
Association of TMEM173/STING1 Gene Variants with Severe COVID-19 Among Fully Vaccinated vs. Non-Vaccinated Individuals

 ,
,  and
and
Abstract
1. Introduction
2. Methods
3. Statistical Analysis
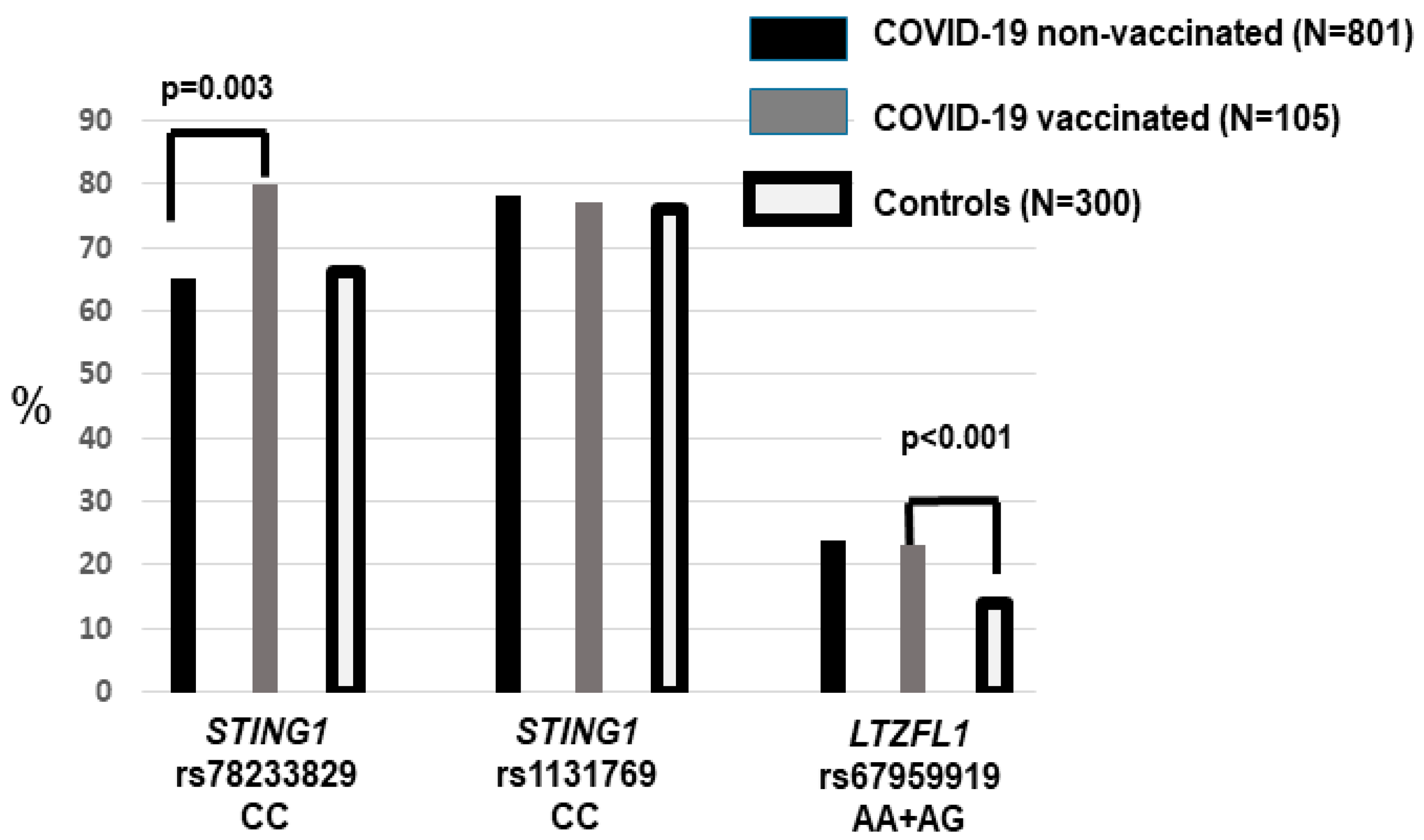
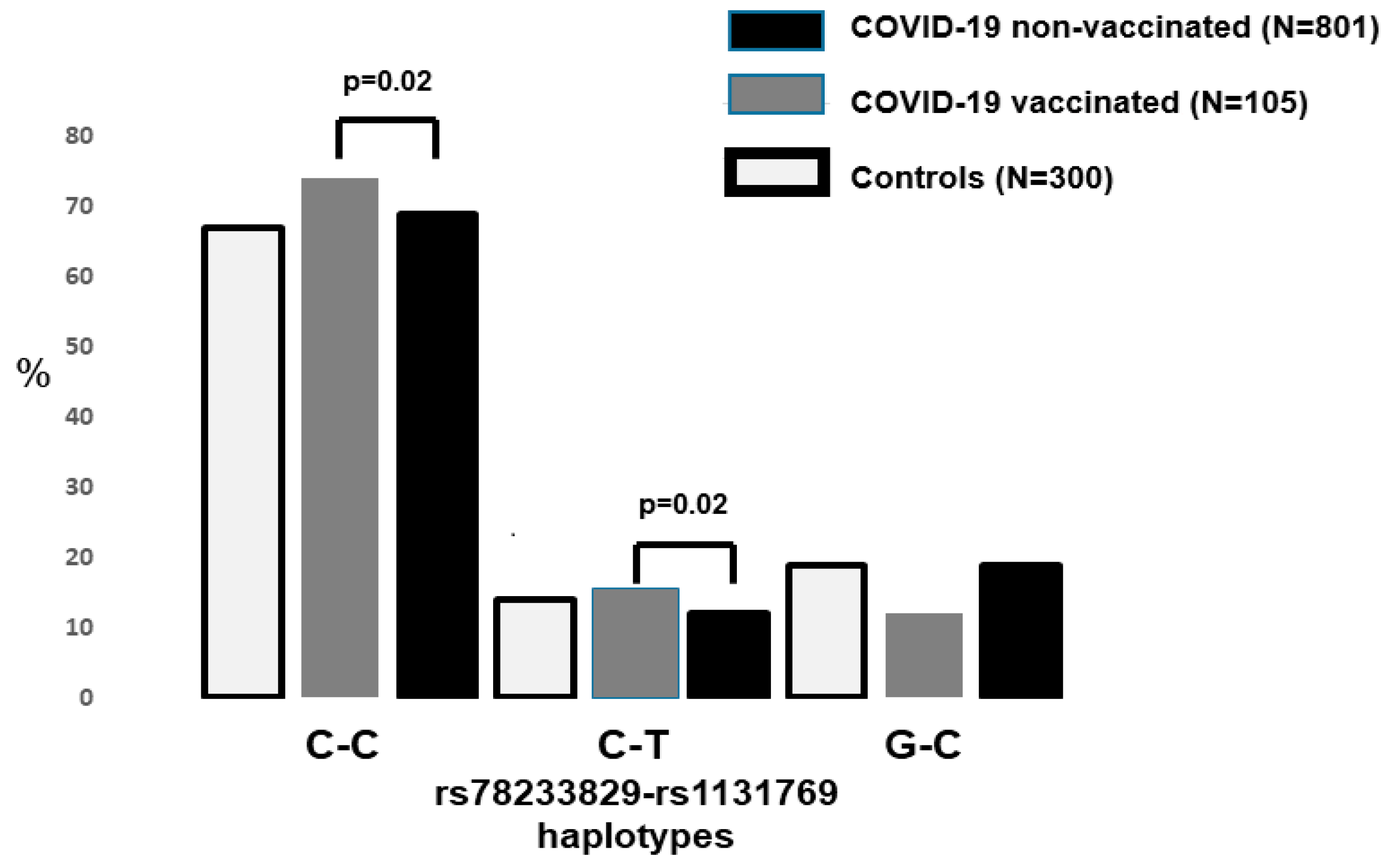
4. Results
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Webb, L.G.; Fernandez-Sesma, A. RNA viruses and the cGAS-STING pathway: Reframing our understanding of innate immune sensing. Curr. Opin. Virol. 2022, 53, 101206. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Damania, B. TecGAS-STING defense pathway and its counteraction by viruses. Cell Host Microbe 2016, 19, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.K.; Gack, M.U. Viral evasion of intracellular DNA and RNA sensing. Nat. Rev. Microbiol. 2016, 14, 360–373. [Google Scholar] [CrossRef] [PubMed]
- Ablasser, A.; Chen, Z.J. cGAS in action: Expanding roles in immunity and inflammation. Science 2019, 363, eaat8657. [Google Scholar] [CrossRef] [PubMed]
- Bouis, D.; Kirstetter, P.; Arbogast, F.; Lamon, D.; Delgado, V.; Jung, S.; Ebel, C.; Jacobs, H.; Knapp, A.-M.; Jeremiah, N.; et al. Severe combined immunodeficiency in stimulator of interferon genes (STING) V154M/wild-type mice. J. Allergy Clin. Immunol. 2019, 143, 712–725.e5. [Google Scholar] [CrossRef] [PubMed]
- Hopfner, K.-P.; Hornung, V. Molecular mechanisms and cellular functions of cGAS–STING signalling. Nat. Rev. Mol. Cell Biol. 2020, 21, 501–521. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Liao, X.; Peng, J.; Quan, J.; Zhang, H.; Huang, Z.; Yi, B. PCSK9 causes inflammation and cGAS/STINGpathway activation indiabeticnephropathy. FASEB J. 2023, 37, e23127. [Google Scholar] [CrossRef] [PubMed]
- Frémond, M.-L.; Crow, Y.J. STING-Mediated Lung Inflammation and Beyond. J. Clin. Immunol. 2021, 41, 501–514. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Ghosh, S.; Vaidya, T.; Bammidi, S.; Huang, C.; Shang, P.; Nair, A.P.; Chowdhury, O.; Stepicheva, N.A.; Strizhakova, A.; et al. Activated cGAS/STINGsignaling elicits endothelial cell senescence in earlydia-beticretinopathy. JCI Insight 2023, 8, e168945. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yang, X.; Zheng, Y.; Yang, Y.; Xing, Y.; Chen, Z. SARS coronavirus papain-like protease inhibits the type I interferon signaling pathway through interaction with the STING-TRAF3-TBK1 complex. Protein Cell 2014, 5, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Zhuang, M.