The Impact of Salts on Single Chain Amphiphile Membranes and Implications for the Location of the Origin of Life

Abstract
1. Early Earth Environments for the Origins of Life
2. Membranes and Life
3. Salt and Membranes
4. Other Environmental Factors
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Knoll, A.H.; Barghoorn, E.S. Archean microfossils showing cell division from the swaziland system of South Africa. Science 1977, 198, 396–398. [Google Scholar] [CrossRef] [PubMed]
- Wacey, D.; Kilburn, M.R.; Saunders, M.; Cliff, J.; Brasier, M.D. Microfossils of sulphur-metabolizing cells in 3.4-billion-year-old rocks of Western Australia. Nat. Geosci. 2011, 4, 698–702. [Google Scholar] [CrossRef]
- Special Issue: Hydrothermal Vents or Hydrothermal Fields: Challenging Paradigms. Life 2017. Available online: https://www.mdpi.com/journal/life/special_issues/hydrothermal (accessed on 15 September 2017).
- Holm, N.G. Why are hydrothermal systems proposed as plausible environments for the origin of life? Orig. Life Evol. Biosph. 1992, 22, 5–14. [Google Scholar] [CrossRef]
- Sojo, V.; Herschy, B.; Whicher, A.; Camprubi, E.; Lane, N. The origin of life in alkaline hydrothermal vents. Astrobiology 2016, 16, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Chyba, C.F. The violent environment of the origin of life–progress and uncertainties. Geochim. Cosmochim. Acta 1993, 57, 3351–3358. [Google Scholar] [CrossRef]
- Simoneit, B.R.T. Prebiotic organic synthesis under hydrothermal conditions: An overview. Adv. Sp. Res. 2004, 33, 88–94. [Google Scholar] [CrossRef]
- McCollom, T.M.; Simoneit, B.R.T. Abiotic formation of hydrocarbons and oxygenated compounds during thermal decomposition of iron oxalate. Orig. Life Evol. Biosph. 1999, 29, 167–186. [Google Scholar] [CrossRef] [PubMed]
- McCollom, T.M.; Ritter, G.; Simoneit, B.R.T. Lipid synthesis under hydrothermal conditions by Fischer-Tropsch-type reactions. Orig. Life Evol. Biosph. 1999, 29, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Damer, B.; Deamer, D. Coupled phases and combinatorial selection in fluctuating hydrothermal pools: A scenario to guide experimental approaches to the origin of cellular life. Life 2015, 5, 872–887. [Google Scholar] [CrossRef] [PubMed]
