Synthetic Mutualism and the Intervention Dilemma


Abstract
:1. Introduction
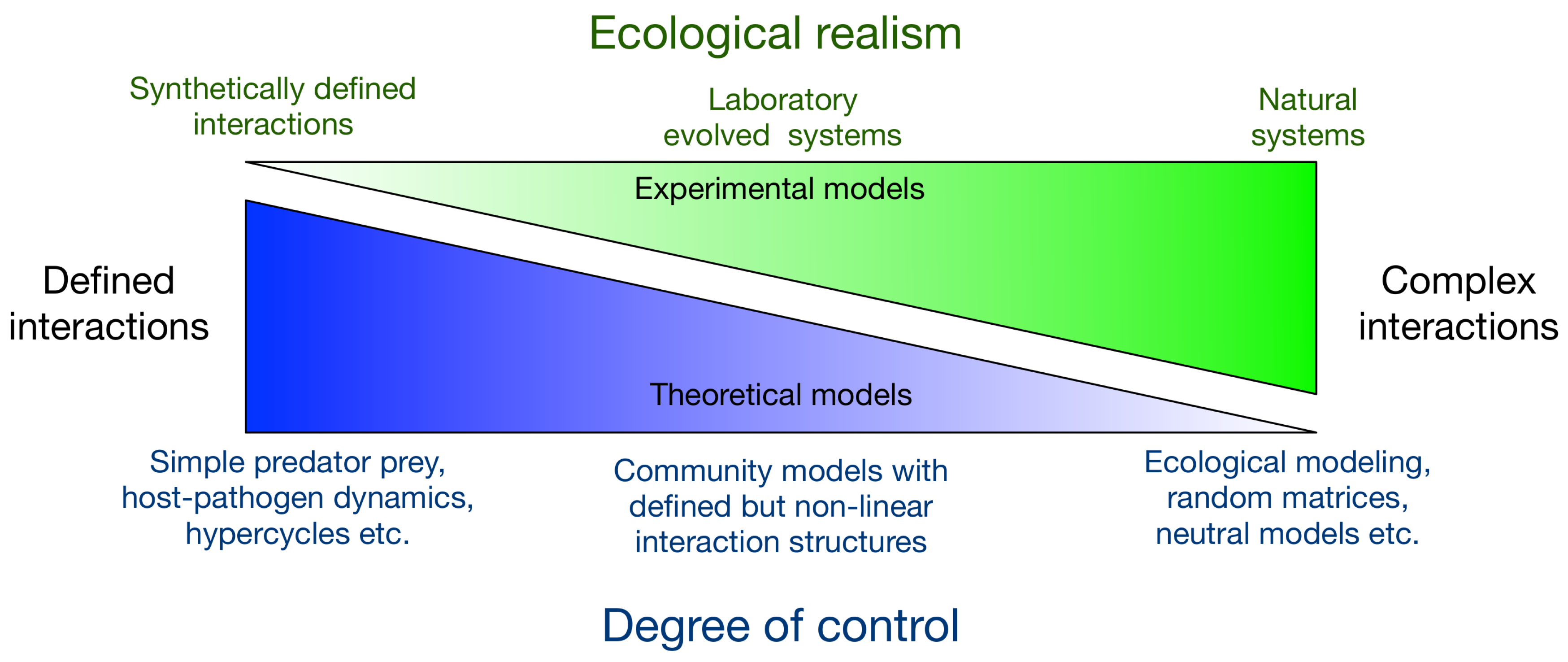
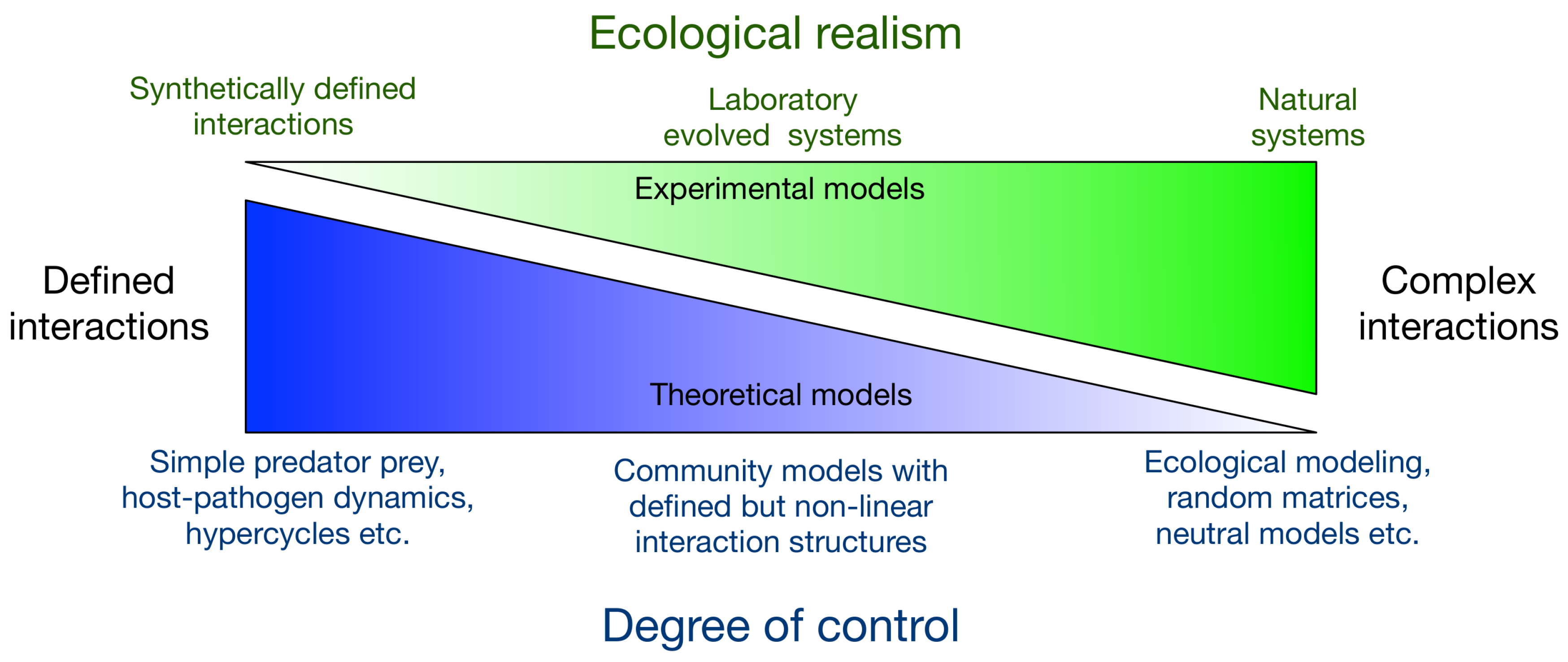
2. Experimental Modelling
2.1. Natural, Selected or Evolved Systems
2.2. Synthetic Systems
3. Integrated Modelling
3.1. Bridging the Gap
3.2. Towards More Complex Integration
3.3. Including the Complete Ecology
3.4. Extension to Evolved Symbiotic Systems
4. Future
4.1. Better Systems
4.2. Intervention
4.2.1. Modifying Life
4.2.2. Law and Implications
4.2.3. Altering the Extreme?
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Serrano, L. Synthetic biology: Promises and challenges. Mol. Syst. Biol. 2007, 3, 158. [Google Scholar] [CrossRef] [PubMed]
- Polizzi, K.M. What is synthetic biology? In Synthetic Biology; Polizzi, K.M., Kontoravdi, C., Eds.; Methods in Molecular Biology (Clifton, N.J.); Humana Press: Totowa, NJ, USA, 2013; pp. 3–6. [Google Scholar]
- Harcombe, W.R. Novel cooperation experimentally evolved between species. Evolution 2010, 64, 2166–2172. [Google Scholar] [CrossRef] [PubMed]
- Harcombe, W.R.; Chacón, J.M.; Adamowicz, E.M.; Chubiz, L.M.; Marx, C.J. Evolution of bidirectional costly mutualism from byproduct consumption. Proc. Natl. Acad. Sci. USA 2018, 115, 12000–12004. [Google Scholar] [CrossRef] [PubMed]
- Eigen, M. Selforganization of matter and the evolution of biological macromolecules. Die Naturwissenschaften 1971, 58, 465–523. [Google Scholar] [CrossRef] [PubMed]
- Farrell, J.M.; Brown, S.P. Evolution of bacterial trade in a two-species community. Proc. Natl. Acad. Sci. USA 2018, 115, 11874–11875. [Google Scholar] [CrossRef] [PubMed]
- Stat, M.; Morris, E.; Gates, R.D. Functional diversity in coral-dinoflagellate symbiosis. Proc. Natl. Acad. Sci. USA 2008, 105, 9256–9261. [Google Scholar] [CrossRef] [PubMed]
- De Fouw, J.; Govers, L.L.; van de Koppel, J.; van Belzen, J.; Dorigo, W.; Sidi Cheikh, M.A.; Christianen, M.J.A.; van der Reijden, K.J.; van der Geest, M.; Piersma, T.; et al. Drought, mutualism breakdown, and landscape-scale degradation of seagrass beds. Curr. Biol. 2016, 26, 1051–1056. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.M.; Wentrup, C.; Verna, C.; Knittel, K.; Dubilier, N. Origins and evolutionary flexibility of chemosynthetic symbionts from deep-sea animals. Biol. Bull. 2012, 223, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Alain, K.; Zbinden, M.; Le Bris, N.; Lesongeur, F.; Quérellou, J.; Gaill, F.; Cambon-Bonavita, M.A. Early steps in microbial colonization processes at deep-sea hydrothermal vents. Environ. Microbiol. 2004, 6, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Woyke, T.; Teeling, H.; Ivanova, N.N.; Huntemann, M.; Richter, M.; Gloeckner, F.O.; Boffelli, D.; Anderson, I.J.; Barry, K.W.; Shapiro, H.J.; et al. Symbiosis insights through metagenomic analysis of a microbial consortium. Nature 2006, 443, 950–955. [Google Scholar] [CrossRef] [PubMed]
- Low-Decarie, E.; Fussmann, G.F.; Dumbrell, A.J.; Bell, G. Communities that thrive in extreme conditions captured from a freshwater lake. Biol. Lett. 2016, 12, 20160562. [Google Scholar] [CrossRef] [PubMed]
- Freilich, S.; Zarecki, R.; Eilam, O.; Segal, E.S.; Henry, C.S.; Kupiec, M.; Gophna, U.; Sharan, R.; Ruppin, E. Competitive and cooperative metabolic interactions in bacterial communities. Nat. Commun. 2011, 2, 589. [Google Scholar] [CrossRef] [PubMed]
- Hoek, T.A.; Axelrod, K.; Biancalani, T.; Yurtsev, E.A.; Liu, J.; Gore, J. Resource availability modulates the cooperative and competitive nature of a microbial cross-feeding mutualism. PLoS Biol. 2016, 14, e1002540. [Google Scholar] [CrossRef] [PubMed]
