
Samotherium boissieri from the Late Miocene of Southern Italy


Abstract
1. Introduction
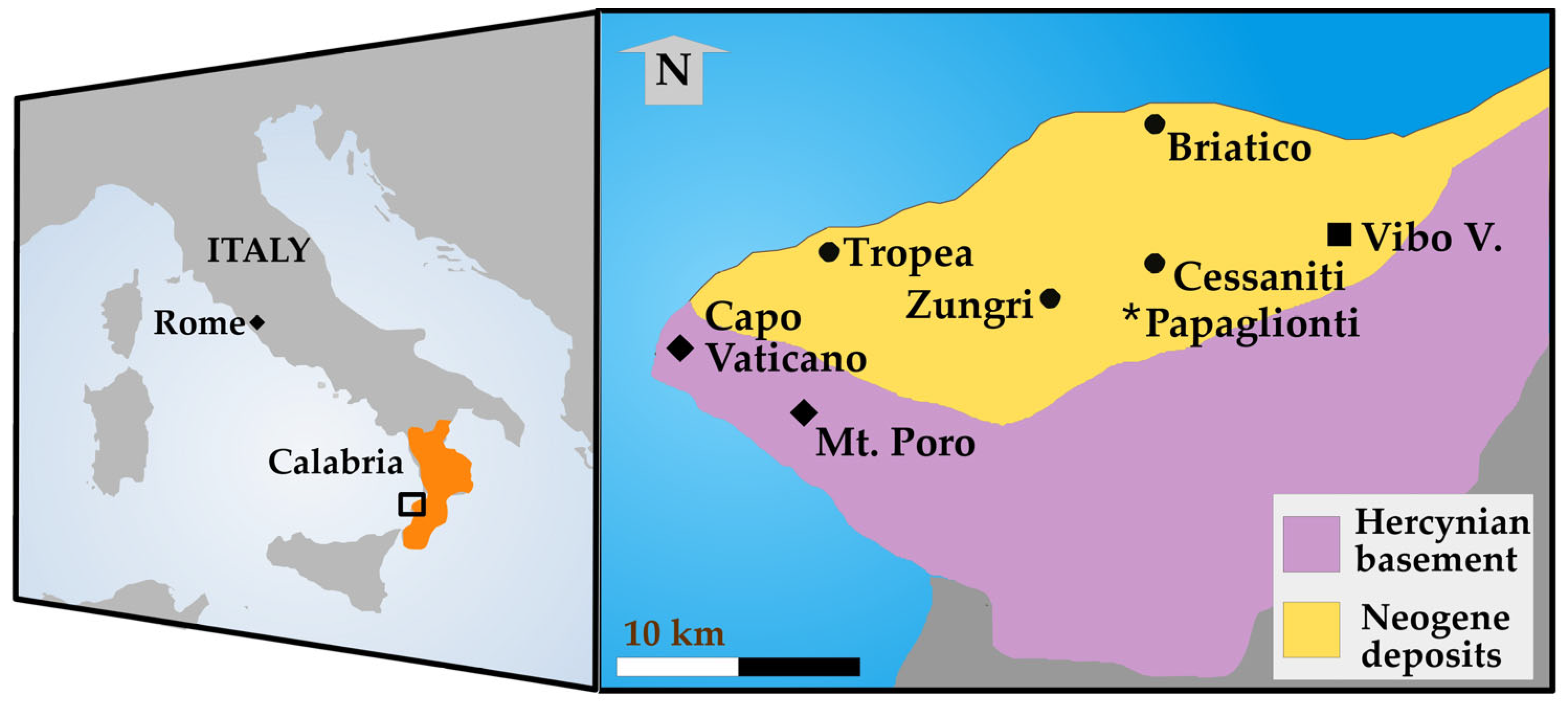
Localities and Geological Setting
2. Materials and Methods
3. Systematic Paleontology
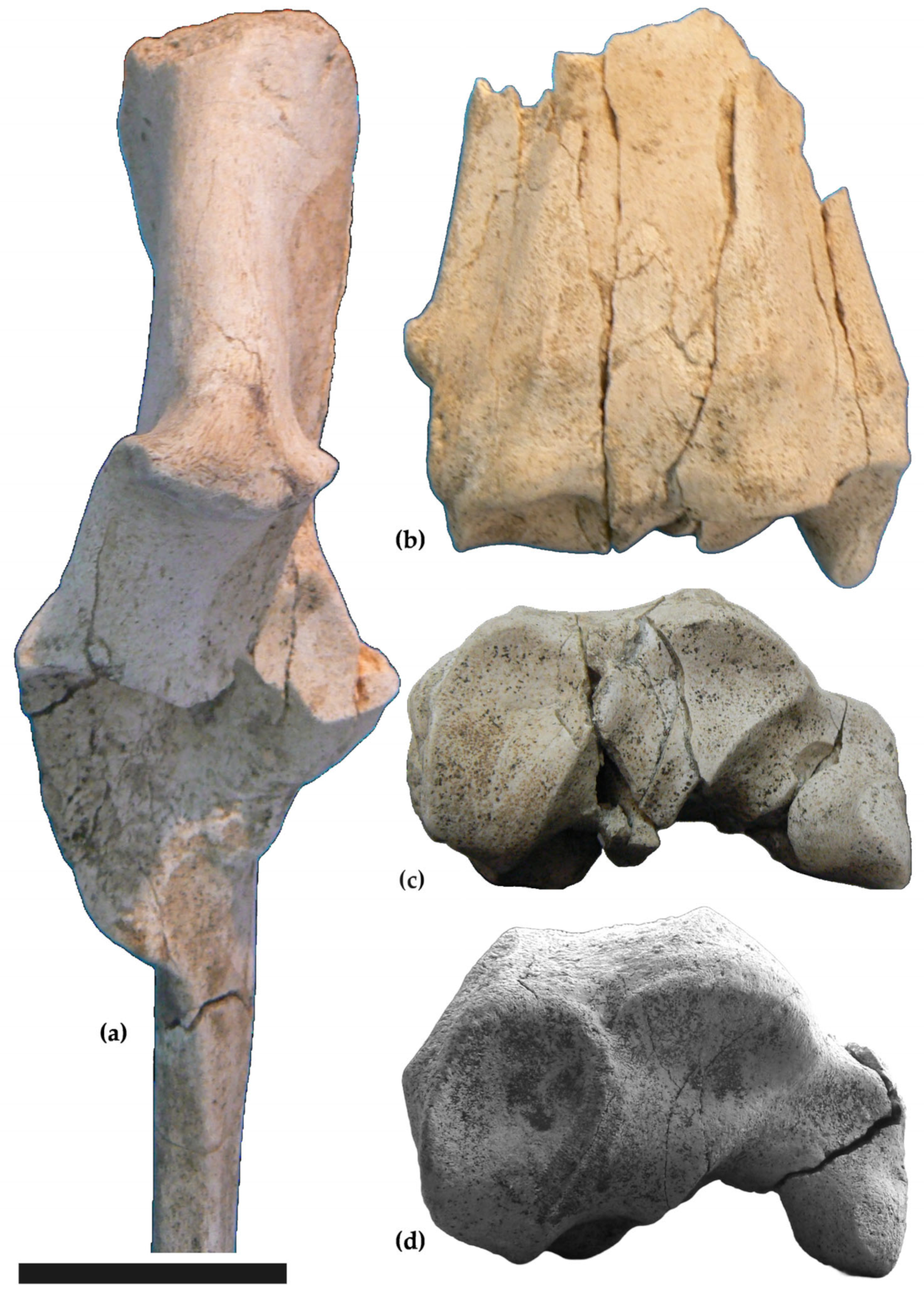
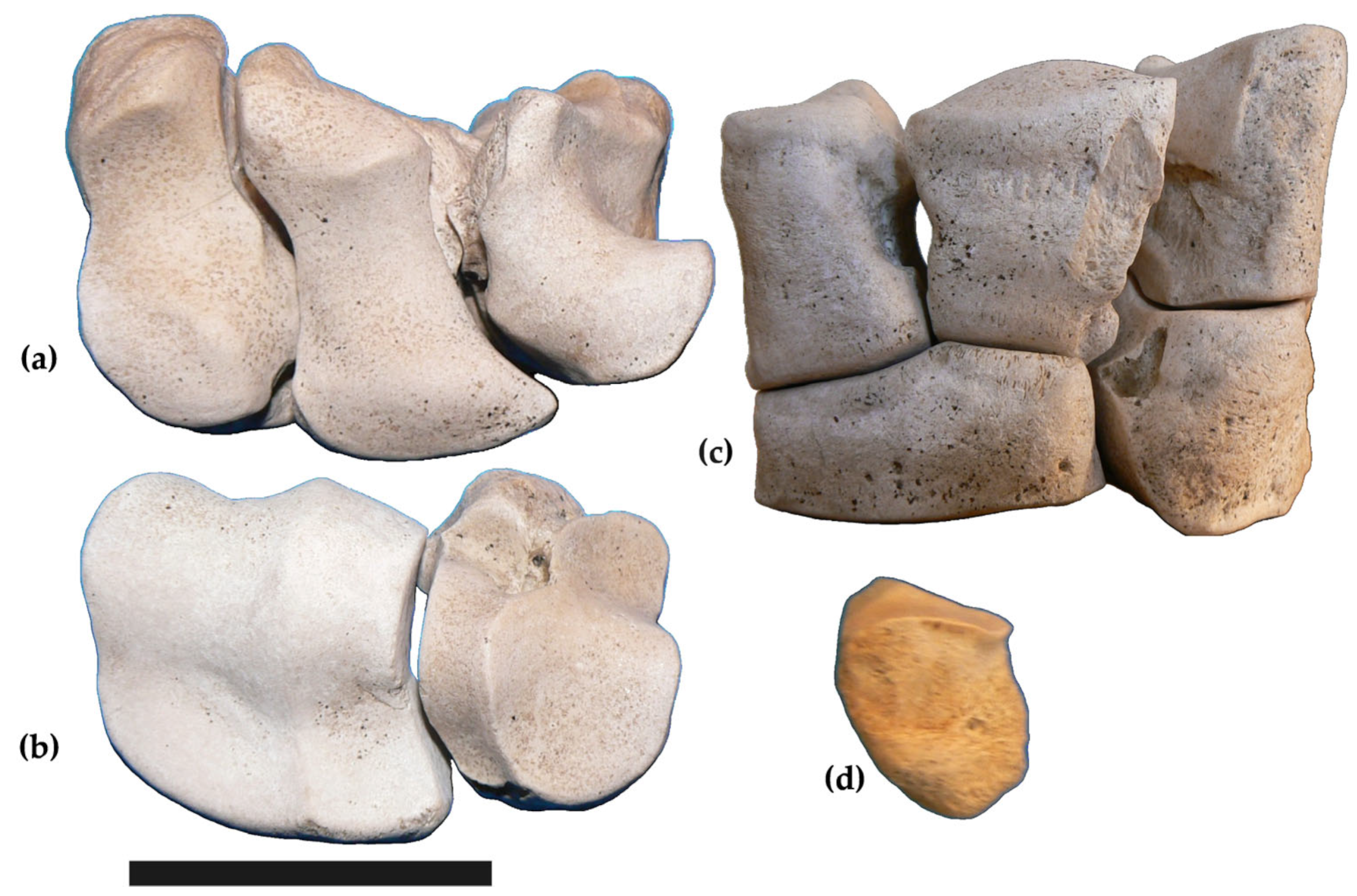
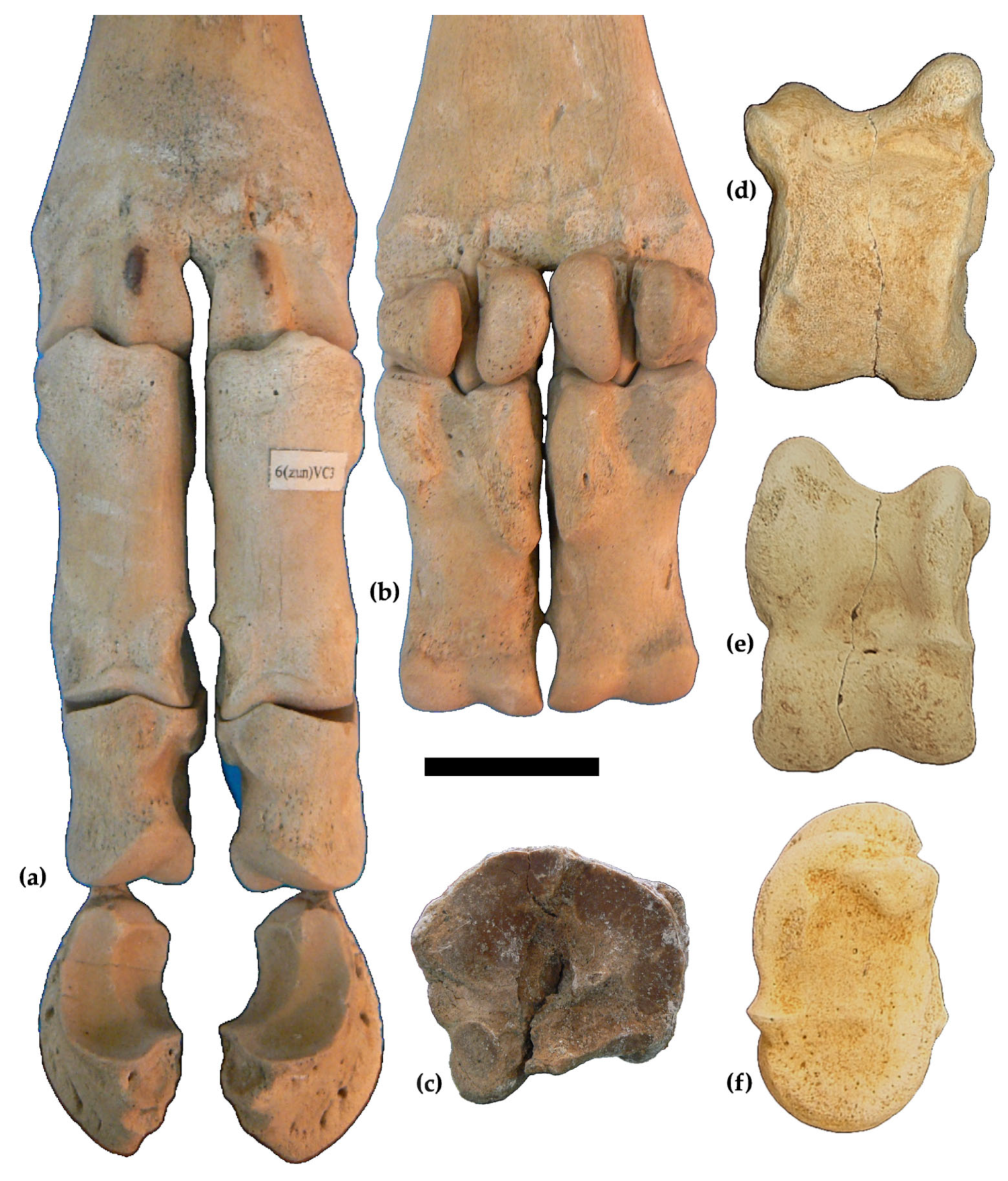
3.1. Descriptions
Forelimbs
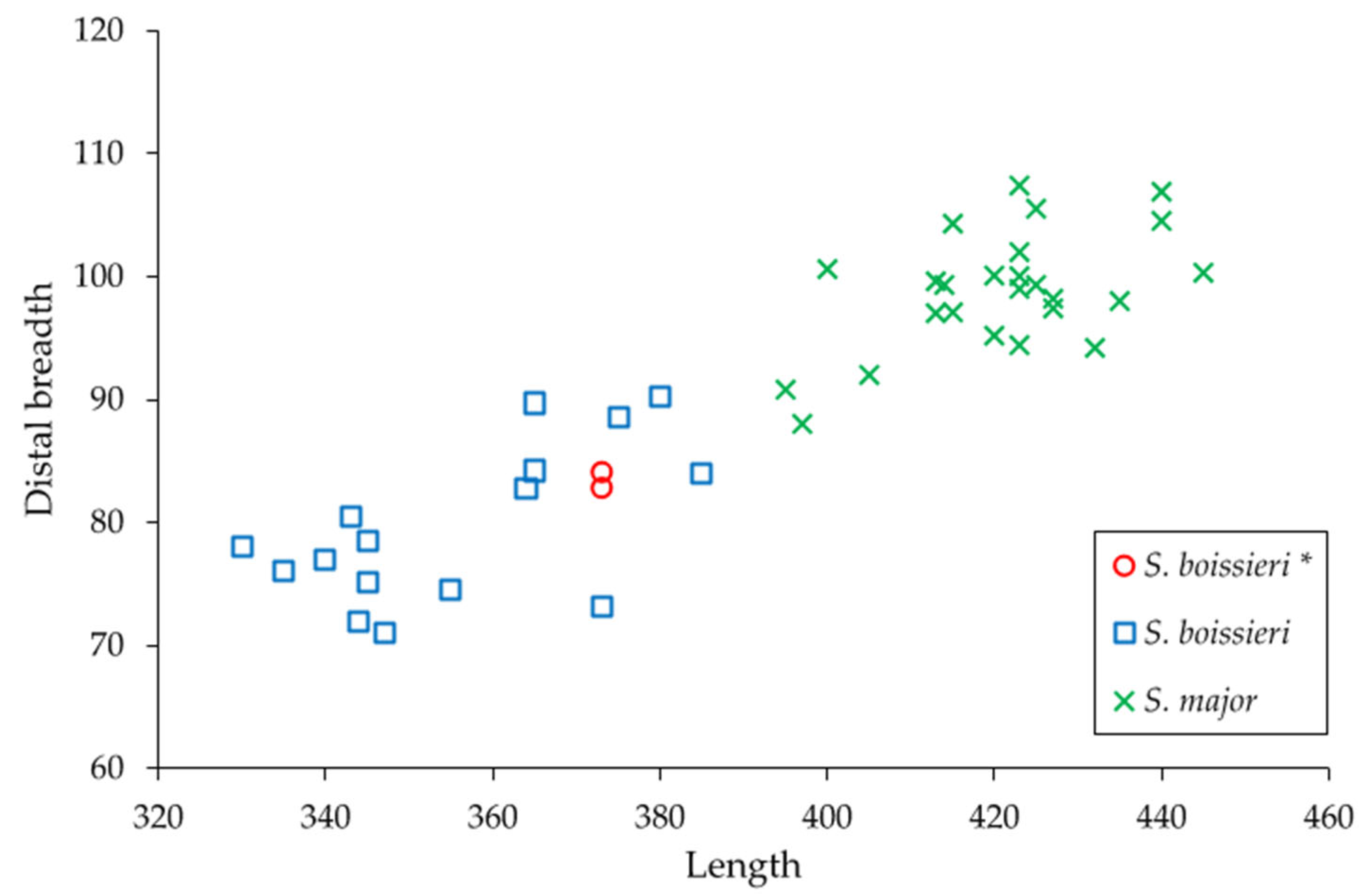
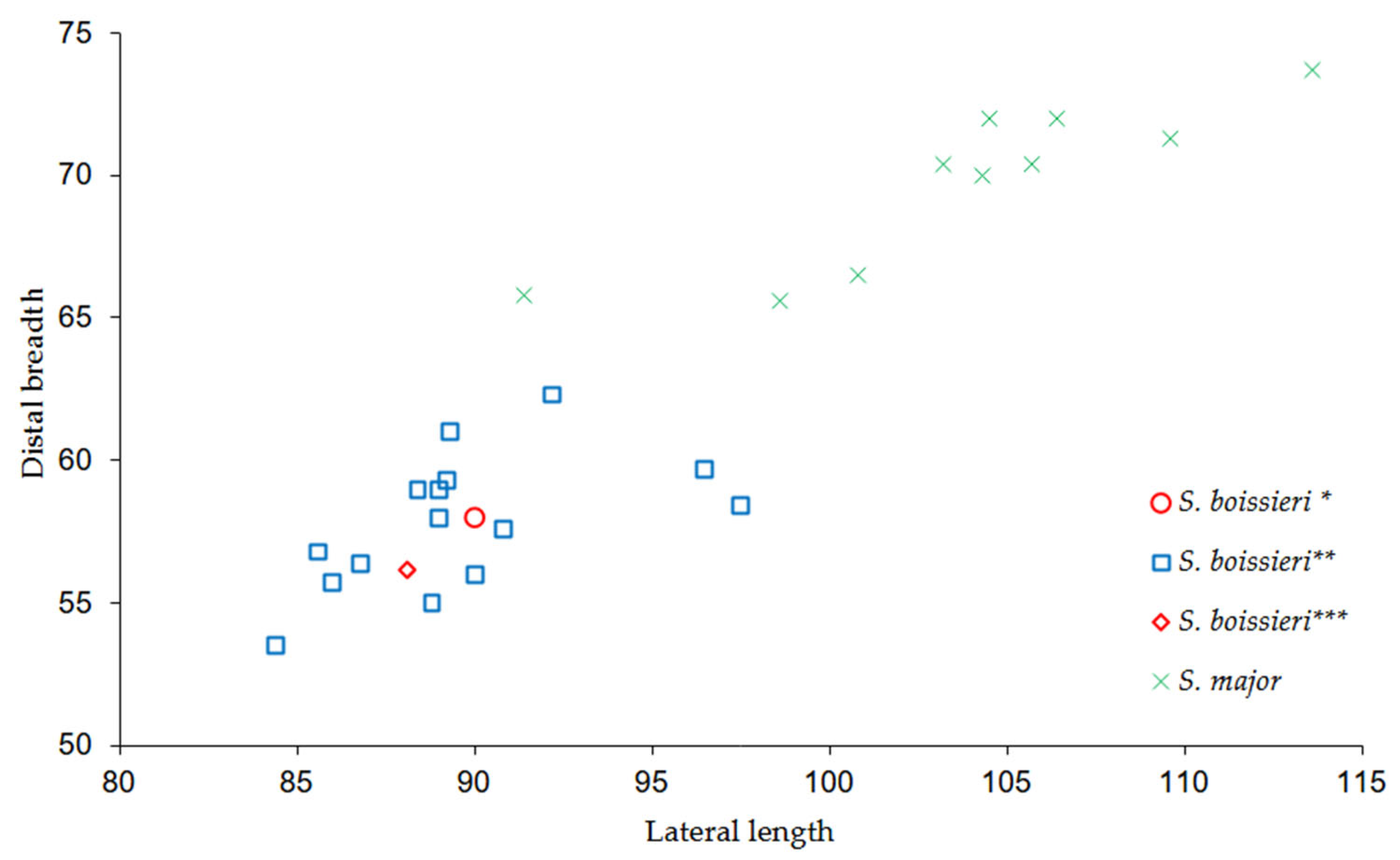
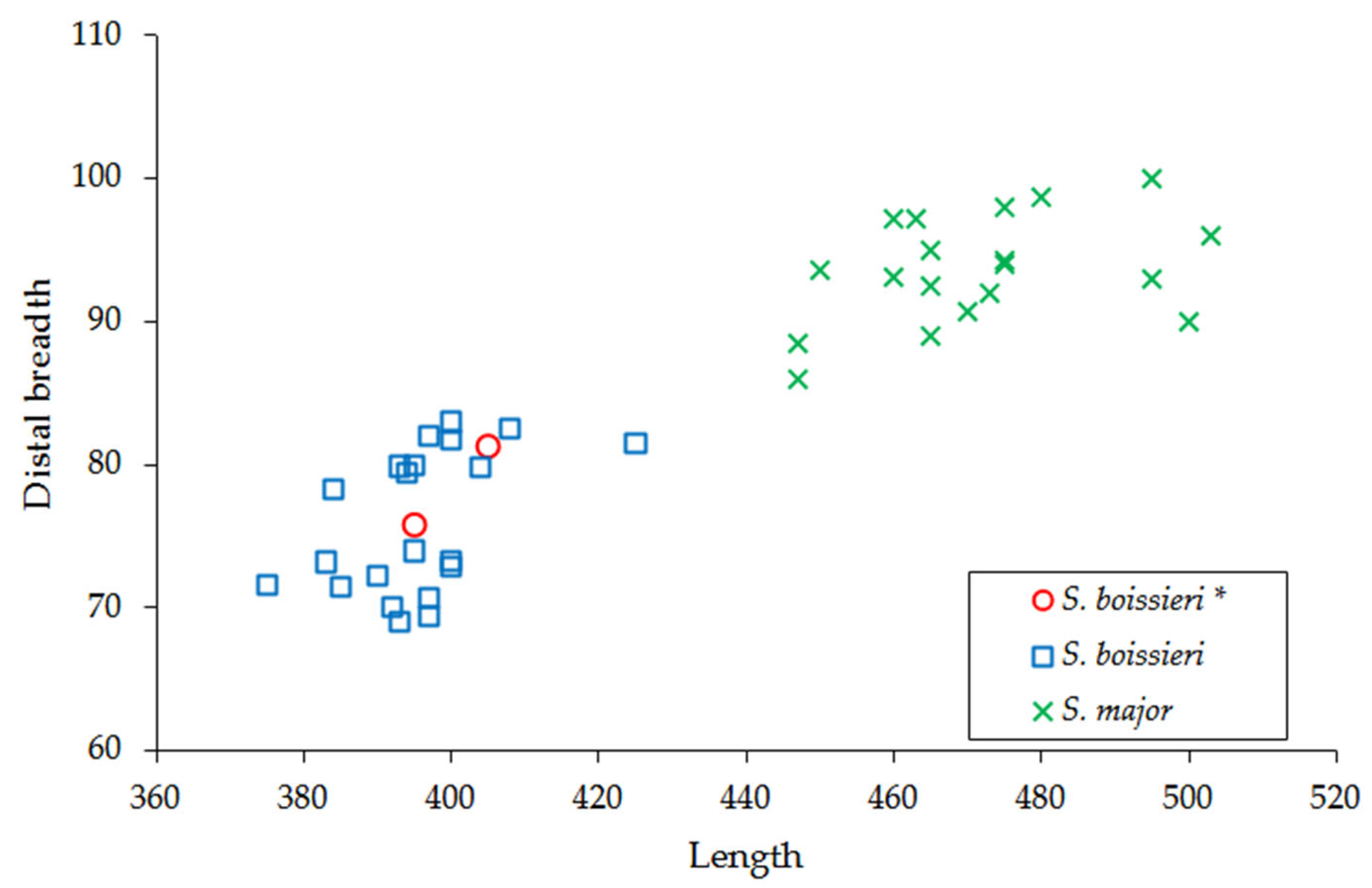
3.2. Comparisons
4. Discussion
4.1. Biochonology and Distribution
4.2. Paleoecology
4.3. Paleobiogeography
- The Gravitelli mammal assemblage (Messina, Sicily) is based on fossils no longer available to the study, because they were destroyed during the earthquake that hit the city of Messina in 1908. The revision of available museum casts and the papers by Seguenza [69,70] revealed a non-endemic fauna with European affinity and possibly two African taxa [71,72]. The re-consideration of the stratigraphy described by Seguenza could indicate an age more ancient than the Messinian age originally attributed, maybe older than 7 Ma [71]. The site is no longer investigable for the intense urbanization. The mammal association does not have taxa in common with Cessaniti, except for Ceratotherium sp.
- The paleogeographic scenarios for the Cessaniti land are as follows:
- Land connected to North Africa: in this case, the Greco-Iranian taxa come from an expansion to Africa of the Pikermian biome; this widely hypothesized condition is not strongly supported by evidence.
- Land connected to Eastern Europe: in this case, the land was subject to complex movements, maybe related to the Adria plate; this hypothesis is not supported by geological data.
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MuRi | City Museum of Ricadi, Via Roma, 12, 89866 Santa Domenica, Vibo Valentia, Italy |
| SiMU | Museal System of the University of Calabria, Section of Palaeontology, Via Pietro Bucci 87036 Rende, Cosenza, Italy |
| MuMe | “Museo della Memoria”, Via Trento 2, Parghelia, Vibo Valentia, Italy |
| GPT | “Gruppo Paleontologico Tropeano”, last address at the “Civico Museo del Mare di Tropea”, vico Ospedale n. 1 89861—Tropea (VV) Vibo Valentia, Italy |