- Damer, B. A field trip to the archaean in search of darwin’s warm little pond. Life 2016, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Rajamani, S.; Vlassov, A.; Benner, S.; Coombs, A.; Olasagasti, F.; Deamer, D. Lipid-assisted synthesis of RNA-like polymers from mononucleotides. Orig. Life Evol. Biosph. 2008, 38, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Garcia, M.; Surman, A.J.; Cooper, G.J.T.; Suárez-Marina, I.; Hosni, Z.; Lee, M.P.; Cronin, L. Formation of oligopeptides in high yield under simple programmable conditions. Nat. Commun. 2015, 6, 8385. [Google Scholar] [CrossRef] [PubMed]
- Monnard, P.A.; Apel, C.L.; Kanavarioti, A.; Deamer, D.W. Influence of ionic inorganic solutes on self-assembly and polymerization processes related to early forms of life: Implications for a prebiotic aqueous medium. Astrobiology 2004, 2, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Bedau, M.A.; McCaskill, J.S.; Packard, N.H.; Parke, E.C.; Rasmussen, S.R. Introduction to recent developments in living technology. Artif. Life 2013, 19, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.; Constantinescu, A.; Svaneborg, C. Generating minimal living systems from non-living materials and increasing their evolutionary abilities. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150440. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.; Chen, L.H.; Deamer, D.; Krakauer, D.C.; Packard, N.H.; Stadler, P.F.; Bedau, M.A. Transitions from nonliving to living matter. Science 2004, 303, 963–965. [Google Scholar] [CrossRef] [PubMed]
- Bedau, M.A.; McCaskill, J.S.; Packard, N.H.; Rasmussen, S. Living technology: Exploiting life’s principles in technology. Artif. Life 2010, 16, 9. [Google Scholar] [CrossRef] [PubMed]
- Mansy, S.S.; Schrum, J.P.; Krishnamurthy, M.; Tobe, S.; Treco, D.A.; Szostak, J.W. Template-directed synthesis of a genetic polymer in a model protocell. Nature 2008, 454, 122–125. [Google Scholar] [CrossRef] [PubMed]
- Adamala, K.P.; Engelhart, A.E.; Szostak, J.W. Collaboration between primitive cell membranes and soluble catalysts. Nat. Commun. 2016, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.A.; Salehi-Ashtiani, K.; Szostak, J.W. RNA catalysis in model protocell vesicles. J. Am. Chem. Soc. 2005, 127, 13213–13219. [Google Scholar] [CrossRef] [PubMed]
