Stem Cells toward the Future: The Space Challenge

{kind=link}
Abstract
:1. Introduction
2. Considerations Concerning Approaches and Models
2.1. Microgravity Simulators
2.2. Experimental Model Organisms
2.3. Osteogenic Media
2.4. Oxygen Tension (pO2)
3.1. Experiments in Simulated Microgravity
3.2. Experiments in Space
4. Mechanobiology
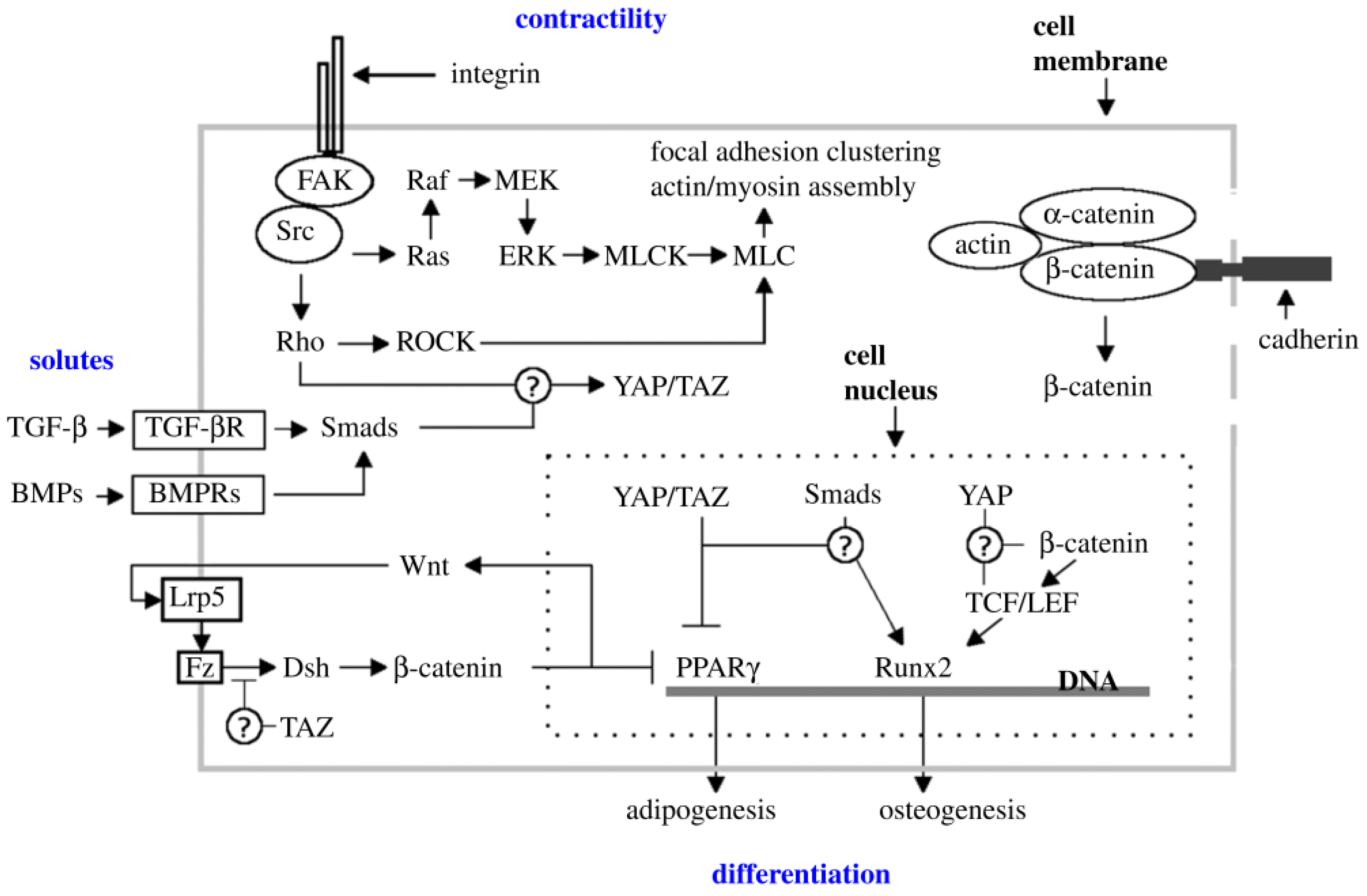
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Orwoll, E.S.; Adler, R.A.; Amin, S.; Binkley, N.; Lewiecki, E.M.; Petak, S.M.; Shapses, S.A.; Sinaki, M.; Watts, N.B.; Sibonga, J.D. Skeletal health in long-duration astronauts: Nature, assessment, and management recommendations from the NASA Bone Summit. J. Bone Miner. Res. 2013, 28, 1243–1255. [Google Scholar] [CrossRef]
- Nagaraja, M.P.; Risin, D. The current state of bone loss research: Data from spaceflight and microgravity simulators. J. Cell. Biochem. 2013, 114, 1001–1008. [Google Scholar] [CrossRef]
- Blaber, E.A.; Dvorochkin, N.; Lee, C.; Alwood, J.S.; Yousuf, R.; Pianetta, P.; Globus, R.K.; Burns, B.P.; Almeida, E.A. Microgravity induces pelvic bone loss through osteoclastic activity, osteocytic osteolysis, and osteoblastic cell cycle inhibition by CDKN1a/p21. PLoS One 2013, 8, e61372. [Google Scholar]
- Zayzafoon, M.; Gathings, W.E.; McDonald, J.M. Modeled microgravity inhibits osteogenic differentiation of human mesenchymal stem cells and increases adipogenesis. Endocrinology 2004, 145, 2421–2432. [Google Scholar] [CrossRef]
- Monticone, M.; Liu, Y.; Pujic, N.; Cancedda, R. Activation of nervous system development genes in bone marrow derived mesenchymal stem cells following spaceflight exposure. J. Cell. Biochem. 2010, 111, 442–452. [Google Scholar] [CrossRef]
- Versari, S.; Klein-Nulend, J.; van Loon, J.; Bradamante, S. Influence of Oxygen in the Cultivation of Human Mesenchymal Stem Cells in Simulated Microgravity: An Explorative Study. Microgravity Sci. Technol. 2013, 25, 59–66. [Google Scholar] [CrossRef]
- Wang, N.; Wang, H.; Chen, J.; Zhang, X.; Xie, J.; Li, Z.; Ma, J.; Wang, W.; Wang, Z. The simulated microgravity enhances multipotential differentiation capacity of bone marrow mesenchymal stem cells. Cytotechnology 2013, 66, 119–131. [Google Scholar]
- Buravkova, L.B.; Gershovich, P.M.; Gershovich, J.G.; Grigoriev, A.I. Microgravity and Mesenchymal Stem Cell Response. Curr. Biotechnol. 2013, 2, 217–225. [Google Scholar] [CrossRef]
- Bianco, P.; Cao, X.; Frenette, P.S.; Mao, J.J.; Robey, P.G.; Simmons, P.J.; Wang, C.Y. The meaning, the sense and the significance: Translating the science of mesenchymal stem cells into medicine. Nat. Med. 2013, 19, 35–42. [Google Scholar]
- Gershovich, P.M.; Gershovich Iu, G.; Buravkova, L.B. Cytoskeleton structures and adhesion properties of human stromal precursors under conditions of simulated microgravity. Tsitologiia 2009, 51, 896–904. [Google Scholar]