References
- Gramigna, P.; Guido, A.; Mastandrea, A.; Russo, F. The Paleontological Site of Cessaniti: A Window on a Coastal Marine Environment of Seven Million Years Ago (Southern Calabria, Italy). Geol. Romana 2008, 41, 25–34. [Google Scholar]
- Marra, A.C.; Carone, G.; Agnini, C.; Ghinassi, M.; Oms, O.; Rook, L. Stratigraphic and Chronologic Framework of the Upper Miocene Cessaniti Succession (Vibo Valentia, Calabria, Italy). Riv. Ital. Paleontol. Stratigr. 2017, 127, 379–393. [Google Scholar]
- Checchia-Rispoli, G. Illustrazione Dei Clipeastri Miocenici Della Calabria Seguita Da Uno Studio Sulla Morfologia Interna e Sulla Classificazione Dei Clipeastri. In Parte 3 di Memorie per Servire alla Descrizione della Carta Geologica d'Italia; Tipografia della Pace E. Cuggiani: Roma, Italy, 1925; Volume 9. [Google Scholar]
- Carone, G.; Domning, D.P. Metaxitherium serresii (Mammalia: Sirenia): New Pre-Pliocene Record, and Implications for Mediterranean Paleoecology before and after the Messinian Salinity Crisis. Boll. Soc. Paleontol. Ital. 2007, 46, 55. [Google Scholar]
- Guido, A.; Marra, A.C.; Mastandrea, A.; Tosti, F.; Russo, F. Micromorphological, Geochemical, and Diagenetic Characterization of Sirenian Ribs Preserved in the Late Miocene Paleontological Site of Cessaniti (Southern Calabria, Italy). Facies 2012, 58, 179–190. [Google Scholar] [CrossRef]
- Ferretti, M.P.; Rook, L.; Torre, D. Stegotetrabelodon (Proboscidea, Elephantidae) from the Late Miocene of Southern Italy. J. Vertebr. Paleontol. 2003, 23, 659–666. [Google Scholar] [CrossRef]
- Ferretti, M.P.; Rook, L.; Carone, G.; Marra, A.C. New Findings of Stegotetrabelodon syrticus from the Late Miocene of Cessaniti, Southern Italy. Boll. Soc. Paleontol. Ital. 2017, 56, 89–92. [Google Scholar]
- Ferretti, M.P. Miocene Proboscideans from Italy: African Elements and Palaeogeographic Implications. Geol. East Libya 2008, 3, 325–334. [Google Scholar]
- Pandolfi, L.; Marra, A.C.; Carone, G.; Maiorino, L.; Rook, L. A New Rhinocerotid (Mammalia, Rhinocerotidae) from the Latest Miocene of Southern Italy. Hist. Biol. 2021, 33, 194–208. [Google Scholar] [CrossRef]