- Cape, J.L.; Monnard, P.A.; Boncella, J.M. Prebiotically relevant mixed fatty acid vesicles support anionic solute encapsulation and photochemically catalyzed trans-membrane charge transport. Chem. Sci. 2011, 2, 661–671. [Google Scholar] [CrossRef]
- Maurer, S.E.; DeClue, M.S.; Albertsen, A.N.; Dorr, M.; Kuiper, D.S.; Ziock, H.; Rasmussen, S.; Boncella, J.M.; Monnard, P.A. Interactions between catalysts and amphiphilic structures and their implications for a protocell model. ChemPhysChem 2011, 12, 828–835. [Google Scholar] [CrossRef] [PubMed]
- Volkov, G.; Gugeshashvili, M.I.; Deamer, D.W. Energy conversion at liquid/liquid interfaces: Artificial photosynthetic systems. Electrochim. Acta 1995, 40, 2849–2868. [Google Scholar] [CrossRef]
- Deamer, D.W. Polycyclic aromatic hydrocarbons: Primitive pigment systems in the prebiotic environment. Adv. Sp. Res. 1992, 12, 183–189. [Google Scholar] [CrossRef]
- Zhu, T.F.; Adamala, K.; Zhang, N.; Szostak, J.W. Photochemically driven redox chemistry induces protocell membrane pearling and division. Proc. Natl. Acad. Sci. USA 2012, 109, 9828–9832. [Google Scholar] [CrossRef] [PubMed]
- Hanczyc, M.M.; Szostak, J.W. Replicating vesicles as models of primitive cell growth and division. Curr. Opin. Chem. Biol. 2004, 8, 660–664. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.A.; Szostak, J.W. Membrane growth can generate a transmembrane pH gradient in fatty acid vesicles. Proc. Natl. Acad. Sci. USA 2004, 101, 7965–7970. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.A.; Szostak, J.W. A kinetic study of the growth of fatty acid vesicles. Biophys. J. 2004, 87, 988–998. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.F.; Szostak, J.W. Coupled growth and division of model protocell membranes. J. Am. Chem. Soc. 2009, 131, 5705–5713. [Google Scholar] [CrossRef] [PubMed]
- Gebicki, J.M.; Hicks, M. Ufasomes are stable particles surrounded by unsaturated fatty-acid membranes. Nature 1973, 243, 232–234. [Google Scholar] [CrossRef] [PubMed]
- Rabenau, A. The role of hydrothermal synthesis in preparative chemistry. Angew. Chem. Int. Ed. Engl. 1985, 24, 1026–1040. [Google Scholar] [CrossRef]