-W.; Deng, J.; Zheng, Y.; Zhang, J.; Nan, M.-L.; Zhang, X.-J.; Gao, C.; Wang, P.-H. SARS-CoV-2 ORF9b antagonizes type I and III interferons by targeting multiple components of the RIG-I/MDA-5–MAVS, TLR3–TRIF, and cGAS–STING signaling pathways. J. Med. Virol. 2021, 93, 5376–5389. [Google Scholar] [CrossRef] [PubMed]
- Rui, Y.; Su, J.; Shen, S.; Hu, Y.; Huang, D.; Zheng, W.; Lou, M.; Shi, Y.; Wang, M.; Chen, S.; et al. Unique and complementary suppression of cGAS-STING and RNA sensing-triggered innate immune responses by SARS-CoV-2 proteins. Signal Transduct. Target. Ther. 2021, 6, 123. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Zhang, X.; Lei, X.; Xiao, X.; Jiao, T.; Ma, R.; Dong, X.; Jiang, Q.; Wang, W.; Shi, Y.; et al. Sensing of cytoplasmic chromatin by cGAS activates innate immune response in SARS-CoV-2 infection. Signal Transduct. Target. Ther. 2021, 6, 382. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wei, L.; Xu, F.; Zhao, F.; Huang, Y.; Fan, Z.; Mei, S.; Hu, Y.; Zhai, L.; Guo, J.; et al. SARS-CoV-2 spike protein–induced cell fusion activates the cGAS-STING pathway and the interferon response. Sci. Signal. 2022, 15, eabg8744. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, H.-S.; Chung, J.H. Molecular mechanisms of mitochondrial DNA release and activation of the cGAS-STING pathway. Exp. Mol. Med. 2023, 55, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.H.; Wu, D.W.; Wu, C.H.; Hung, L.F.; Huang, C.Y.; Ka, S.M.; Chen, A.; Ho, L.J. USP18 enhances denguevirusreplication by regula-tingmitochondrialDNArelease. Sci. Rep. 2023, 13, 20126. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, S.; Subbian, S. Critical determinants of cytokine storm and type I interferon response in COVID-19 pathogene-sis. Clin. Microbiol. Rev. 2021, 34, e0016321. [Google Scholar] [CrossRef]
- Neufeldt, C.J.; Cerikan, B.; Cortese, M.; Frankish, J.; Lee, J.-Y.; Plociennikowska, A.; Heigwer, F.; Prasad, V.; Joecks, S.; Burkart, S.S.; et al. SARS-CoV-2 infection induces a pro-inflammatory cytokine response through cGAS-STING and NF-κB. Commun. Biol. 2022, 5, 45. [Google Scholar] [CrossRef] [PubMed]