- López-García, P.; Eme, L.; Moreira, D. Symbiosis in eukaryotic evolution. J. Theor. Biol. 2017, 434, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Sojo, V.; Herschy, B.; Whicher, A.; Camprubí, E.; Lane, N. The origin of life in alkaline hydrothermal vents. Astrobiology 2016, 16, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Schirrmeister, B.E.; de Vos, J.M.; Antonelli, A.; Bagheri, H.C. Evolution of multicellularity coincided with increased diversification of cyanobacteria and the Great Oxidation Event. Proc. Natl. Acad. Sci. USA 2013, 110, 1791–1796. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.M.; Freeman, C.J.; Wong, J.C.Y.; Fogel, M.L.; Knowlton, N. Climate change promotes parasitism in a coral symbiosis. ISME J. 2018, 12, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.E.; Gulledge, J.; Engelhaupt, E.; Burow, M.E.; McLachlan, J.A. Pesticides reduce symbiotic efficiency of nitrogen-fixing rhizobia and host plants. Proc. Natl. Acad. Sci. USA 2007, 104, 10282–10287. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.P.; McCormick, M.I.; Srinivasan, M.; Eagle, J.V. Coral decline threatens fish biodiversity in marine reserves. Proc Natl. Acad. Sci. USA 2004, 101, 8251–8253. [Google Scholar] [CrossRef] [PubMed]
- Wulff, J.L.; Buss, L.W. Do sponges help hold coral reefs together. Nature 1979, 281, 474–475. [Google Scholar] [CrossRef]
- Redford, K.H.; Adams, W.; Mace, G.M. Synthetic biology and conservation of nature: Wicked problems and wicked solutions. PLoS Biol. 2013, 11, e1001530. [Google Scholar] [CrossRef] [PubMed]
- Piaggio, A.J.; Segelbacher, G.; Seddon, P.J.; Alphey, L.; Bennett, E.L.; Carlson, R.H.; Friedman, R.M.; Kanavy, D.; Phelan, R.; Redford, K.H.; et al. Is it time for synthetic biodiversity conservation? Trends Ecol. Evol. 2017, 32, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Levin, R.A.; Voolstra, C.R.; Agrawal, S.; Steinberg, P.D.; Suggett, D.J.; van Oppen, M.J.H. Engineering strategies to decode and enhance the genomes of coral symbionts. Front. Microbiol. 2017, 8, 1220. [Google Scholar] [CrossRef] [PubMed]
- Kiers, E.T.; Rousseau, R.A.; West, S.A.; Denison, R.F. Host sanctions and the legume-rhizobium mutualism. Nature 2003, 425, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Spottiswoode, C.N.; Begg, K.S.; Begg, C.M. Reciprocal signaling in honeyguide-human mutualism. Science 2016, 353, 387–389. [Google Scholar] [CrossRef] [PubMed]
- Guezennec, J.; Ortega-Morales, O.; Raguenes, G.; Geesey, G. Bacterial colonization of artificial substrate in the vicinity of deep-sea hydrothermal vents. FEMS Microbiol. Ecol. 1998, 26, 89–99. [Google Scholar] [CrossRef]
- Schreier, J.E.; Van Dover, C.L. Hydrothermal vent ecology. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: New York, NY, USA, 2018. [Google Scholar]
- Ponnudurai, R.; Kleiner, M.; Sayavedra, L.; Petersen, J.M.; Moche, M.; Otto, A.; Becher, D.; Takeuchi, T.; Satoh, N.; Dubilier, N.; et al. Metabolic and physiological interdependencies in the Bathymodiolus azoricus symbiosis. ISME J. 2017, 11, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Kelly, L.C.; Cockell, C.S.; Thorsteinsson, T.; Marteinsson, V.; Stevenson, J. Pioneer microbial communities of the Fimmvörðuháls lava flow, Eyjafjallajökull, Iceland. Microb. Ecol. 2014, 68, 504–518. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, R.; Kim, S.W.; Sato, Y.; Oshima, K.; Hattori, M.; Kamijo, T.; Ohta, H. Unique pioneer microbial communities exposed to volcanic sulfur dioxide. Sci. Rep. 2016, 6, 19687. [Google Scholar] [CrossRef] [PubMed]
- Crews, T.E.; Kurina, L.M.; Vitousek, P.M. Organic matter and nitrogen accumulation and nitrogen fixation during early ecosystem development in Hawaii. Biogeochemistry 2001, 52, 259–279. [Google Scholar] [CrossRef]
- Noffke, N.; Christian, D.; Wacey, D.; Hazen, R.M. Microbially induced sedimentary structures recording an ancient ecosystem in the ca. 3.48 billion-year-old dresser formation, Pilbara, Western Australia. Astrobiology 2013, 13, 1103–1124. [Google Scholar] [CrossRef] [PubMed]
- Mack, K.M.L.; Rudgers, J.A. Balancing multiple mutualists: asymmetric interactions among plants, arbuscular mycorrhizal fungi, and fungal endophytes. OIKOS 2008, 117, 310–320. [Google Scholar] [CrossRef]