- Simoneit, B.R.T.; Rushdi, A.I.; Deamer, D.W. Abiotic formation of acylglycerols under simulated hydrothermal conditions and self-assembly properties of such lipid products. Adv. Sp. Res. 2007, 40, 1649–1656. [Google Scholar] [CrossRef]
- Dworkin, L.P.; Deamer, D.W.; Sandford, S.A.; Allamandola, L.J. Self-Assembling amphiphilic molecules: Synthesis in simulated interstellar/precometary ices. Proc. Natl. Acad. Sci. USA 2001, 98, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Deamer, D.W. Boundary structures are formed by organic-components of the murchison carbonaceous chondrite. Nature 1985, 317, 792–794. [Google Scholar] [CrossRef]
- Hargreaves, W.R.; Deamer, D.W. Liposomes from ionic, single-chain amphiphiles. Biochemistry 1978, 17, 3759–3768. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, W.R.; Mulvihill, S.J.; Deamer, D.W. Synthesis of phospholipids and membranes in prebiotic conditions. Nature 1977, 266, 78–80. [Google Scholar] [CrossRef] [PubMed]
- Dalai, P.; Kaddour, H.; Sahai, N. Incubating life: Prebiotic sources of organics for the origin of life. Elements 2016, 12, 401–406. [Google Scholar] [CrossRef]
- Maurer, S.E.; Nguyen, G. Prebiotic vesicle formation and the necessity of salts. Orig. Life Evol. Biosph. 2016, 46, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.; Tanford, C. Hydrophobicity of long chain n-Alkyl carboxylic acids, as measured by their distribution between heptane and aqueous solutions. Proc. Natl. Acad. Sci. USA 1973, 70, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Knauth, L.P. Temperature and salinity history of Precambrian ocean: Implications for the course of microbial evolution. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 219, 53–69. [Google Scholar] [CrossRef]
- Theillet, F.X.; Binolfi, A.; Frembgen-Kesner, T.; Hingorani, K.; Sarkar, M.; Kyne, C.; Li, C.; Crowley, P.B.; Gierasch, L.; Pielak, G.J.; et al. Physicochemical properties of cells and their effects on intrinsically disordered proteins (IDPs). Chem. Rev. 2014, 114, 6661–6714. [Google Scholar] [CrossRef] [PubMed]
- Lodish, H.F. Molecular Cell Biology, 6th ed.; W.H. Freeman: New York, NY, USA, 2008. [Google Scholar]