- Marra, A.C. Out of Pikermi: The Occurrence of Bohlinia in the Late Miocene of the Central Mediterranean. Geosciences 2025, 15, 44. [Google Scholar] [CrossRef]
- Marra, A.C.; Solounias, N.; Carone, G.; Rook, L. Palaeogeographic Significance of the Giraffid Remains (Mammalia, Arctiodactyla) from Cessaniti (Late Miocene, Southern Italy). Geobios 2011, 44, 189–197. [Google Scholar] [CrossRef]
- Marra, A.C. Tragoportax cf. rugosifrons (Schlosser, 1904) from the Late Miocene of Cessaniti (Southern Italy). Comptes Rendus Palevol. 2018, 17, 378–387. [Google Scholar]
- Marra, A.C. Contribution of the Late Miocene Mammals from Calabria and Sicily to the Palaeogeography of the Central Mediterranean. Atti Della Accad. Pelorit. Pericolanti Cl. Sci. Fis. Mat. Nat. 2019, 97, 29. [Google Scholar]
- Kostopoulos, D. The Late Miocene Mammal Faunas of the Mytilinii Basin, Samos Island, Greece: New Collection 13. Giraffidae. Beitr. Zur Palaeontol. 2009, 31, 299–343. [Google Scholar]
- Solounias, N. Putting Samotherium in Its Place: The Morphology of Giraffids and the Geology of Samos; Cambridge Scholars Publishing: Newcastle upon Tyne, UK, 2024; ISBN 9781036404147. [Google Scholar]
- Witmer, L. The Paleoecology of the Pikermian Biome and the Savanna Myth. In Hominoid Evolution and Climatic Change in Europe: Volume 1, The Evolution of Neogene Terrestrial Ecosystems in Europe; Solounias, N., Plavcan, J.M., Quade, J., Witmer, L., Eds.; Cambridge University Press: Cambridge, UK, 1999; Volume 1. [Google Scholar]
- Solounias, N.; Rivals, F.; Semprebon, G.M. Dietary Interpretation and Paleoecology of Herbivores from Pikermi and Samos (Late Miocene of Greece). Paleobiology 2010, 36, 113–136. [Google Scholar] [CrossRef]
- Kostopoulos, D.S. The Pikermian Event: Temporal and Spatial Resolution of the Turolian Large Mammal Fauna in SE Europe. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 274, 82–95. [Google Scholar] [CrossRef]
- Mayor, A. The First Fossil Hunters: Paleontology in Greek and Roman Times; Princeton University Press: Princeton, NJ, USA, 2000; ISBN 0691089779. [Google Scholar]