- Wang, B.; Yao, M.; Lv, L.; Ling, Z.; Engineering, L.L. The human microbiota in health and disease. Engineering 2017, 3, 71–82. [Google Scholar] [CrossRef]
- Kubo, I.; Hosoda, K.; Suzuki, S.; Yamamoto, K.; Kihara, K.; Mori, K.; Yomo, T. Construction of bacteria–eukaryote synthetic mutualism. Biosystems 2013, 113, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Cairns, J.; Jokela, R.; Hultman, J.; Tamminen, M.; Virta, M.; Hiltunen, T. Construction and characterization of synthetic bacterial community for experimental ecology and evolution. Front. Genet. 2018, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.P.; Supekova, L.; Chen, J.H.; Pestonjamasp, K.; Webster, P.; Ko, Y.; Henderson, S.C.; McDermott, G.; Supek, F.; Schultz, P.G. Engineering yeast endosymbionts as a step toward the evolution of mitochondria. Proc. Natl. Acad. Sci. USA 2018, 91, 201813143. [Google Scholar] [CrossRef] [PubMed]
- Bryk, J.; Reeves, R.G.; Reed, F.A.; Denton, J.A. Transcriptional effects of a positive feedback circuit in Drosophila melanogaster. BMC Genom. 2017, 18, 990. [Google Scholar] [CrossRef] [PubMed]
- Rogers, D.; Denton, J.; McConnell, E.; Greig, D. Experimental evolution of species recognition. Curr. Biol. 2015, 25, 1753–1758. [Google Scholar] [CrossRef] [PubMed]
- Shou, W.; Ram, S.; Vilar, J.M.G. Synthetic cooperation in engineered yeast populations. Proc. Natl. Acad. Sci. USA 2007, 104, 1877–1882. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.J.I.; Neugeboren, B.I.; Nelson, D.R.; Murray, A.W. Genetic drift opposes mutualism during spatial population expansion. Proc. Natl. Acad. Sci. USA 2014, 111, 1037–1042. [Google Scholar] [CrossRef] [PubMed]
- Hosoda, K.; Suzuki, S.; Yamauchi, Y.; Shiroguchi, Y.; Kashiwagi, A.; Ono, N.; Mori, K.; Yomo, T. Cooperative adaptation to establishment of a synthetic bacterial mutualism. PLoS ONE 2011, 6, e17105. [Google Scholar] [CrossRef] [PubMed]
- Pande, S.; Kaftan, F.; Lang, S.; Svatoš, A.; Germerodt, S.; Kost, C. Privatization of cooperative benefits stabilizes mutualistic cross-feeding interactions in spatially structured environments. ISME J. 2016, 10, 1413–1423. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Meldgin, D.R.; Collins, J.J.; Lu, T. Designing microbial consortia with defined social interactions. Nat. Chem. Biol. 2018, 14, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Armitt, S.; Woods, R.A. Purine-excreting mutants of Saccharomyces cerevisiae. I. Isolation and genetic analysis. Genet. Res. 1970, 15, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Gray, G.; Bhattacharjee, J.K. Biosynthesis of lysine in Saccharomyces cerevisiae: regulation of homocitrate synthase in analogue-resistant mutants. Microbiology 1976, 97, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Gokhale, C.S.; Papkou, A.; Traulsen, A.; Schulenburg, H. Lotka-Volterra dynamics kills the Red Queen: Population size fluctuations and associated stochasticity dramatically change host-parasite coevolution. BMC Evol. Biol. 2013, 13, 254. [Google Scholar] [CrossRef] [PubMed]
- Amor, D.R.; Montañez, R.; Duran-Nebreda, S.; Solé, R.V. Spatial dynamics of synthetic microbial mutualists and their parasites. PLoS Comput. Biol. 2017, 13, e1005689. [Google Scholar] [CrossRef] [PubMed]
- Momeni, B.; Waite, A.J.; Shou, W. Spatial self-organization favors heterotypic cooperation over cheating. eLife 2013, 2, e00960. [Google Scholar] [CrossRef] [PubMed]
- Denton, J.A.; Gokhale, C.S. Promoting synthetic symbiosis under environmental disturbances. bioRxiv, 2018; bioRxiv:395426. [Google Scholar]
- Kerner, A.; Park, J.; Williams, A.; Lin, X.N. A programmable Escherichia coli consortium via tunable symbiosis. PLoS ONE 2012, 7, e34032. [Google Scholar] [CrossRef] [PubMed]