- Ehninger, A.; Trumpp, A. The bone marrow stem cell niche grows up: Mesenchymal stem cells and macrophages move in. J. Exp. Med. 2011, 208, 421–428. [Google Scholar] [CrossRef]
- Nombela-Arrieta, C.; Ritz, J.; Silberstein, L.E. The elusive nature and function of mesenchymal stem cells. Nat. Rev. Mol. Cell Biol. 2011, 12, 126–131. [Google Scholar] [CrossRef]
- Frenette, P.S.; Pinho, S.; Lucas, D.; Scheiermann, C. Mesenchymal Stem Cell: Keystone of the Hematopoietic Stem Cell Niche and a Stepping-Stone for Regenerative Medicine. Annu. Rev. Immunol. 2013, 31, 285–316. [Google Scholar] [CrossRef]
- Morrison, S.J.; Scadden, D.T. The bone marrow niche for haematopoietic stem cells. Nature 2014, 505, 327–334. [Google Scholar] [CrossRef]
- DuFort, C.C.; Paszek, M.J.; Weaver, V.M. Balancing forces: Architectural control of mechanotransduction. Nat. Rev. Mol. Cell Biol. 2011, 12, 308–319. [Google Scholar] [CrossRef]
- Hoffman, B.D.; Grashoff, C.; Schwartz, M.A. Dynamic molecular processes mediate cellular mechanotransduction. Nature 2011, 475, 316–323. [Google Scholar] [CrossRef]
- Jaalouk, D.E.; Lammerding, J. Mechanotransduction gone awry. Nat. Rev. Mol. Cell Biol. 2009, 10, 63–73. [Google Scholar] [CrossRef]
- Janmey, P.A.; Miller, R.T. Mechanisms of mechanical signaling in development and disease. J. Cell Sci. 2011, 124, 9–18. [Google Scholar] [CrossRef]
- Sukharev, S.; Sachs, F. Molecular force transduction by ion channels: Diversity and unifying principles. J. Cell Sci. 2012, 125, 3075–3083. [Google Scholar] [CrossRef]
- Huang, C.; Ogawa, R. Mechanotransduction in bone repair and regeneration. FASEB J. 2010, 24, 3625–3632. [Google Scholar] [CrossRef]
- Ingber, D.E. Tensegrity II. How structural networks influence cellular information processing networks. J. Cell Sci. 2003, 116, 1397–1408. [Google Scholar] [CrossRef]
- Ingber, D.E. Tensegrity-based mechanosensing from macro to micro. Prog. Biophys. Mol. Biol. 2008, 97, 163–179. [Google Scholar]
- Pavalko, F.M.; Norvell, S.M.; Burr, D.B.; Turner, C.H.; Duncan, R.L.; Bidwell, J.P. A model for mechanotransduction in bone cells: The load-bearing mechanosomes. J. Cell. Biochem. 2003, 88, 104–112. [Google Scholar] [CrossRef]
- Hughes-Fulford, M. Function of the cytoskeleton in gravisensing during spaceflight. Adv. Space Res. 2003, 32, 1585–1593. [Google Scholar] [CrossRef]
- Walther, I.; Bechler, B.; Muller, O.; Hunzinger, E.; Cogoli, A. Cultivation of Saccharomyces cerevisiae in a bioreactor in microgravity. J. Biotechnol. 1996, 47, 113–127. [Google Scholar] [CrossRef]
- Vorselen, D.; Roos, W.H.; Mackintosh, F.C.; Wuite, G.J.; van Loon, J.J. The role of the cytoskeleton in sensing changes in gravity by nonspecialized cells. FASEB J. 2014, 28, 536–547. [Google Scholar] [CrossRef]
- Karp, J.M.; Leng Teo, G.S. Mesenchymal stem cell homing: The devil is in the details. Cell Stem Cell 2009, 4, 206–216. [Google Scholar] [CrossRef]