- Mayor, A. The ‘Monster of Troy’ Vase: The Earliest Artistic Record of a Vertbrate Fossil Discovery? Oxf. J. Archaeol. 2000, 19, 57–63. [Google Scholar] [CrossRef]
- Monge-Nájera, J. Evaluation of the Hypothesis of the Monster of Troy Vase as the Earliest Artistic Record of a Vertebrate Fossil. Uniciencia 2020, 34, 147–151. [Google Scholar] [CrossRef]
- Solounias, N.; Mayor, A. Ancient References to the Fossils from the Land of Pythagoras. Earth Sci. Hist. 2004, 23, 283–296. [Google Scholar] [CrossRef]
- Nicotera, P. Rilevamento Geologico Del Versante Settentrionale Del Monte Poro (Calabria). Mem. Note Dell’istituto Geol. Appl. Napoli 1959, 7, 1–92. [Google Scholar]
- Marra, A.C.; Somma, R.; Guido, A. The Palaeontological Heritage of the Capo Vaticano–Monte Poro Area (Vibo Valentia, Italy): Research, Protection, and Management. Acta IMEKO 2023, 12, 1–11. [Google Scholar] [CrossRef]
- Carone, G.; Domning, D.P.; Marra, A.C. New Finds of Metaxytherium serresii (Gervais, 1847) (Mammalia: Sirenia) from the Upper Miocene of Monte Poro (Calabria, Italy). Boll. Della Soc. Paleontol. Ital. 2013, 52, 187–196. [Google Scholar]
- Gramigna, P.; Bassi, D.; Russo, F. An Upper Miocene Siliciclastic-Carbonate Ramp: Depositional Architecture, Facies Distribution, and Diagenetic History (Capo Vaticano Area, Southern Italy). Facies 2012, 58, 191–215. [Google Scholar] [CrossRef]
- Gramigna, P.; Morsilli, M.; Russo, F. Facies Changes in Siliciclastic-Carbonate Depositional Environments of the M.te Poro Area (Upper Miocene, Southern Calabria). Rend. Online Soc. Geol. Ital. 2008, 2, 93–96. [Google Scholar]
- Caracciolo, L.; Gramigna, P.; Critelli, S.; Calzona, A.B.; Russo, F. Petrostratigraphic Analysis of a Late Miocene Mixed Siliciclastic-Carbonate Depositional System (Calabria, Southern Italy): Implications for Mediterranean Paleogeography. Sediment. Geol. 2013, 284–285, 117–132. [Google Scholar] [CrossRef]
- Marra, A.C.; Sudano, F.; Rao, A.; Calzona, V.; Guido, A. Promotion of Paleontological Heritage: Case Histories from Southern Italy. Heritage 2025, 8, 100. [Google Scholar] [CrossRef]
- Von Den Driesch, A. A Guide to the Measurement of Animal Bones from Archaeological Sites; Peabody Museum Press: Cambridge, MA, USA, 1976; Volume 1, ISBN 0873659503. [Google Scholar]