- Eigen, M.; Schuster, P. The hypercycle. A principle of natural self-organization. Part B: The abstract hypercycle. Die Naturwissenschaften 1978, 65, 7–41. [Google Scholar] [CrossRef]
- Santala, S.; Karp, M.; Santala, V. Rationally engineered synthetic coculture for improved biomass and product formation. PLoS ONE 2014, 9, e113786. [Google Scholar] [CrossRef] [PubMed]
- May, R.M.; Anderson, R.M. Parasite host co-evolution. Parasitology 1990, 100, S89–S101. [Google Scholar] [CrossRef] [PubMed]
- Rabajante, J.F.; Tubay, J.M.; Uehara, T.; Morita, S.; Ebert, D.; Yoshimura, J. Red Queen dynamics in multi-host and multi-parasite interaction systems. Sci. Rep. 2015, 5, 10004. [Google Scholar] [CrossRef] [PubMed]
- Cadotte, M.W. Concurrent niche and neutral processes in the competition–colonization model of species coexistence. Proc. R. Soc. B Biol. Sci. 2007, 274, 2739–2744. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Sukumaran, J.; Wu, S.; Rodrigo, A. Neutral models of microbiome evolution. PLoS Comput. Biol. 2015, 11, e1004365. [Google Scholar] [CrossRef] [PubMed]
- Sieber, M.; Pita, L.; Weiland-Bräuer, N.; Dirksen, P.; Wang, J.; Mortzfeld, B.; Franzenburg, S.; Schmitz, R.A.; Baines, J.F.; Fraune, S.; et al. The neutral metaorganism. bioRxiv, 2018; bioRxiv:367243. [Google Scholar]
- Zomorrodi, A.R.; Segrè, D. Synthetic ecology of microbes: Mathematical models and applications. J. Mol. Biol. 2016, 428, 837–861. [Google Scholar] [CrossRef] [PubMed]
- Eigen, M.; Schuster, P. The hypercycle. A principle of natural self-organization. Part A: Emergence of the hypercycle. Die Naturwissenschaften 1977, 64, 541–565. [Google Scholar] [CrossRef] [PubMed]
- Maynard Smith, J.; Szathmáry, E. The Major Transitions in Evolution; Freeman, W.H., Ed.; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Maynard Smith, J. Hypercycles and the origin of life. Nature 1979, 280, 445–446. [Google Scholar] [CrossRef]
- Hofbauer, J. A general cooperation theorem for hypercycles. Monatshefte Math. 1981, 91, 233–240. [Google Scholar] [CrossRef]
- Hofbauer, J. A difference equation model for the hypercycle. SIAM J. Appl. Math. 1984, 44, 762–772. [Google Scholar] [CrossRef]
- Boerlijst, M.C.; Hogeweg, P. Spiral wave structure in pre-biotic evolution—Hypercycles stable against parasites. Phys. D Nonlinear Phenom. 1991, 48, 17–28. [Google Scholar] [CrossRef]
- Lee, D.H.; Severin, K.; Ghadiri, M.R. Autocatalytic networks: the transition from molecular self-replication to molecular ecosystems. Curr. Opin. Chem. Biol. 1997, 1, 491–496. [Google Scholar] [CrossRef]
- Sardanyes, J.; Solé, R.V. Bifurcations and phase transitions in spatially extended two-member hypercycles. J. Theor. Biol. 2006, 243, 468–482. [Google Scholar] [CrossRef] [PubMed]
- Hordijk, W.; Hein, J.; Steel, M. Autocatalytic sets and the origin of life. Entropy 2010, 12, 1733–1742. [Google Scholar] [CrossRef]
- Guillamon, A.; Fontich, E.; Sardanyés, J. Bifurcations analysis of oscillating hypercycles. J. Theor. Biol. 2015, 387, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, N.; Manapat, M.; Chen, I.A.; Xulvi-Brunet, R.; Hayden, E.J.; Lehman, N. Spontaneous network formation among cooperative RNA replicators. Nature 2012, 491, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Mizuuchi, R.; Ichihashi, N. Sustainable replication and coevolution of cooperative RNAs in an artificial cell-like system. Nat. Ecol. Evol. 2018, 2, 1654–1660. [Google Scholar] [CrossRef] [PubMed]
- Lundh, T.; Gerlee, P. Cross-feeding dynamics described by a series expansion of the replicator equation. Bull. Math. Biol. 2013, 75, 709–724. [Google Scholar] [CrossRef] [PubMed]
- Vessman, B.; Gerlee, P.; Lundh, T. Estimating the probability of coexistence in cross-feeding communities. J. Theor. Biol. 2016, 408, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Seth, E.C.; Taga, M.E. Nutrient cross-feeding in the microbial world. Front. Microbiol. 2014, 5, 350. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.; Higgins, L.M.; Gore, J. Community structure follows simple assembly rules in microbial microcosms. Nat. Ecol. Evol. 2017, 1, 109. [Google Scholar] [CrossRef] [PubMed]