- Bianco, P. Reply to MSCs: Science and trials. Nat. Med. 2013, 19, 813–814. [Google Scholar] [CrossRef]
- Alvarez, C.V.; Garcia-Lavandeira, M.; Garcia-Rendueles, M.E.R.; Diaz-Rodriguez, E.; Garcia-Rendueles, A.R.; Perez-Romero, S.; Vila, T.V.; Rodrigues, J.S.; Lear, P.V.; Bravo, S.B. Defining stem cell types: Understanding the therapeutic potential of ESCs, ASCs, and iPS cells. J. Mol. Endocrinol. 2012, 49, R89–R111. [Google Scholar] [CrossRef]
- Herranz, R.; Anken, R.; Boonstra, J.; Braun, M.; Christianen, P.C.M.; de Geest, M.; Hauslage, J.; Hilbig, R.; Hill, R.J.A.; Lebert, M.; et al. Ground-Based Facilities for Simulation of Microgravity: Organism-Specific Recommendations for Their Use, and Recommended Terminology. Astrobiology 2013, 13, 1–17. [Google Scholar] [CrossRef]
- Liu, N.; Zang, R.; Yang, S.T.; Li, Y. Stem cell engineering in bioreactors for large-scale bioprocessing. Eng. Life Sci. 2014, 14, 4–15. [Google Scholar] [CrossRef]
- Rodrigues, C.A.V.; Fernandes, T.G.; Diogo, M.M.; da Silva, C.L.; Cabral, J.M.S. Stem cell cultivation in bioreactors. Biotechnol. Adv. 2011, 29, 815–829. [Google Scholar] [CrossRef]
- Sart, S.B.; Agathos, S.N.; Li, Y. Process Engineering of Stem Cell Metabolism for Large Scale Expansion and Differentiation in Bioreactors. Biochem. Eng. J. 2014, 84, 74–82. [Google Scholar] [CrossRef]
- Stenderup, K.; Justesen, J.; Clausen, C.; Kassem, M. Aging is associated with decreased maximal life span and accelerated senescence of bone marrow stromal cells. Bone 2003, 33, 919–926. [Google Scholar] [CrossRef]
- Arvidson, K.; Abdallah, B.M.; Applegate, L.A.; Baldini, N.; Cenni, E.; Gomez-Barrena, E.; Granchi, D.; Kassem, M.; Konttinen, Y.T.; Mustafa, K.; et al. Bone regeneration and stem cells. J. Cell. Mol. Med. 2011, 15, 718–746. [Google Scholar] [CrossRef]
- Phinney, D.G.; Kopen, G.; Isaacson, R.L.; Prockop, D.J. Plastic adherent stromal cells from the bone marrow of commonly used strains of inbred mice: Variations in yield, growth, and differentiation. J. Cell. Biochem. 1999, 72, 570–585. [Google Scholar]
- Russell, K.C.; Phinney, D.G.; Lacey, M.R.; Barrilleaux, B.L.; Meyertholen, K.E.; O’Connor, K.C. In vitro high-capacity assay to quantify the clonal heterogeneity in trilineage potential of mesenchymal stem cells reveals a complex hierarchy of lineage commitment. Stem Cells 2010, 28, 788–798. [Google Scholar] [CrossRef]
- Placzek, M.R.; Chung, I.M.; Macedo, H.M.; Ismail, S.; Mortera Blanco, T.; Lim, M.; Cha, J.M.; Fauzi, I.; Kang, Y.; Yeo, D.C.; et al. Stem cell bioprocessing: Fundamentals and principles. J. R. Soc. Interface 2009, 6, 209–232. [Google Scholar] [CrossRef]
- Carmeliet, G.; Nys, G.; Bouillon, R. Microgravity reduces the differentiation of human osteoblastic MG-63 cells. J. Bone Miner. Res. 1997, 12, 786–794. [Google Scholar] [CrossRef]