- Bohlin, B.; Sefve, I.; Ringström, T.J.; Zdansky, O. Die Familie Giraffidae Mit Besonderer Berücksichtigung Der Fossilen Formen Aus China; Geological Survey of China: Beijing, China, 1926; Volume 4. [Google Scholar]
- Kostopoulos, D.; Koufos, G.; Koliadimou, K. The Giraffids (Mammalia, Artiodactyla) from the Late Miocene Mammalian Localities of Nikiti (Macedonia, Greece; Palaeontogr. Abt. A 1996, 239, 61–88. [Google Scholar]
- Koufos, G.D.; Kostopoulos, D.S.; Vlachou, T.D. The Late Miocene Mammal Faunas of the Mytilinii Basin, Samos Island, Greece: New Collection. 16. Biochronology. Beitr. Zur Paläontol. 2009, 31, 397–408. [Google Scholar]
- Ríos, M.; Danowitz, M.; Solounias, N. First Comprehensive Morphological Analysis on the Metapodials of Giraffidae. Palaeontol. Electron. 2016, 19.3.50A, 1–39. [Google Scholar] [CrossRef]
- Solounias, N.; Danowitz, M. Astragalar Morphology of Selected Giraffidae. PLoS ONE 2016, 11, e0151310. [Google Scholar] [CrossRef]
- Roussiakis, S.; Iliopoulos, G. Preliminary Observations on the Metrical Variation of Helladotherium duvernoyi and Bohlinia a Attica. In Proceedings of the 5th International Symposium on Eastern Mediterranean Geology, Thessaloniki, Greece, 14–20 April 2004; pp. 343–346. [Google Scholar]
- Xafis, A.; Mayda, S.; Grímsson, F.; Nagel, D.; Kaya, T.; Halaçlar, K. Fossil Giraffidae (Mammalia, Artiodactyla) from the Early Turolian of Kavakdere (Central Anatolia, Turkey). Comptes Rendus Palevol. 2019, 18, 619–642. [Google Scholar] [CrossRef]
- Geraads, D. Remarques Sur La Systématique et La Phylogénie Des Giraffidae (Artiodactyla, Mammalia). Geobios 1986, 19, 465–477. [Google Scholar] [CrossRef]
- Geraads, D. Les Giraffinae (Artiodactyla, Mammalia) Du Miocène Supérieur de La Région de Thessalonique (Grèce). Bull. Muséum Natl. D’histoire Nat. Sect. C Sci. Terre Paléontol. Géol. Minéral. 1979, 1, 377. [Google Scholar]
- Geraads, D.; Spassov, N.; Kovachev, D. Giraffidae (Artiodactyla, Mammalia) from the Late Miocene of Kalimantsi and Hadjidimovo, Southwestern Bulgaria. Geol. Balc. 2005, 35, 11–18. [Google Scholar] [CrossRef]