- Momeni, B.; Xie, L.; Shou, W. Lotka-Volterra pairwise modeling fails to capture diverse pairwise microbial interactions. eLife 2017, 6, 205. [Google Scholar] [CrossRef] [PubMed]
- Momeni, B.; Brileya, K.A.; Fields, M.W.; Shou, W. Strong inter-population cooperation leads to partner intermixing in microbial communities. eLife 2013, 2, e00230. [Google Scholar] [CrossRef] [PubMed]
- Zomorrodi, A.R.; Segrè, D. Genome-driven evolutionary game theory helps understand the rise of metabolic interdependencies in microbial communities. Nat. Commun. 2017, 8, 6449. [Google Scholar] [CrossRef] [PubMed]
- Van Hoek, M.J.A.; Merks, R.M.H. Emergence of microbial diversity due to cross-feeding interactions in a spatial model of gut microbial metabolism. BMC Syst. Biol. 2017, 11, 56. [Google Scholar]
- Frickel, J.; Sieber, M.; Becks, L. Eco-evolutionary dynamics in a coevolving host–virus system. Ecol. Lett. 2016, 19, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Estrela, S.; Libby, E.; Van Cleve, J.; Débarre, F.; Deforet, M.; Harcombe, W.R.; Peña, J.; Brown, S.P.; Hochberg, M.E. Environmentally mediated social dilemmas. Trends Ecol. Evol. 2019, 34, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.M.; Catella, S.A.; Abbott, K.C. Population dynamics of mutualism and intraspecific density dependence: How θ-logistic density dependence affects mutualistic positive feedback. Ecol. Model. 2018, 368, 191–197. [Google Scholar] [CrossRef]
- Szathmary, E. The Origin of Replicators and Reproducers. Philos Trans. Biol. Sci. 2006, 361, 1761–1776. [Google Scholar] [CrossRef] [PubMed]
- Hogeweg, P.; Takeuchi, N. Multilevel selection in models of prebiotic evolution: Compartments and spatial self-organization. Orig. Life Evol. Biosph. 2003, 33, 375–403. [Google Scholar] [CrossRef] [PubMed]
- Kinsler, G.; Sinai, S.; Lee, N.K.; Nowak, M.A. Prebiotic selection for motifs in a model of template-free elongation of polymers within compartments. PLoS ONE 2017, 12, e0180208. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, S.; Kun, Á.; Ryckelynck, M.; Coldren, F.; Szilágyi, A.; Jossinet, F.; Rick, C.; Nghe, P.; Szathmary, E.; Griffiths, A.D. Transient compartmentalization of RNA replicators prevents extinction due to parasites. Science 2016, 354, 1293–1296. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.A. Evolutionary Dynamics; Harvard University Press: Cambridge, MA, USA, 2006. [Google Scholar]
- Akçay, E. Evolutionary models of mutualism. In Mutualism; Bronstein, J.L., Ed.; Oxford University Press: New York, NY, USA, 2015; pp. 57–76. [Google Scholar]
- Germerodt, S.; Bohl, K.; Lück, A.; Pande, S.; Schröter, A.; Kaleta, C.; Schuster, S.; Kost, C. Pervasive selection for cooperative cross-feeding in bacterial communities. PLoS Comput. Biol. 2016, 12, e1004986-21. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Gerchman, Y.; Collins, C.H.; Arnold, F.H.; Weiss, R. A synthetic multicellular system for programmed pattern formation. Nature 2005, 434, 1130–1134. [Google Scholar] [CrossRef] [PubMed]
- Brenner, K.; Karig, D.K.; Weiss, R.; Arnold, F.H. Engineered bidirectional communication mediates a consensus in a microbial biofilm consortium. Proc. Natl. Acad. Sci. USA 2007, 104, 17300–17304. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Blanchard, A.E.; Liao, C.; Lu, T. Engineering robust and tunable spatial structures with synthetic gene circuits. Nucleic Acids Res. 2017, 45, 1005–1014. [Google Scholar] [CrossRef] [PubMed]
- Campbell, K.; Vowinckel, J.; Mülleder, M.; Malmsheimer, S.; Lawrence, N.; Calvani, E.; Miller-Fleming, L.; Alam, M.T.; Christen, S.; Keller, M.A.; et al. Self-establishing communities enable cooperative metabolite exchange in a eukaryote. eLife 2015, 4, e09943. [Google Scholar] [CrossRef] [PubMed]