- Huang, X.; Sillanpää, M.; Gjessing, E.T.; Vogt, R.D. Water quality in the Tibetan Plateau: Major ions and trace elements in the headwaters of four major Asian rivers. Sci. Total Environ. 2009, 407, 6242–6254. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Tan, X.; Li, T.; Chen, Y.; Huang, X. Influence of salt and polymer on the critical vesicle concentration in aqueous mixture of zwitterionic/anionic surfactants. Colloids Surf. A Physicochem. Eng. Asp. 2006, 276, 28–33. [Google Scholar] [CrossRef]
- Monnard, P.A.; Deamer, D.W. Preparation of vesicles from nonphospholipid amphiphiles. Liposomes 2003, 372 Pt B, 133–151. [Google Scholar]
- Tanford, C. The Hydrophobic Effect: Formation of Micelles and Biological Membranes, 2nd ed.; J. Wiley: New York, NY, USA, 1980. [Google Scholar]
- Maurer, S.E.; Deamer, D.W.; Boncella, J.M.; Monnard, P.A. Chemical evolution of amphiphiles: Glycerol monoacyl derivatives stabilize plausible prebiotic membranes. Astrobiology 2009, 9, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Albertsen, A.N.; Duffy, C.D.; Sutherland, J.D.; Monnard, P.A. Self-Assembly of phosphate amphiphiles in mixtures of prebiotically plausible surfactants. Astrobiology 2014, 14, 462–472. [Google Scholar] [CrossRef] [PubMed]
- Caschera, F.; de la Serna, J.B.; Loffler, P.M.G.; Rasmussen, T.E.; Hanczyc, M.M.; Bagatolli, L.A.; Monnard, P.A. Stable vesicles composed of monocarboxylic or dicarboxylic fatty acids and trimethylammonium amphiphiles. Langmuir 2011, 27, 14078–14090. [Google Scholar] [CrossRef] [PubMed]
- Namani, T.; Deamer, D.W. Stability of model membranes in extreme environments. Orig. Life Evol. Biosph. 2008, 38, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Apel, C.L.; Deamer, D.W.; Mautner, M.N. Self-assembled vesicles of monocarboxylic acids and alcohols: conditions for stability and for the encapsulation of biopolymers. Biochim. Biophys. Acta Biomembr. 2002, 1559, 1–9. [Google Scholar] [CrossRef]
- Gotoh, M.; Miki, A.; Nagano, H.; Ribeiro, N.; Elhabiri, M.; Gumienna-Kontecka, E.; Albrecht-Gary, A.; Schmutz, M.; Ourisson, G.; Nakatani, Y. Membrane properties of branched polyprenyl phosphates, postulated as primitive membrane constituents. Chem. Biodivers. 2006, 3, 434–455. [Google Scholar] [CrossRef] [PubMed]