- Carmeliet, G.; Nys, G.; Stockmans, I.; Bouillon, R. Gene expression related to the differentiation of osteoblastic cells is altered by microgravity. Bone 1998, 22, 139S–143S. [Google Scholar] [CrossRef]
- Hughes-Fulford, M.; Gilbertson, V. Osteoblast fibronectin mRNA, protein synthesis, and matrix are unchanged after exposure to microgravity. FASEB J. 1999, 13 (Suppl.), S121–S127. [Google Scholar]
- Hughes-Fulford, M.; Lewis, M.L. Effects of microgravity on osteoblast growth activation. Exp. Cell Res. 1996, 224, 103–109. [Google Scholar] [CrossRef]
- Nabavi, N.; Khandani, A.; Camirand, A.; Harrison, R.E. Effects of microgravity on osteoclast bone resorption and osteoblast cytoskeletal organization and adhesion. Bone 2011, 49, 965–974. [Google Scholar] [CrossRef]
- Pardo, S.J.; Patel, M.J.; Sykes, M.C.; Platt, M.O.; Boyd, N.L.; Sorescu, G.P.; Xu, M.; van Loon, J.J.W.A.; Wang, M.D.; Jo, H. Simulated microgravity using the Random Positioning Machine inhibits differentiation and alters gene expression profiles of 2T3 preosteoblasts. Am. J. Physiol. Cell Physiol. 2005, 288, C1211–C1221. [Google Scholar] [CrossRef]
- Heng, B.C.; Cao, T.; Stanton, L.W.; Robson, P.; Olsen, B. Strategies for Directing the Differentiation of Stem Cells Into the Osteogenic Lineage in Vitro. J. Bone Miner. Res. 2004, 19, 1379–1394. [Google Scholar] [CrossRef]
- Yong-sheng, Z.; Yun-song, L.I.U.; Jian-guo, T.A.N. Is 1,25-dihydroxyvitamin D3 an ideal substitute for dexame-thasone for inducing osteogenic differentiation of human adipose tissue-derived stromal cells in vitro? Chin. Med. J. 2006, 119, 1278–1286. [Google Scholar]
- Kelly, K.A.; Gimble, J.M. 1,25-dihydroxy vitamin D-3 inhibits adipocyte differentiation and gene expression in murine bone marrow stromal cell clones and primary cultures. Endocrinology 1998, 139, 2622–2628. [Google Scholar]
- Kong, J.; Li, Y.C. Molecular mechanism of 1,25-dihydroxyvitamin D3 inhibition of adipogenesis in 3T3-L1 cells. Am. J. Physiol. Endocrinol. MeTab. 2006, 290, E916–E924. [Google Scholar] [CrossRef]
- Sakaguchi, K.; Morita, I.; Murota, S. Relationship between the ability to support differentiation of osteoclastlike cells and adipogenesis in murine stromal cells derived from bone marrow. Prostaglandins Leukot. Essent. Fat. Acids 2000, 62, 319–327. [Google Scholar] [CrossRef]
- Guo, F.M.; Dai, Z.Q.; Wu, F.; Liu, Z.X.; Tan, Y.J.; Wan, Y.M.; Shang, P.; Li, Y.H. Gravity affects the responsiveness of Runx2 to 1, 25-dihydroxyvitamin D3 (VD3). Acta Astronaut. 2013, 84, 189–196. [Google Scholar] [CrossRef]
- Oganov, V.S.; Baranov, V.S.; Kabitskaya, O.E.; Novikov, V.E.; Bakulin, A.V.; Moskalenko, M.V.; Aseev, M.V.; Voitulevich, L.V. Analysis of polymorphism of bone metabolism genes and evaluation of the risk of osteopenia in cosmonauts. Hum. Physiol. 2012, 38, 732–737. [Google Scholar] [CrossRef]
- Moore, K.A.; Lemischka, I.R. Stem cells and their niches. Science 2006, 311, 1880–1885. [Google Scholar] [CrossRef]