- Alon, U. Network motifs: Theory and experimental approaches. Nat. Rev. Genet. 2007, 8, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Feist, A.M.; Herrgård, M.J.; Thiele, I.; Reed, J.L.; Palsson, B.Ø. Reconstruction of biochemical networks in microorganisms. Nat. Rev. Microbiol. 2009, 7, 129–143. [Google Scholar] [CrossRef] [PubMed]
- Momeni, B.; Chen, C.C.; Hillesland, K.L.; Waite, A.; Shou, W. Using artificial systems to explore the ecology and evolution of symbioses. Cell. Mol. Life Sci. 2011, 68, 1353–1368. [Google Scholar] [CrossRef] [PubMed]
- Solé, R.V. Bioengineering the biosphere? Ecol. Complex. 2015, 22, 40–49. [Google Scholar] [CrossRef]
- Church, G.M. From systems biology to synthetic biology. Mol. Syst. Biol. 2005, 1, 2005.0032. [Google Scholar] [CrossRef] [PubMed]
- Escalante, A.E.; Rebolleda-Gómez, M.; Benítez, M.; Travisano, M. Ecological perspectives on synthetic biology: Insights from microbial population biology. Front. Microbiol. 2015, 6, 143. [Google Scholar] [CrossRef] [PubMed]
- Gossen, M.; Bujard, H. Tight control of gene-expression in mammalian-cells by tetracycline-responsive promoters. Proc. Natl. Acad. Sci. USA 1992, 89, 5547–5551. [Google Scholar] [CrossRef] [PubMed]
- Zeidler, M.P.; Tan, C.; Bellaiche, Y.; Cherry, S.; Häder, S.; Gayko, U.; Perrimon, N. Temperature-sensitive control of protein activity by conditionally splicing inteins. Nat. Biotechnol. 2004, 22, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Quarton, T.; Ehrhardt, K.; Lee, J.; Kannan, S.; Li, Y.; Ma, L.; Bleris, L. Mapping the operational landscape of microRNAs in synthetic gene circuits. NPJ Syst. Biol. Appl. 2018, 4, 6. [Google Scholar] [CrossRef] [PubMed]
- Dublanche, Y.; Michalodimitrakis, K.; Kümmerer, N.; Foglierini, M.; Serrano, L. Noise in transcription negative feedback loops: Simulation and experimental analysis. Mol. Syst. Biol. 2006, 2, 41. [Google Scholar] [CrossRef] [PubMed]
- Hensel, Z. A plasmid-based Escherichia coli gene expression system with cell-to-cell variation below the extrinsic noise limit. PLoS ONE 2017, 12, e0187259. [Google Scholar] [CrossRef] [PubMed]
- Crocker, J.; Tsai, A.; Stern, D.L. A fully aynthetic transcriptional platform for a multicellular eukaryote. Cell Rep. 2017, 18, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Gokhale, C.S.; Traulsen, A. Evolutionary games in the multiverse. Proc. Natl. Acad. Sci. USA 2010, 107, 5500–5504. [Google Scholar] [CrossRef] [PubMed]
- Li, X.X.; Yang, T.; Mbadinga, S.M.; Liu, J.F.; Yang, S.Z.; Gu, J.D.; Mu, B.Z. Responses of microbial community composition to temperature gradient and carbon steel corrosion in production water of petroleum reservoir. Front. Microbiol. 2017, 8, 2379. [Google Scholar] [CrossRef] [PubMed]
- M, V. The new world screwworm in Mexico and Central America. World Anim. Rev. 1991, 2835. [Google Scholar]
- Dodd, A. The control and eradication of prickly-pear in Australia. Bull. Entomol. Res. 1936, 27, 503–517. [Google Scholar] [CrossRef]
- Enkerlin, W. Impact of Fruit Fly Control Programmes Using the Sterile Insect Technique; Springer: Dordrecht, The Netherlands, 2005. [Google Scholar]
- Painter, L.E.; Beschta, R.L.; Larsen, E.J.; Ripple, W.J. Recovering aspen follow changing elk dynamics in Yellowstone: Evidence of a trophic cascade? Ecology 2015, 96, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Dobson, A.P. Yellowstone wolves and the forces that structure natural systems. PLoS Biol. 2014, 12, e1002025. [Google Scholar] [CrossRef] [PubMed]
- Rands, M.R.W.; Adams, W.M.; Bennun, L.; Butchart, S.H.M.; Clements, A.; Coomes, D.; Entwistle, A.; Hodge, I.; Kapos, V.; Scharlemann, J.P.W.; et al. Biodiversity conservation: Challenges beyond 2010. Science 2010, 329, 1298–1303. [Google Scholar] [CrossRef] [PubMed]
- Esvelt, K.M.; Gemmell, N.J. Conservation demands safe gene drive. PLoS Biol. 2017, 15, e2003850. [Google Scholar] [CrossRef] [PubMed]