- Meibers, H.E.; Warrick, K.A.; VonHandorf, A.; Vallez, C.N.; Kawarizadeh, K.; Saha, I.; Donmez, O.; Jain, V.G.; Kottyan, L.C.; Weirauch, M.T.; et al. Effector memory T cells induce innate inflammation by triggering DNA damage and a non-canonical STING pathway in dendritic cells. Cell Rep. 2023, 42, 113180. [Google Scholar] [CrossRef] [PubMed]
- Humphries, F.; Shmuel-Galia, L.; Jiang, Z.; Wilson, R.; Landis, P.; Ng, S.-L.; Parsi, K.M.; Maehr, R.; Cruz, J.; Morales, A.; et al. A diamidobenzimidazole STING agonist protects against SARS-CoV-2 infection. Sci. Immunol. 2021, 6, eabi9002. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Ferretti, M.; Ying, B.; Descamps, H.; Lee, E.; Dittmar, M.; Lee, J.S.; Whig, K.; Kamalia, B.; Dohnalová, L.; et al. Pharmacological activation of STING blocks SARS-CoV-2 infection. Sci. Immunol. 2021, 6, eabi9007. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Zhang, Y.; Wang, L.; Yao, X.; Wu, D.; Cheng, J.; Pan, X.; Liu, H.; Yan, Z.; Gao, L. Inhibition of coronavirus infection by a synthetic STING agonist in primary human airway system. Antivir. Res. 2021, 187, 105015. [Google Scholar] [CrossRef] [PubMed]
- Padron-Regalado, E.; Ulaszewska, M.; Douglas, A.D.; Hill, A.V.S.; Spencer, A.J. STING-pathway modulation to enhance the im-munogenicity of adenoviral-vectored vaccines. Sci. Rep. 2022, 12, 14464. [Google Scholar] [CrossRef] [PubMed]
- Van Herck, S.; Feng, B.; Tang, L. Delivery of STING agonists for adjuvanting subunit vaccines. Adv. Drug Deliv. Rev. 2021, 179, 114020. [Google Scholar] [CrossRef] [PubMed]
- Gutjahr, A.; Papagno, L.; Nicoli, F.; Kanuma, T.; Kuse, N.; Cabral-Piccin, M.P.; Rochereau, N.; Gostick, E.; Lioux, T.; Perouzel, E.; et al. The STING ligand cGAMP potentiates the efficacy of vaccine-induced CD8+ T cells. J. Clin. Investig. 2019, 4, e125107. [Google Scholar] [CrossRef] [PubMed]
- Chauveau, L.; Bridgeman, A.; Tan, T.K.; Beveridge, R.; Frost, J.N.; Rijal, P.; Pedroza-Pacheco, I.; Partridge, T.; Gilbert-Jaramillo, J.; Knight, M.L.; et al. Inclusion of cGAMP within virus-like particle vaccines enhances their immunogenicity. EMBO Rep. 2021, 22, e52447. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhou, J.; Xu, W.; Deng, W.; Wang, Y.; Wang, M.; Wang, Q.; Hsieh, M.; Dong, J.; Wang, X.; et al. A novel STING agonist-adjuvanted pan-sarbecovirus vaccine elicits potent and durable neutralizing antibody and T cell responses in mice, rabbits and NHPs. Cell Res. 2022, 32, 269–287. