiltration underlies IL-1beta- but not VEGF-A-induced lymph- and angiogenesis. Am. J. Pathol. 2011, 178, 1913–1921. [Google Scholar] [CrossRef]
- Kostoro, J.; Chang, S.J.; Clark Lai, Y.C.; Wu, C.C.; Chai, C.Y.; Kwan, A.L. Overexpression of vascular adhesion protein-1 is associated with poor prognosis of astrocytomas. APMIS 2016, 124, 462–468. [Google Scholar] [CrossRef]
- Fukuhara, J.; Kase, S.; Noda, K.; Murata, M.; Noda, M.; Ando, R.; Dong, Z.; Kanda, A.; Ishida, S. Immunolocalization of vascular adhesion protein-1 in human conjunctival tumors. Ophthalmic Res. 2012, 48, 33–37. [Google Scholar] [CrossRef]
- Li, Y.I.; Hung, J.S.; Yu, T.Y.; Liou, J.M.; Wei, J.N.; Kao, H.L.; Chuang, L.M.; Shun, C.T.; Lee, P.H.; Lai, H.S.; et al. Serum vascular adhesion protein-1 predicts all-cause mortality and cancer-related mortality in subjects with colorectal cancer. Clin. Chim. Acta 2014, 428, 51–56. [Google Scholar] [CrossRef]
- Lai, Y.C.; Chang, S.J.; Kostoro, J.; Kwan, A.L.; Chai, C.Y. Vascular adhesion protein-1 as indicator of breast cancer tumor aggressiveness and invasiveness. APMIS 2018, 126, 755–761. [Google Scholar] [CrossRef]
- Varis, A.; Wolf, M.; Monni, O.; Vakkari, M.L.; Kokkola, A.; Moskaluk, C.; Frierson, H., Jr.; Powell, S.M.; Knuutila, S.; Kallioniemi, A.; et al. Targets of gene amplification and overexpression at 17q in gastric cancer. Cancer Res. 2002, 62, 2625–2629. [Google Scholar] [PubMed]
- Remmele, W.; Stegner, H.E. Recommendation for uniform definition of an immunoreactive score (IRS) for immunohistochemical estrogen receptor detection (ER-ICA) in breast cancer tissue. Der. Pathol. 1987, 8, 138–140. [Google Scholar]
- Fedchenko, N.; Reifenrath, J. Different approaches for interpretation and reporting of immunohistochemistry analysis results in the bone tissue - a review. Diagnostic Pathol. 2014, 9, 221. [Google Scholar] [CrossRef]
- Marttila-Ichihara, F.; Castermans, K.; Auvinen, K.; Oude Egbrink, M.G.; Jalkanen, S.; Griffioen, A.W.; Salmi, M. Small-molecule inhibitors of vascular adhesion protein-1 reduce the accumulation of myeloid cells into tumors and attenuate tumor growth in mice. J. Immunol. 2010, 184, 3164–3173. [Google Scholar] [CrossRef] [PubMed]
- Komohara, Y.; Ohnishi, K.; Kuratsu, J.; Takeya, M. Possible involvement of the M2 anti-inflammatory macrophage phenotype in growth of human gliomas. J. Pathol. 2008, 216, 15–24. [Google Scholar] [CrossRef]
- Annovazzi, L.; Mellai, M.; Bovio, E.; Mazzetti, S.; Pollo, B.; Schiffer, D. Microglia immunophenotyping in gliomas. Oncol. Lett. 2018, 15, 998–1006. [Google Scholar] [CrossRef]
- Kennedy, B.C.; Showers, C.R.; Anderson, D.E.; Anderson, L.; Canoll, P.; Bruce, J.N.; Anderson, R.C. Tumor-associated macrophages in glioma: Friend or foe? J. Oncol. 2013, 2013, 486912. [Google Scholar] [CrossRef]