- Badylak, S.F.; Nerem, R.M. Progress in tissue engineering and regenerative medicine. Proc. Natl. Acad. Sci. USA 2010, 107, 3285–3286. [Google Scholar] [CrossRef]
- Mohyeldin, A.; Garzon-Muvdi, T.; Quinones-Hinojosa, A. Oxygen in stem cell biology: A critical component of the stem cell niche. Cell Stem Cell 2010, 7, 150–161. [Google Scholar] [CrossRef]
- Klaus, D.M. Clinostats and bioreactors. Gravit. Space Biol. Bull. 2001, 14, 55–64. [Google Scholar]
- Begley, C.M.; Kleis, S.J. RWPV bioreactor mass transport: Earth-based and in microgravity. Biotechnol. Bioeng. 2002, 80, 465–476. [Google Scholar] [CrossRef]
- Rivera-Solorio, I.; Kleis, S.J. Model of the mass transport to the surface of animal cells cultured in a rotating bioreactor operated in micro gravity. Biotechnol. Bioeng. 2006, 94, 495–504. [Google Scholar] [CrossRef]
- Benjamin, S.; Sheyn, D.; Ben-David, S.; Oh, A.; Kallai, I.; Li, N.; Gazit, D.; Gazit, Z. Oxygenated environment enhances both stem cell survival and osteogenic differentiation. Tissue Eng. Part A 2012, 19, 748–758. [Google Scholar]
- Nichols, H.L.; Zhang, N.; Wen, X. Proteomics and genomics of microgravity. Physiol. Genomics 2006, 26, 163–171. [Google Scholar] [CrossRef]
- Hughes-Fulford, M. To infinity … and beyond! Human spaceflight and life science. FASEB J. 2011, 25, 2858–2864. [Google Scholar] [CrossRef]
- Dai, Z.Q.; Wang, R.; Ling, S.K.; Wan, Y.M.; Li, Y.H. Simulated microgravity inhibits the proliferation and osteogenesis of rat bone marrow mesenchymal stem cells. Cell Prolif. 2007, 40, 671–684. [Google Scholar] [CrossRef]
- Huang, Y.; Dai, Z.Q.; Ling, S.K.; Zhang, H.Y.; Wan, Y.M.; Li, Y.H. Gravity, a regulation factor in the differentiation of rat bone marrow mesenchymal stem cells. J. Biomed. Sci. 2009, 16. [Google Scholar] [CrossRef]
- Meyers, V.E.; Zayzafoon, M.; Gonda, S.R.; Gathings, W.E.; McDonald, J.M. Modeled microgravity disrupts collagen I/integrin signaling during osteoblastic differentiation of human mesenchymal stem cells. J. Cell. Biochem. 2004, 93, 697–707. [Google Scholar] [CrossRef]
- Meyers, V.E.; Zayzafoon, M.; Douglas, J.T.; McDonald, J.M. RhoA and cytoskeletal disruption mediate reduced osteoblastogenesis and enhanced adipogenesis of human mesenchymal stem cells in modeled microgravity. J. Bone Miner. Res. 2005, 20, 1858–1866. [Google Scholar] [CrossRef]
- Gershovich, P.M.; Gershovich, J.G.; Zhambalova, A.P.; Romanov, Y.A.; Buravkova, L.B. Cytoskeletal proteins and stem cell markers gene expression in human bone marrow mesenchymal stromal cells after different periods of simulated microgravity. Acta Astronaut. 2012, 70, 36–42. [Google Scholar] [CrossRef]
- Gershovich, P.M.; Gershovich, Y.G.; Buravkova, L.B. Molecular genetic features of human mesenchymal stem cells after their osteogenic differentiation under the conditions of microgravity. Hum. Physiol. 2013, 39, 540–544. [Google Scholar] [CrossRef]