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yan, J.; Hou, X.; Wang, C.; Kang, D.D.; Xue, Y.; Du, S.; Deng, B.; McComb, D.W.; Liu, S.-L.; et al. STING Agonist-Derived LNP-mRNA Vaccine Enhances Protective Immunity Against SARS-CoV-2. Nano Lett. 2023, 23, 2593–2600. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Xu, L.-G.; Yang, I.V.; Davidson, E.J.; Schwartz, D.A.; Wurfel, M.M.; Cambier, J.C. Identification and characterization of a loss-of-function human MPYS variant. Genes Immun. 2011, 12, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Yi, G.; Brendel, V.P.; Shu, C.; Li, P.; Palanathan, S.; Kao, C.C.; Li, K. Single nucleotide polymorphisms of human STING can affect innate immune response to cyclic dinucleotides. PLoS ONE 2013, 8, e77846. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Moreno, J.S.; Hamann, L.; Shah, J.A.; Verbon, A.; Mockenhaupt, F.P.; Puzianowska-Kuznicka, M.; Naujoks, J.; Sander, L.E.; Witzenrath, M.; Cambier, J.C.; et al. The common HAQ STING variant impairs cGAS-dependent antibacterial responses and is associated with susceptibility to Legionnaires’ disease in humans. PLoS Pathog. 2018, 14, e1006829. [Google Scholar] [CrossRef] [PubMed]
- Nissen, S.K.; Pedersen, J.G.; Helleberg, M.; Kjaer, K.; Thavachelvam, K.; Obel, N.; Tolstrup, M.; Jakobsen, M.R.; Mogensen, T.H. Multiple Homozygous Variants in the STING-Encoding TMEM173 Gene in HIV Long-Term Nonprogressors. J. Immunol. 2018, 200, 3372–3382. [Google Scholar] [CrossRef] [PubMed]
- Hamann, L.; Ruiz-Moreno, J.S.; Szwed, M.; Mossakowska, M.; Lundvall, L.; Schumann, R.R.; Opitz, B.; Puzianowska-Kuznicka, M. STING SNP R293Q Is Associated with a Decreased Risk of Aging-Related Diseases. Gerontology 2018, 65, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Jin, L. TMEM173 variants and potential importance to human biology and disease. Genes Immun. 2018, 20, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, R.B.; Haralambieva, I.H.; Ovsyannikova, I.G.; Voigt, E.A.; Larrabee, B.R.; Schaid, D.J.; Zimmermann, M.T.; Oberg, A.L.; Poland, G.A. Polymorphisms in STING Affect Human Innate Immune Responses to Poxviruses. Front. Immunol. 2020, 11, 567348. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xiao, Y.; Loupakis, F.; Stintzing, S.; Yang, Y.; Arai, H.; Battaglin, F.; Kawanishi, N.; Jayachandran, P.; Soni, S.; et al. Genetic variants involved in the cGAS-STING pathway predict outcome in patients with metastatic colorectal cancer: Data from FIRE-3 and TRIBE trials. Eur. J. Cancer 2022, 172, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, X.; Fan, Y.; Guo, Z.; Duan, X.; Zhou, X.; Yang, Y.; Wang, W. The polymorphisms in cGAS-STING pathway are associated with mitochondrial DNA copy number in coke oven workers. Int. J. Environ. Health Res. 2022, 33, 1070–1080. [Google Scholar] [CrossRef] [PubMed]
- Tang, E.D.; Wang, C.-Y.; Lenz, L.L. Single amino acid change in STING leads to constitutive active signaling. PLoS ONE 2015, 10, e0120090. [Google Scholar] [CrossRef] [PubMed]
- Froechlich, G.; Finizio, A.; Napolano, A.; Amiranda, S.; De Chiara, A.; Pagano, P.; Mallardo, M.; Leoni, G.; Zambrano, N.; Sasso, E. The common H232 STING allele shows impaired activities in DNA sensing, susceptibility to viral infection, and in monocyte cell function, while the HAQ variant possesses wild-type properties. Sci. Rep. 2023, 13, 19541. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Alba, J.M.; Rojo-Alba, S.; Perez-Martinez, Z.; Boga, J.A.; Alvarez-Arguelles, M.E.; Gomez, J.; Herrero, P.; Costales, I.; Alba, L.M.; Martin-Rodriguez, G.; et al. Monitoring and tracking the spread of SARS-CoV-2 in Asturias, Spain. Access Microbiol. 2023, 5, 000573.v4. [Google Scholar] [CrossRef] [PubMed]
- Mazagatos, C.; Delgado-Sanz, C.; Monge, S.; Pozo, F.; Oliva, J.; Sandonis, V.; Gandarillas, A.; Quiñones-Rubio, C.; Ruiz-Sopeña, C.; Gallardo-García, V.; et al. COVID-19 vaccine effectiveness against hospitalization due to SARS-CoV-2: A test-negative design study based on Severe Acute Respiratory Infection (SARI) sentinel surveillance in Spain. Influ. Other Respir. Viruses 2022, 16, 1014–1025. [Google Scholar] [CrossRef] [PubMed]
- Downes, D.J.; Cross, A.R.; Hua, P.; Roberts, N.; Schwessinger, R.; Cutler, A.J.; Munis, A.M.; Brown, J.; Mielczarek, O.; de Andrea, C.E.; et al. Identification of LZTFL1 as a candidate effector gene at a COVID-19 risk locus. Nat. Genet. 2021, 53, 1606–1615. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Pu, F. Updated roles of cGAS-STING signaling in autoimmune diseases. Front. Immunol. 2023, 14, 1254915. [Google Scholar] [CrossRef] [PubMed]