- Fiore, M.; Madanamoothoo, W.; Berlioz-Barbier, A.; Maniti, O.; Girard-Egrot, A.; Buchet, R.; Strazewski, P. Giant vesicles from rehydrated crude mixtures containing unexpected mixtures of amphiphiles formed under plausibly prebiotic conditions. Org. Biomol. Chem. 2017, 15, 4231–4240. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Du, N.; Song, Y.; Song, S.; Hou, W. Vesicles of 2-ketooctanoic acid in water. Soft Matter 2017, 13, 2246–2252. [Google Scholar] [CrossRef] [PubMed]
- Maurer, S.E.; Monnard, P.A.; Hanczyc, M.M. Vesicle self-assembly of monoalkyl amphiphiles under hydrothermal vent-like conditions: Effects of high ionic strength, extreme pH and high temperature environments. Langmuir 2017. in preparation. [Google Scholar]
- Piedrafita, G.; Montero, F.; Morán, F.; Cárdenas, M.L.; Cornish-Bowden, A. A Simple self-maintaining metabolic system: Robustness, autocatalysis, bistability. PLoS Comput. Biol. 2010, 6, e1000872. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.; Baross, J.; Kelley, D.; Russell, M.J. Hydrothermal vents and the origin of life. Nat. Rev. Microbiol. 2008, 6, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.S.; Karson, J.A.; Blackman, D.K.; Früh-Green, G.L.; Butterfield, D.A.; Lilley, M.D.; Olson, E.J.; Schrenk, M.O.; Roe, K.K.; Lebon, G.T.; et al. An off-axis hydrothermal vent field near the Mid-Atlantic Ridge at 30 degrees N. Nature 2001, 412, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Walde, P.; Wessicken, M.; Rädler, U.; Berclaz, N.; Conde-Frieboes, K.; Luisi, P.L. Preparation and characterization of vesicles from mono-n-alkyl phosphates and phosphonates. J. Phys. Chem. B 1997, 101, 8. [Google Scholar] [CrossRef]
- Lauerwald, R.; Hartmann, J.; Moosdorf, N.; Kempe, S.; Raymond, P.A. What controls the spatial patterns of the riverine carbonate system? A case study for North America. Chem. Geol. 2013, 337–338, 114–127. [Google Scholar] [CrossRef]
- Koynova, R.; Caffrey, M. Phases and phase transitions of the phosphatidylcholines. Biochim. Biophys. Acta Rev. Biomembr. 1998, 1376, 91–145. [Google Scholar] [CrossRef]