- Sheyn, D.; Pelled, G.; Netanely, D.; Domany, E.; Gazit, D. The effect of simulated microgravity on human mesenchymal stem cells cultured in an osteogenic differentiation system: A bioinformatics study. Tissue Eng. Part A 2003, 16, 3403–3412. [Google Scholar]
- Morey-Holton, E.R.; Globus, R.K. Hindlimb unloading rodent model: Technical aspects. J. Appl. Physiol. 2002, 92, 1367–1377. [Google Scholar] [CrossRef]
- Kostenuik, P.J.; Halloran, B.P.; Morey-Holton, E.R.; Bikle, D.D. Skeletal unloading inhibits the in vitro proliferation and differentiation of rat osteoprogenitor cells. Am. J. Physiol. 1997, 273, E1133–E1139. [Google Scholar]
- Basso, N.; Bellows, C.G.; Heersche, J.N.M. Effect of simulated weightlessness on osteoprogenitor cell number and proliferation in young and adult rats. Bone 2005, 36, 173–183. [Google Scholar] [CrossRef]
- Barzegari, A.; Saei, A.A. An update to space biomedical research: Tissue engineering in microgravity bioreactors. Bioimpacts 2012, 2, 23–32. [Google Scholar]
- Ksiazek, K. A comprehensive review on mesenchymal stem cell growth and senescence. Rejuvenation Res. 2009, 12, 105–116. [Google Scholar] [CrossRef]
- Gunes, C.; Rudolph, K.L. The role of telomeres in stem cells and cancer. Cell 2013, 152, 390–393. [Google Scholar] [CrossRef]
- Yuge, L.; Kajiume, T.; Tahara, H.; Kawahara, Y.; Umeda, C.; Yoshimoto, R.; Wu, S.L.; Yamaoka, K.; Asashima, M.; Kataoka, K.; et al. Microgravity potentiates stem cell proliferation while sustaining the capability of differentiation. Stem Cells Dev. 2006, 15, 921–929. [Google Scholar] [CrossRef]
- Sun, L.; Gan, B.; Fan, Y.; Xie, T.; Hu, Q.; Zhuang, F. Simulated microgravity alters multipotential differentiation of rat mesenchymal stem cells in association with reduced telomerase activity. Acta Astronaut. 2008, 63, 968–973. [Google Scholar] [CrossRef]
- Bradamante, S.; Villa, A.; Versari, S.; Barenghi, L.; Orlandi, I.; Vai, M. Oxidative stress and alterations in actin cytoskeleton trigger glutathione efflux in Saccharomyces cerevisiae. Biochim. Biophys. Acta 2010, 1803, 1376–1385. [Google Scholar] [CrossRef]
- Versari, S.; Longinotti, G.; Barenghi, L.; Maier, J.A.M.; Bradamante, S. The challenging environment on board the International Space Station affects endothelial cell function by triggering oxidative stress through thioredoxin interacting protein overexpression: The ESA-SPHINX experiment. FASEB J. 2013, 27, 4466–4475. [Google Scholar] [CrossRef]
- Tamma, R.; Colaianni, G.; Camerino, C.; di Benedetto, A.; Greco, G.; Strippoli, M.; Vergari, R.; Grano, A.; Mancini, L.; et al. Microgravity during spaceflight directly affects in vitro osteoclastogenesis and bone resorption. FASEB J. 2009, 23, 2549–2554. [Google Scholar] [CrossRef]