- Gabrusiewicz, K.; Rodriguez, B.; Wei, J.; Hashimoto, Y.; Healy, L.M.; Maiti, S.N.; Thomas, G.; Zhou, S.; Wang, Q.; Elakkad, A.; et al. Glioblastoma-infiltrated innate immune cells resemble M0 macrophage phenotype. JCI Insight 2016. [Google Scholar] [CrossRef]
- Gutmann, D.H.; Kettenmann, H. Microglia/Brain Macrophages as Central Drivers of Brain Tumor Pathobiology. Neuron 2019, 104, 442–449. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Jetten, N.; Verbruggen, S.; Gijbels, M.J.; Post, M.J.; De Winther, M.P.; Donners, M.M. Anti-inflammatory M2, but not pro-inflammatory M1 macrophages promote angiogenesis in vivo. Angiogenesis 2014, 17, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Corliss, B.A.; Azimi, M.S.; Munson, J.M.; Peirce, S.M.; Murfee, W.L. Macrophages: An Inflammatory Link Between Angiogenesis and Lymphangiogenesis. Microcirculation 2016, 23, 95–121. [Google Scholar] [CrossRef] [PubMed]
- Murdoch, C.; Muthana, M.; Coffelt, S.B.; Lewis, C.E. The role of myeloid cells in the promotion of tumour angiogenesis. Nat. Rev. Cancer 2008, 8, 618–631. [Google Scholar] [CrossRef] [PubMed]
- Sica, A.; Larghi, P.; Mancino, A.; Rubino, L.; Porta, C.; Totaro, M.G.; Rimoldi, M.; Biswas, S.K.; Allavena, P.; Mantovani, A. Macrophage polarization in tumour progression. Semin. Cancer Biol. 2008, 18, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Pollard, J.W. Tumour-educated macrophages promote tumour progression and metastasis. Nat. Rev. Cancer 2004, 4, 71–78. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Median (Range) or n (%) |
|---|---|
| Total number of patients | 108 |
| Age (years) mean ± SD medians (corresponding ranges) | 50.26 ± 17.51 52.00 (20.00–83.00) |
| Tumor size (cm) mean ± SD medians (corresponding ranges) | 1.88 ± 1.32 1.60(0.20–7.70) |
| Follow-up of the patient cohort (months) mean ± SD medians (corresponding ranges) | 20.88 ± 17.39 16.82 (0.23–60.00) |
| Gender, n (%) Female Male | 40 (37.04) 68 (62.96) |
| WHO grade, n (%) II III IV | 27 (25.00) 35 (32.41) 46 (42.59) |
| Histological type, n (%) diffuse astrocytoma oligoastrocytoma oligodendroglioma anaplstic astrocytoma GBM | 28 (25.93) 10 (9.26) 3 (2.78) 28 (25.93) 39 (36.11) |
| Recurrence, n (%) Absent Present | 53 (49.07) 55 (50.93) |
| Survival status, n (%) survived died | 34 (31.48) 74 (68.52) |
| IDH1 mutant, n (%) Negative Positive | 81 (75.00) 27 (25.00) |
| Parameters | VAP-1- n (%) | VAP-1+ n (%) | p Value | Other Groups * n (%) | VAP-1+/ CD68+ n (%) | p Value | Other Groups ** n (%) | VAP-1+/ iNOS+ n (%) | p Value | Other Groups *** n (%) | VAP-1+/ CD163+ n (%) | p Value |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 52 (48.15) | 56 (51.85) | 61 (56.48) | 47 (43.52) | 79 (73.15) | 29 (26.85) | 59 (54.63) | 49 (45.37) | |||||