- Geraads, D. Les Gisements de Mammifères Du Miocène Supérieur de Kemiklitepe, Turquie: 8. Giraffidae. Bull. Muséum Natl. D’histoire Nat. Sect. C Sci. Terre Paléontol. Géol. Minéral. 1994, 16, 159. [Google Scholar]
- Gentry, A.W.; Rössner, G.E.; Heizmann, E.P.J. Suborder Ruminantia. Miocene Land Mamm. Eur. 1999, 225, 258. [Google Scholar]
- Bernor, R.L.; Rook, L. A Current View of Sahabi Large Mammal Biogeographic Relationships. Garyounis Sci. Bull. Spec. Issue 2008, 5, 285–292. [Google Scholar]
- Sanders, W.J. Proboscidea from the Baynunah Formation. In Sands of Time: Ancient Life in the Late Miocene of Abu Dhabi, United Arab Emirates; Springer: Berlin/Heidelberg, Germany, 2022; pp. 141–177. [Google Scholar]
- Bibi, F.; Kaya, F.; Varela, S. Paleoecology and Paleobiogeography of the Baynunah Fauna. In Sands of Time: Ancient Life in the Late Miocene of Abu Dhabi, United Arab Emirates; Springer: Berlin/Heidelberg, Germany, 2022; pp. 333–350. [Google Scholar]
- Whybrow, P.J.; Hill, A.P. Fossil Vertebrates of Arabia: With Emphasis on the Late Miocene Faunas, Geology, and Palaeoenvironments of the Emirate of Abu Dhabi, United Arab Emirates; Yale University Press: New Haven, CT, USA, 1999. [Google Scholar]
- Bibi, F. Bovidae and Giraffidae from the Baynunah Formation. In Sands of Time: Ancient Life in the Late Miocene of Abu Dhabi, United Arab Emirates; Springer: Berlin/Heidelberg, Germany, 2022; pp. 219–241. [Google Scholar]
- Solounias, N.; McGraw, W.S.; Hayek, L.-A.; Werdelin, L. The Paleodiet of Giraffidae. In Antelopes, Deer and Relatives; Yale University Press: New Haven, CT, USA, 2000. [Google Scholar]
- Ríos, M.; Sánchez, I.M.; Morales, J. A New Giraffid (Mammalia, Ruminantia, Pecora) from the Late Miocene of Spain, and the Evolution of the Sivathere-Samothere Lineage. PLoS ONE 2017, 12, e0185378. [Google Scholar] [CrossRef]
- Spassov, N.; Geraads, D. Tragoportax PILGRIM, 1937 and Miotragocerus STROMER, 1928 (Mammalia, Bovidae) from the Turolian of Hadjidimovo, Bulgaria, and a Revision of the Late Miocene Mediterranean Boselaphini. Geodiversitas 2004, 26, 339. [Google Scholar]
- Eronen, J.T.; Ataabadi, M.M.; Micheels, A.; Karme, A.; Bernor, R.L.; Fortelius, M. Distribution History and Climatic Controls of the Late Miocene Pikermian Chronofauna. Proc. Natl. Acad. Sci. USA 2009, 106, 11867–11871. [Google Scholar] [CrossRef]