- Montufar-Solis, D.; Duke, P.J.; Durnova, G. Spaceflight and age affect tibial epiphyseal growth plate histomorphometry. J. Appl. Physiol. 1992, 73, S19–S25. [Google Scholar]
- Garetto, L.P.; Morey, E.R.; Durnova, G.N.; Kaplansky, A.S.; Roberts, W.E. Preosteoblast production in COSMOS 2044 rats: Short-term recovery of osteogenic potential. J. Appl. Physiol. 1992, 73, S14–S18. [Google Scholar]
- Tavella, S.; Ruggiu, A.; Giuliani, A.; Brun, F.; Canciani, B.; Manescu, A.; Marozzi, K.; Cilli, M.; Costa, D.; Liu, Y.; et al. Bone turnover in wild type and pleiotrophin-transgenic mice housed for three months in the International Space Station (ISS). PLoS One 2012, 7, e33179. [Google Scholar]
- Higuera, G.A.; van Boxtel, A.; van Blitterswijk, C.A.; Moroni, L. The physics of tissue formation with mesenchymal stem cells. Trends Biotechnol. 2012, 30, 583–590. [Google Scholar] [CrossRef]
- Titushkin, I.; Cho, M. Distinct membrane mechanical properties of human mesenchymal stem cells determined using laser optical tweezers. Biophys. J. 2006, 90, 2582–2591. [Google Scholar] [CrossRef]
- Wang, Y.K.; Chen, C.S. Cell adhesion and mechanical stimulation in the regulation of mesenchymal stem cell differentiation. J. Cell. Mol. Med. 2013, 17, 823–832. [Google Scholar] [CrossRef]
- Guilak, F.; Cohen, D.M.; Estes, B.T.; Gimble, J.M.; Liedtke, W.; Chen, C.S. Control of stem cell fate by physical interactions with the extracellular matrix. Cell Stem Cell 2009, 5, 17–26. [Google Scholar] [CrossRef]
- Riddle, R.C.; Taylor, A.F.; Genetos, D.C.; Donahue, H.J. MAP kinase and calcium signaling mediate fluid flow-induced human mesenchymal stem cell proliferation. Am. J. Physiol. Cell Physiol. 2006, 290, C776–C784. [Google Scholar]
- Riddle, R.C.; Donahue, H.J. From streaming-potentials to shear stress: 25 years of bone cell mechanotransduction. J. Orthop. Res. 2009, 27, 143–149. [Google Scholar] [CrossRef]
- MacQueen, L.; Sun, Y.; Simmons, C.A. Mesenchymal stem cell mechanobiology and emerging experimental platforms. J. R. Soc. Interface 2013, 10. [Google Scholar] [CrossRef]
- Halder, G.; Dupont, S.; Piccolo, S. Transduction of mechanical and cytoskeletal cues by YAP and TAZ. Nat. Rev. Mol. Cell Biol. 2012, 13, 591–600. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bradamante, S.; Barenghi, L.; Maier, J.A.M. Stem Cells toward the Future: The Space Challenge. Life 2014, 4, 267-280. https://doi.org/10.3390/life4020267
Bradamante S, Barenghi L, Maier JAM. Stem Cells toward the Future: The Space Challenge. Life. 2014; 4(2):267-280. https://doi.org/10.3390/life4020267
Chicago/Turabian StyleBradamante, Silvia, Livia Barenghi, and Jeanette A.M. Maier. 2014. "Stem Cells toward the Future: The Space Challenge" Life 4, no. 2: 267-280. https://doi.org/10.3390/life4020267
APA StyleBradamante, S., Barenghi, L., & Maier, J. A. M. (2014). Stem Cells toward the Future: The Space Challenge. Life, 4(2), 267-280. https://doi.org/10.3390/life4020267