| Gender Female Male | 21 (40.38) 31 (59.62) | 19 (33.93) 37 (66.07) | 0.4876 | 25 (40.98) 36 (59.02) | 15(31.91) 32(68.09) | 0.3333 | 31 (39.24) 48 (60.76) | 9 (31.03) 20 (68.97) | 0.4338 | 24 (40.68) 35 (59.32) | 16 (32.65) 33 (67.35) | 0.3899 |
| Age ≤45 years >45 years | 25 (48.08) 27 (51.92) | 17 (30.36) 39 (69.64) | 0.0591 | 28 (45.90) 33 (54.10) | 14 (29.79) 33 (70.21) | 0.0885 | 33 (41.77) 46 (58.23) | 9 (31.03) 20 (68.97) | 0.3104 | 29 (49.15) 30 (50.85) | 13 (26.53) 36 (73.47) | 0.0164 |
| WHO grade II III IV | 24 (46.15) 21 (40.38) 7 (13.46) | 3 (5.36) 14 (25.00) 39 (69.64) | <0.0001 | 24 (39.34) 22 (36.07) 15 (24.59) | 3 (6.38) 13 (27.66) 31 (65.96) | <0.0001 | 26 (32.91) 31 (39.24) 22 (27.85) | 1 (3.45) 4 (13.79) 24 (82.76) | <0.0001 | 25 (42.37) 24 (40.68) 10 (16.95) | 2 (4.08) 11 (22.45) 36 (73.47) | <0.0001 |
| Tumor size <2 cm ≥2 cm | 33 (63.46) 19 (36.54) | 30 (53.57) 26 (46.43) | 0.2976 | 39 (63.93) 22 (36.07) | 24 (51.06) 23 (48.94) | 0.1786 | 47 (59.49) 32 (40.51) | 16 (55.17) 13 (44.83) | 0.6864 | 38 (64.41) 21 (35.59) | 25 (51.02) 24 (48.98) | 0.1601 |
| Recurrence Absent Present | 25 (48.08) 27 (51.92) | 28 (50.00) 28 (50.00) | 0.8417 | 30 (49.18) 31 (50.82) | 23 (48.94) 24 (51.06) | 0.9799 | 41 (51.90) 38 (48.10) | 12 (41.38) 17 (58.62) | 0.3325 | 29 (49.15) 30 (50.85) | 24 (48.98) 25 (51.02) | 0.9857 |
| Survival status survived died | 29 (55.77) 23 (44.23) | 5 (8.93) 51 (91.07) | <0.0001 | 29 (47.54) 32 (52.46) | 5 (10.64) 42 (89.36) | <0.0001 | 30 (37.97) 49 (62.03) | 4 (13.79) 25 (86.21) | 0.0165 | 30 (50.85) 29 (49.15) | 4 (8.16) 45 (91.84) | <0.0001 |
| IDH1 mutant Negative Positive | 34 (65.38) 18 (34.62) | 47 (83.93) 9 (16.07) | 0.0262 | 43 (70.49) 18 (29.51) | 38 (80.85) 9 (19.15) | 0.2177 | 57 (72.15) 22 (27.85) | 24 (82.76) 5 (17.24) | 0.2592 | 38 (64.41) 21 (35.59) | 43 (87.76) 6 (12.24) | 0.0053 |
| Variable | VAP-1 | CD68 | iNOS | CD163 | |||||
|---|---|---|---|---|---|---|---|---|---|
| Correlation | p Value | Correlation | p Value | Correlation | p Value | Correlation | p Value | ||
| VAP-1 | 1.00000 | – | 0.40181 | <0.0001 | 0.05629 | 0.5628 | 0.54450 | <0.0001 | |
| CD68 | 0.40181 | <0.0001 | 1.00000 | – | 0.04654 | 0.6325 | 0.35519 | 0.0002 | |
| iNOS | 0.05629 | 0.5628 | 0.04654 | 0.6325 | 1.00000 | – | 0.11909 | 0.2196 | |
| CD163 | 0.54450 | <0.0001 | 0.35519 | 0.0002 | 0.11909 | 0.2196 | 1.00000 | – | |
| Parameters | Univariate | Multivariate | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| HR (95% CI) | p-Value | VAP-1 | p-Value | VAP-1/CD68 | p-Value | VAP-1/iNOS | p-Value | VAP-1/CD163 | p-Value | |