- Mayer, C.; Schreiber, U.; Dávila, M.J. Periodic vesicle formation in tectonic fault zones: An ideal scenario for molecular evolution. Orig. Life Evol. Biosph. 2015, 45, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Groen, J.; Deamer, D.W.; Kros, A.; Ehrenfreund, P. Polycyclic aromatic hydrocarbons as plausible prebiotic membrane components. Orig. Life Evol. Biosph. 2012, 42, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Oleson, T.A.; Sahai, N. Oxide-Dependent adsorption of a model membrane phospholipid dipalmitoylphosphatidylcholine: Bulk adsorption isotherms. Langmuir 2008, 24, 4865–4873. [Google Scholar] [CrossRef] [PubMed]
- Hanczyc, M.M.; Mansy, S.S.; Szostak, J.W. Mineral surface directed membrane assembly. Orig. Life Evol. Biosph. 2007, 37, 67–82. [Google Scholar] [CrossRef] [PubMed]




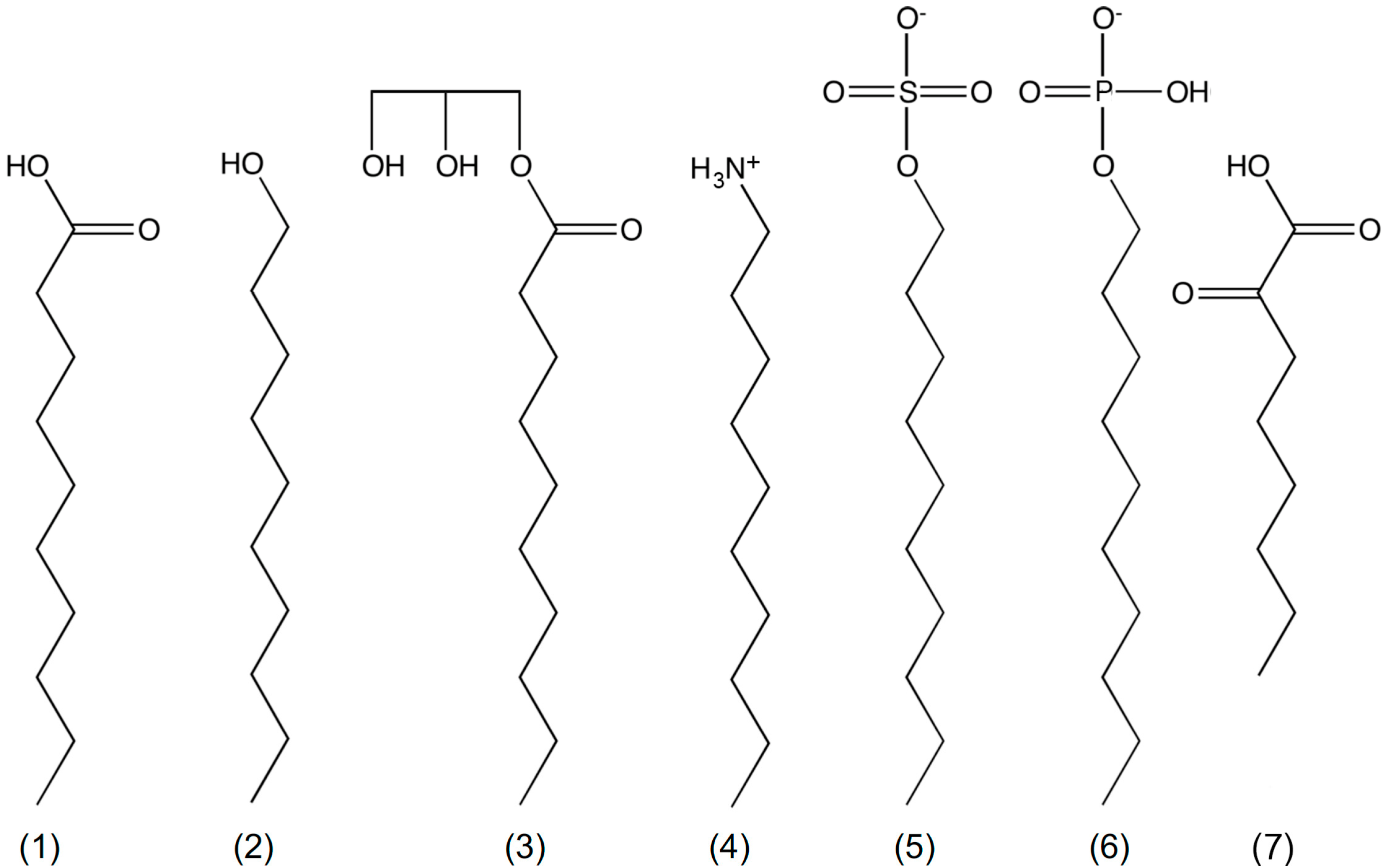{kind=link}
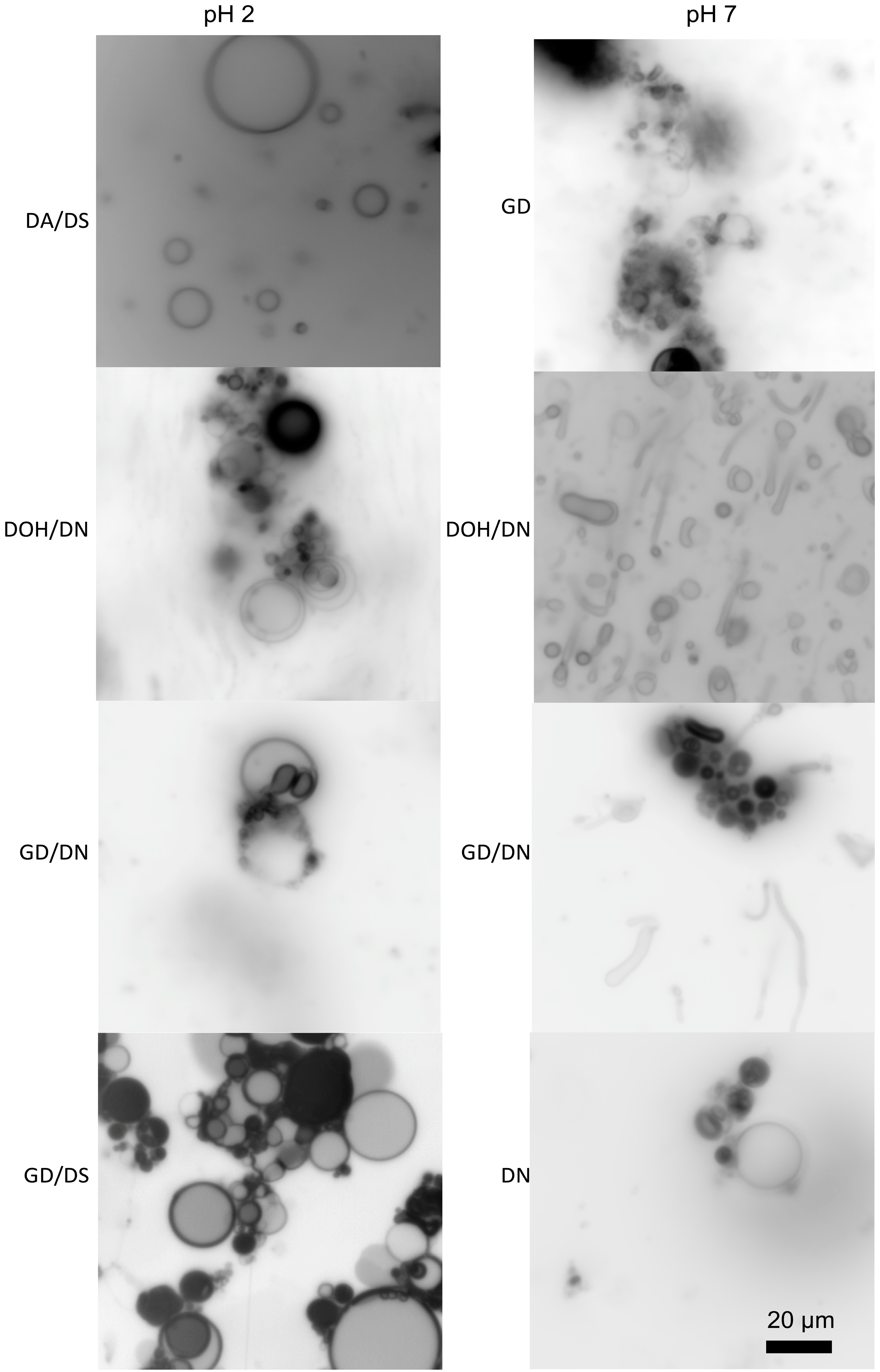{kind=link}
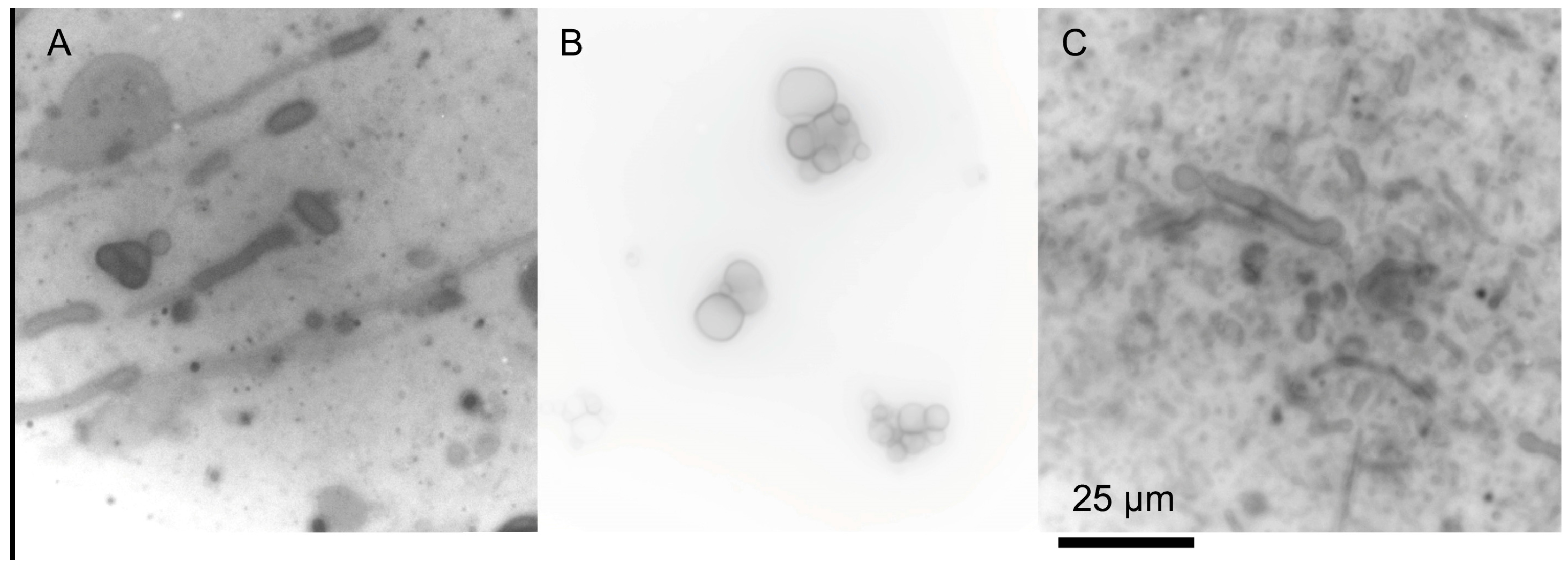{kind=link}
| Ion | Cytosol (mM) [42] a | Cytosol (mM) [43] b | Blood (mM) [43] b | River Water (mM) [44] | Seawater (mM) [14] |
|---|---|---|---|---|---|
| Potassium | 200–250 | 139 | 4 | 0.051 | 10 |
| Sodium | 5 | 12 | 145 | 0.261 | 400 |
| Chloride | 6 | 4 | 116 | 0.226 | 460 |
| Bicarbonate | 12 | 29 | 0.951 | 2.1 | |
| Magnesium(free) | 1–2 | 0.8 | 1.5 | 0.165 | 50 |
| Calcium (free) | 0.1 | <0.0002 | 1.8 | 0.374 | 10 |
| Sulfate | 0.115 | 20 | |||
| Other | 0.233 | 0.5 |
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maurer, S. The Impact of Salts on Single Chain Amphiphile Membranes and Implications for the Location of the Origin of Life. Life 2017, 7, 44. https://doi.org/10.3390/life7040044
Maurer S. The Impact of Salts on Single Chain Amphiphile Membranes and Implications for the Location of the Origin of Life. Life. 2017; 7(4):44. https://doi.org/10.3390/life7040044
Chicago/Turabian StyleMaurer, Sarah. 2017. "The Impact of Salts on Single Chain Amphiphile Membranes and Implications for the Location of the Origin of Life" Life 7, no. 4: 44. https://doi.org/10.3390/life7040044
APA StyleMaurer, S. (2017). The Impact of Salts on Single Chain Amphiphile Membranes and Implications for the Location of the Origin of Life. Life, 7(4), 44. https://doi.org/10.3390/life7040044