- Jaenisch, R.; Mintz, B. Simian virus 40 DNA sequences in DNA of healthy adult mice derived from preimplantation blastocysts injected with viral DNA. Proc. Natl. Acad. Sci. USA 1974, 71, 1250–1254. [Google Scholar] [CrossRef] [PubMed]
- Fraley, R.T.; Rogers, S.G.; Horsch, R.B.; Sanders, P.R.; Flick, J.S.; Adams, S.P.; Bittner, M.L.; Brand, L.A.; Fink, C.L.; Fry, J.S.; et al. Expression of bacterial genes in plant cells. Proc. Natl. Acad. Sci. USA 1983, 80, 4803–4807. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA—Guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Champer, J.; Buchman, A.; Akbari, O.S. Cheating evolution: engineering gene drives to manipulate the fate of wild populations. Nat. Rev. Genet. 2016, 17, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Noble, C.; Olejarz, J.; Esvelt, K.M.; Church, G.M.; Nowak, M.A. Evolutionary dynamics of CRISPR gene drives. Sci. Adv. 2017, 3, e1601964. [Google Scholar] [CrossRef] [PubMed]
- Reeves, R.G.; Bryk, J.; Altrock, P.M.; Denton, J.A.; Reed, F.A. First steps towards underdominant genetic transformation of insect populations. PLoS ONE 2014, 9, e97557. [Google Scholar] [CrossRef] [PubMed]
- Altrock, P.M.; Traulsen, A.; Reeves, R.G.; Reed, F.A. Using underdominance to bi-stably transform local populations. J. Theor. Biol. 2010, 267, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Dhole, S.; Vella, M.R.; Lloyd, A.L.; Gould, F. Invasion and migration of spatially self-limiting gene drives: A comparative analysis. Evol Appl. 2018, 11, 794–808. [Google Scholar] [CrossRef] [PubMed]
- Reed, F.A.; Aquino-Michaels, T.G.; Costantini, M.S.; Láruson, Á.J.; Sutton, J.T. RPM-Drive: A robust, safe, and reversible gene drive system that remains functional after 200+ generations. arXiv, 2018; arXiv:1806.05304. [Google Scholar]
- Wunderlich, S.; Gatto, K.A. Consumer perception of genetically modified organisms and sources of information. Adv. Nutr. 2015, 6, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Waltz, E. First genetically engineered salmon sold in Canada. Nature News, 4 August 2017. [Google Scholar]
- Phuc, H.K.; Andreasen, M.H.; Burton, R.S.; Vass, C.; Epton, M.J.; Pape, G.; Fu, G.; Condon, K.C.; Scaife, S.; Donnelly, C.A.; et al. Late-acting dominant lethal genetic systems and mosquito control. BMC Biol. 2007, 5, 11. [Google Scholar] [CrossRef] [PubMed]
- Reeves, R.G.; Denton, J.A.; Santucci, F.; Bryk, J.; Reed, F.A. Scientific standards and the regulation of genetically modified insects. PLoS Negl. Trop. Dis. 2012, 6, e1502. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, G.; Ehrlich, P.R.; Dirzo, R. Biological annihilation via the ongoing sixth mass extinction signaled by vertebrate population losses and declines. Proc. Natl. Acad. Sci. USA 2017, 114, E6089–E6096. [Google Scholar] [CrossRef] [PubMed]
- Harari, Y.N. Sapiens: A Brief History of Humankind; Harper Perennial: New York, NY, USA, 2018. [Google Scholar]
- Barnosky, A.D.; Hadly, E.A.; Bascompte, J.; Berlow, E.L.; Brown, J.H.; Fortelius, M.; Getz, W.M.; Harte, J.; Hastings, A.; Marquet, P.A.; et al. Approaching a state shift in Earth’s biosphere. Nature 2012, 486, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Rogers, C.; Oldroyd, G.E.D. Synthetic biology approaches to engineering the nitrogen symbiosis in cereals. J. Exp. Bot. 2014, 65, 1939–1946. [Google Scholar] [CrossRef] [PubMed]
- Steinle, L.; Knittel, K.; Felber, N.; Casalino, C.; de Lange, G.; Tessarolo, C.; Stadnitskaia, A.; Sinninghe Damsté, J.S.; Zopfi, J.; Lehmann, M.F.; et al. Life on the edge: Active microbial communities in the Kryos MgCl2-brine basin at very low water activity. ISME J. 2018, 12, 1414–1426. [Google Scholar] [CrossRef] [PubMed]
- Ruff, S.W.; Farmer, J.D. Silica deposits on Mars with features resembling hot spring biosignatures at El Tatio in Chile. Nat. Commun. 2016, 7, 13554. [Google Scholar] [CrossRef] [PubMed]