- Olson, E.C. The Evolution of a Permian Vertebrate Chronofauna. Evolution 1952, 6, 181–196. [Google Scholar] [CrossRef]
- Ataabadi, M.M.; Bernor, R.L.; Kostopoulos, D.S.; Wolf, D.; Orak, Z.; Zare, G.; Nakaya, H.; Watabe, M.; Fortelius, M. Chapter 25. Recent Advances in Paleobiological Research of the Late Miocene Maragheh Fauna, Northwest Iran. In Fossil Mammals of Asia: Neogene Biostratigraphy and Chronology; Columbia University Press: New York, NY, USA, 2013; pp. 546–565. [Google Scholar]
- Eronen, J.T.; Fortelius, M.; Micheels, A.; Portmann, F.T.; Puolamäki, K.; Janis, C.M. Neogene Aridification of the Northern Hemisphere. Geology 2012, 40, 823–826. [Google Scholar] [CrossRef]
- Kurtén, B. The Chinese Hipparion Fauna: A Quantitative Survey with Comments on the Ecology of the Machairodonts and Hyaenids and the Taxonomy of the Gazelles; Munksgaard: Copenhagen, Denmark, 1952. [Google Scholar]
- Casanovas-Vilar, I.; García-Paredes, I.; Alba, D.M.; van den Hoek Ostende, L.W.; Moyà-Solà, S. The European Far West: Miocene Mammal Isolation, Diversity and Turnover in the Iberian Peninsula. J. Biogeogr. 2010, 37, 1079–1093. [Google Scholar] [CrossRef]
- Bibi, F. Mio-Pliocene Faunal Exchanges and African Biogeography: The Record of Fossil Bovids. PLoS ONE 2011, 6, e16688. [Google Scholar] [CrossRef] [PubMed]
- Böhme, M.; Spassov, N.; Majidifard, M.R.; Gärtner, A.; Kirscher, U.; Marks, M.; Dietzel, C.; Uhlig, G.; El Atfy, H.; Begun, D.R. Neogene Hyperaridity in Arabia Drove the Directions of Mammalian Dispersal between Africa and Eurasia. Commun. Earth Environ. 2021, 2, 85. [Google Scholar] [CrossRef]
- Cirrincione, R.; Fazio, E.; Fiannacca, P.; Ortolano, G.; Pezzino, A.; Punturo, R. The Calabria-Peloritani Orogen, a Composite Terrane in Central Mediterranean; Its Overall Architecture and Geodynamic Significance for a Pre-Alpine Scenario around the Tethyan Basin. Period. Mineral. 2015, 84, 701–749. [Google Scholar]
- Russo, D.; Fiannacca, P.; Fazio, E.; Cirrincione, R.; Mamtani, M.A. Geology and Microstructural Evolution of Syn-to Late-Tectonic Granitoids from Capo Vaticano Promontory (Southern Calabria, Italy). Geol. Mag. 2025, 162, e8. [Google Scholar] [CrossRef]