- di Flora, D.C.; Lara, J.P.Z.; Dionizio, A.; Buzalaf, M.A.R. The Dual Role of cGAS-STING Signaling in COVID-19: Implications for Therapy. Cells 2025, 14, 362. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Blaauboer, S.M.; Tucker, H.R.; Mansouri, S.; Ruiz-Moreno, J.S.; Hamann, L.; Schumann, R.R.; Opitz, B.; Jin, L. The Common R71H-G230A-R293Q Human TMEM173 Is a Null Allele. J. Immunol. 2017, 198, 776–787. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| 1st–2nd WaveNon-Vaccinated | 3rd–4th WaveNon-Vaccinated | 3rd–4th WaveVaccinated | Controls | ||||
|---|---|---|---|---|---|---|---|
| Non-ICU | ICU | Non-ICU | ICU | Non-ICU | ICU | ||
| n = 225 | n = 216 | n = 185 | n = 175 | n = 96 | n = 9 | n = 300 | |
| Male | 133 (59%) | 140 (65%) | 113 (61%) | 128 (73%) | 60 (63%) | 7 (78%) | 165 (55%) |
| Female | 92 (41%) | 76 (35%) | 72 (39%) | 47 (27%) | 36 (37%) | 2 (22%) | 135 (45%) |
| <65 years | 143 (64%) | 94 (44%) | 145 (78%) | 102 (58%) | 20 (21%) | 2 (22%) | 160 (53%) |
| ≥65 years | 82 (36%) | 122 (56%) | 40 (22%) | 73 (42%) | 80 (79%) | 7 (78%) | 140 (47%) |
| Death | 5 (2%) | 59 (27%) | 2 (2%) | 32 (18%) | 0 | 1 (11%) | - |
| Hypertensives | 85 (38%) | 127 (59%) | 59 (32%) | 95 (54%) | 60 (63%) | 6 (67%) | - |
| Dyslipidemia | 68 (30%) | 98 (45%) | 56 (30%) | 83 (48%) | 49 (51%) | 5 (56%) | - |
| STING1 rs78233829 C/G | |||||||
| GG | 4 (2%) | 10 (4%) | 9 (5%) | 9 (5%) | 4 (4%) | 0 | 11 (3%) |
| CG | 72 (32%) | 64 (30%) | 56 (30%) | 54 (31%) | 15 (16%) | 2 (22%) | 92 (31%) |
| CC | 149 (66%) | 142 (66%) | 120 (65%) | 112 (64%) | 77 (80%) | 7 (78%) | 197 (66%) |
| MAF G | 0.18 | 0.19 | 0.20 | 0.21 | 0.12 | 0.11 | 0.19 |
| STING1 rs1131769 C/T | |||||||
| TT | 4 (2%) | 4 (2%) | 2 (1%) | 3 (2%) | 4 (5%) | 0 | 11 (4%) |
| CT | 57 (25%) | 38 (18%) | 40 (22%) | 30 (17%) | 18 (19%) | 1 (11%) | 62 (21%) |
| CC | 164 (73%) | 174 (81%) | 143 (77%) | 142 (81%) | 73 (76%) | 8 (89%) | 227 (76%) |
| MAF T | 0.14 | 0.11 | 0.12 | 0.10 | 0.14 | 0.06 | 0.14 |
| LZTFL1 rs67959919 A/G | |||||||
| AA | 3 (1%) | 2 (1%) | 4 (2%) | 4 (3%) | 1 (1%) | 0 | 4 (1%) |
| AG | 44 (20%) | 54 (25%) | 37 (20%) | 40 (23%) | 21 (22%) | 2 (22%) | 39 (13%) |