- Schulze-Makuch, D.; Wagner, D.; Kounaves, S.P.; Mangelsdorf, K.; Devine, K.G.; de Vera, J.P.; Schmitt-Kopplin, P.; Grossart, H.P.; Parro, V.; Kaupenjohann, M.; et al. Transitory microbial habitat in the hyperarid Atacama Desert. Proc. Natl. Acad. Sci. USA 2018, 115, 2670–2675. [Google Scholar] [CrossRef] [PubMed]
- Des Marais, D.J.; Harwit, M.O.; Jucks, K.W.; Kasting, J.F.; Lin, D.N.C.; Lunine, J.I.; Schneider, J.; Seager, S.; Traub, W.A.; Woolf, N.J. Remote sensing of planetary properties and biosignatures on extrasolar terrestrial planets. Astrobiology 2002, 2, 153–181. [Google Scholar] [CrossRef] [PubMed]
- Friedmann, E.I.; Ocampo-Friedmann, R. A primitive cyanobacterium as pioneer microorganism for terraforming Mars. Adv. Space Res. 1995, 15, 243–246. [Google Scholar] [CrossRef]
- Myers, J.H. What can we learn from biological control failures? In Proceedings of the X International Symposium on Biological Control of Weeds, Bozeman, MT, USA, 4–14 July 2000; pp. 151–154. [Google Scholar]
- Fredrickson, J.K. Ecological communities by design. Science 2015, 348, 1425–1427. [Google Scholar] [CrossRef] [PubMed]
- Dolinšek, J.; Goldschmidt, F.; Johnson, D.R. Synthetic microbial ecology and the dynamic interplay between microbial genotypes. FEMS Microbiol. Rev. 2016, 40, 961–979. [Google Scholar] [CrossRef] [PubMed]
- Cira, N.J.; Pearce, M.T.; Quake, S.R. Neutral and selective dynamics in a synthetic microbial community. Proc. Natl. Acad. Sci. USA 2018, 115, E9842–E9848. [Google Scholar] [CrossRef] [PubMed]
- BioBricks Foundation, 77 Van Ness Ave, Ste 101-1626 San Francisco, CA 94102-6041. Available online: https://biobricks.org (accessed on 23 January 2019).
- Solé, R.V.; Montañez, R.; Duran-Nebreda, S.; Rodriguez-Amor, D.; Vidiella, B.; Sardanyés, J. Population dynamics of synthetic terraformation motifs. R. Soc. Open Sci. 2018, 5, 180121. [Google Scholar] [CrossRef] [PubMed]
- McKay, C.P.; Toon, O.B.; Kasting, J.F. Making Mars habitable. Nature 1991, 352, 489–496. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Species | Theoretical Analysis | Citation |
|---|---|---|
| S. cerevisiae | Minimal birth death growth model | Shou et al. [41] |
| Spatial frequency-dependent selection model | Müller et al. [42] | |
| including drift and explicit nutrient dynamics | ||
| Diffusion dynamics and individual-based simulation | Momeni et al. [50] based on Shou et al. [41] | |
| and explicit nutrient dynamics and environment | ||
| ODE and hybrid dynamical systems | Denton and Gokhale [51] based on Shou et al. [41] | |
| ODE and stability analysis | Hoek et al. [14] based on Müller et al. [42] | |
| E. coli | Monod type growth kinetics based model | Hosoda et al. [43] |
| ODE-based model with Monod kinetics | Kerner et al. [52] | |
| ODE-based population dynamics model | Amor et al. [49] based on Hosoda et al. [43] | |
| akin to hypercycles [53] but including diffusion dynamics | ||
| E. coli—A. baylyi | No theory | Santala et al. [54] |
| Individual-based model | Pande et al. [44] | |
| E. coli—D. discoideum | ODE-based population dynamics model | Kubo et al. [36] |
| L. lactis | ODE-based population dynamics with explicit | Kong et al. [45] |
| signaling molecules and nutrient dynamics and varied | ||
| ecological interactions |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Denton, J.A.; Gokhale, C.S. Synthetic Mutualism and the Intervention Dilemma. Life 2019, 9, 15. https://doi.org/10.3390/life9010015
Denton JA, Gokhale CS. Synthetic Mutualism and the Intervention Dilemma. Life. 2019; 9(1):15. https://doi.org/10.3390/life9010015
Chicago/Turabian StyleDenton, Jai A., and Chaitanya S. Gokhale. 2019. "Synthetic Mutualism and the Intervention Dilemma" Life 9, no. 1: 15. https://doi.org/10.3390/life9010015
APA StyleDenton, J. A., & Gokhale, C. S. (2019). Synthetic Mutualism and the Intervention Dilemma. Life, 9(1), 15. https://doi.org/10.3390/life9010015