| HR (95% CI) | HR (95% CI) | HR (95% CI) | HR (95% CI) | |||||||
| VAP-1 | 5.057 (3.016–8.481) | <0.0001 | 4.688(2.736–8.033) | <0.0001 | – | – | – | – | – | – |
| VAP-1/CD68 | 3.483 (2.164–5.605) | <0.0001 | – | – | 3.226 (1.980–5.256) | <0.0001 | – | – | – | – |
| VAP-1/iNOS | 1.908 (1.157–3.145) | 0.0113 | – | – | – | – | 1.572 (0.945–2.615) | 0.0817 | – | – |
| VAP-1/CD163 | 7.047 (4.085–12.155) | <0.0001 | – | – | – | – | 6.597 (3.677–11.836) | <0.0001 | ||
| Gender (female = 1) | 0.956 (0.597–1.531) | 0.8513 | 0.735 (0.426–1.267) | 0.2675 | 0.725 (0.419–1.252) | 0.2480 | 0.736 (0.436–1.240) | 0.2495 | 0.652 (0.372–1.140) | 0.1336 |
| Age | 1.853 (1.140–3.012) | 0.0129 | 1.816 (1.071–3.077) | 0.0266 | 1.886 (1.117–3.185) | 0.0176 | 1.817 (1.090–3.028) | 0.0219 | 1.755 (1.014–3.039) | 0.0445 |
| Tumor size | 1.231 (0.776–1.953) | 0.3765 | 1.645 (0.955–2.835) | 0.0730 | 1.474 (0.866–2.510) | 0.1526 | 1.411 (0.849–2.347) | 0.1843 | 1.438 (0.848–2.439) | 0.1774 |
| Recurrence | 0.924 (0.583–1.465) | 0.7368 | 0.848 (0.516–1.392) | 0.5144 | 0.900 (0.552–1.465) | 0.6709 | 1.043 (0.648–1.679) | 0.8619 | 0.782 (0.470–1.304) | 0.3464 |
| IDH1 mutant | 0.356 (0.191–0.662) | 0.0011 | 0.517 (0.270–0.993) | 0.0474 | 0.455 (0.239–0.866) | 0.0165 | 0.412 (0.216–0.782) | 0.0067 | 0.533 (0.279–1.019) | 0.0569 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, S.-J.; Tu, H.-P.; Lai, Y.-C.C.; Luo, C.-W.; Nejo, T.; Tanaka, S.; Chai, C.-Y.; Kwan, A.-L. Increased Vascular Adhesion Protein 1 (VAP-1) Levels Are Associated with Alternative M2 Macrophage Activation and Poor Prognosis for Human Gliomas. Diagnostics 2020, 10, 256. https://doi.org/10.3390/diagnostics10050256
Chang S-J, Tu H-P, Lai Y-CC, Luo C-W, Nejo T, Tanaka S, Chai C-Y, Kwan A-L. Increased Vascular Adhesion Protein 1 (VAP-1) Levels Are Associated with Alternative M2 Macrophage Activation and Poor Prognosis for Human Gliomas. Diagnostics. 2020; 10(5):256. https://doi.org/10.3390/diagnostics10050256
Chicago/Turabian StyleChang, Shu-Jyuan, Hung-Pin Tu, Yen-Chang Clark Lai, Chi-Wen Luo, Takahide Nejo, Shota Tanaka, Chee-Yin Chai, and Aij-Lie Kwan. 2020. "Increased Vascular Adhesion Protein 1 (VAP-1) Levels Are Associated with Alternative M2 Macrophage Activation and Poor Prognosis for Human Gliomas" Diagnostics 10, no. 5: 256. https://doi.org/10.3390/diagnostics10050256
APA StyleChang, S.-J., Tu, H.-P., Lai, Y.-C. C., Luo, C.-W., Nejo, T., Tanaka, S., Chai, C.-Y., & Kwan, A.-L. (2020). Increased Vascular Adhesion Protein 1 (VAP-1) Levels Are Associated with Alternative M2 Macrophage Activation and Poor Prognosis for Human Gliomas. Diagnostics, 10(5), 256. https://doi.org/10.3390/diagnostics10050256