| GG | 178 (77%) | 160 (74%) | 144 (78%) | 131 (75%) | 74 (77%) | 7 (78%) | 157 (86%) |
| MAF A | 0.11 | 0.13 | 0.12 | 0.14 | 0.12 | 0.11 | 0.12 |
| <65 YEARS | ≥65 YEARS | |||||
|---|---|---|---|---|---|---|
| ICU n = 196 | NON-ICU n = 288 | p-Value * | ICU n = 195 | NON-ICU n = 122 | p-Value * | |
| rs78233829 | ||||||
| GG | 130 (66%) | 189 (65%) | ns | 124 (64%) | 80 (66%) | |
| CG | 55 (28%) | 89 (31%) | 63 (32%) | 39 (32%) | ns | |
| CC | 11 (6%) | 10 (4%) | 8 (4%) | 3 (2%) | ||
| MAF G | 0.20 | 0.19 | 0.20 | 0.19 | ||
| rs1131769 | ||||||
| CC | 153 (78%) | 217 (75%) | 163 (84%) | 90 (74%) | ||
| CT | 40 (20%) | 65 (23%) | ns | 28 (14%) | 32 (26%) | ns |
| TT | 3 (2%) | 6 (2%) | 4 (2%) | 0 | ||
| MAF T | 0.12 | 0.13 | 0.10 | 0.13 | ||
| rs67959919 | ||||||
| AA | 6 (3%) | 5 (2%) | ns | 0 | 2 (2%) | ns |
| AG | 49 (25%) | 58 (20%) | 45 (23%) | 23 (19%) | ||
| GG | 141 (72%) | 225 (78%) | 150 (77%) | 97 (80%) | ||
| MAF A | 0.16 | 0.12 | 0.12 | 0.11 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vázquez-Coto, D.; García-Clemente, M.; Albaiceta, G.M.; Amado, L.; Vega-Prado, L.M.; García-Lago, C.; Lorca, R.; Gómez, J.; Coto, E. Association of TMEM173/STING1 Gene Variants with Severe COVID-19 Among Fully Vaccinated vs. Non-Vaccinated Individuals. Life 2025, 15, 1171. https://doi.org/10.3390/life15081171
Vázquez-Coto D, García-Clemente M, Albaiceta GM, Amado L, Vega-Prado LM, García-Lago C, Lorca R, Gómez J, Coto E. Association of TMEM173/STING1 Gene Variants with Severe COVID-19 Among Fully Vaccinated vs. Non-Vaccinated Individuals. Life. 2025; 15(8):1171. https://doi.org/10.3390/life15081171
Chicago/Turabian StyleVázquez-Coto, Daniel, Marta García-Clemente, Guillermo M. Albaiceta, Laura Amado, Lorena M. Vega-Prado, Claudia García-Lago, Rebeca Lorca, Juan Gómez, and Eliecer Coto. 2025. "Association of TMEM173/STING1 Gene Variants with Severe COVID-19 Among Fully Vaccinated vs. Non-Vaccinated Individuals" Life 15, no. 8: 1171. https://doi.org/10.3390/life15081171
APA StyleVázquez-Coto, D., García-Clemente, M., Albaiceta, G. M., Amado, L., Vega-Prado, L. M., García-Lago, C., Lorca, R., Gómez, J., & Coto, E. (2025). Association of TMEM173/STING1 Gene Variants with Severe COVID-19 Among Fully Vaccinated vs. Non-Vaccinated Individuals. Life, 15(8), 1171. https://doi.org/10.3390/life15081171