- Rook, L.; Gallai, G.; Torre, D. Lands and Endemic Mammals in the Late Miocene of Italy: Constrains for Paleogeographic Outlines of Tyrrhenian Area. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2006, 238, 263–269. [Google Scholar] [CrossRef]
- Rook, L. Geopalaeontological Setting, Chronology and Palaeoenvironmental Evolution of the Baccinello-Cinigiano Basin Continental Successions (Late Miocene, Italy). Comptes Rendus Palevol 2016, 15, 825–836. [Google Scholar] [CrossRef]
- Cirilli, O.; Benvenuti, M.; Carnevale, G.; Casanovas Vilar, I.; Delfino, M.; Furio, M.; Papini, M.; Villa, A.; Rook, L. Fosso Della Fittaia: The Oldest Tusco-Sardinian Late Miocene Endemic Vertebrate Assemblage (Baccinello-Cinigiano Basin, Tuscany, Italy). Riv. Ital. Paleontol. E Stratigr. 2016, 122, 13–34. [Google Scholar]
- Abbazzi, L.; Delfino, M.; Gallai, G.; Trebini, L.; Rook, L. New Data on the Vertebrate Assemblage of Fiume Santo (North-West Sardinia, Italy), and Overview on the Late Miocene Tusco-Sardinian Palaeobioprovince. Palaeontology 2008, 51, 425–451. [Google Scholar] [CrossRef]
- van den Hoek Ostende, L.W.; Meijer, H.J.M.; van der Geer, A.A.E. A Bridge Too Far. Comment on “Processes of Island Colonization by Oligo-Miocene Land Mammals in the Central Mediterranean: New Data from Scontrone (Abruzzo, Central Italy) and Gargano (Apulia, Southern Italy)” by PPA Mazza and M. Rustioni [Palaeogeography, Palaeoclimatology, Palaeoecology 267 (2008) 208–215]. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 279, 128–130. [Google Scholar]
- Mazza, P.P.A.; Rustioni, M. Five New Species of Hoplitomeryx from the Neogene of Abruzzo and Apulia (Central and Southern Italy) with Revision of the Genus and of Hoplitomeryx Matthei Leinders, 1983. Zool. J. Linn. Soc. 2011, 163, 1304–1333. [Google Scholar] [CrossRef]
- Mazza, P.P.A.; Rustioni, M. Processes of Island Colonization by Oligo–Miocene Land Mammals in the Central Mediterranean: New Data from Scontrone (Abruzzo, Central Italy) and Gargano (Apulia, Southern Italy). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 267, 208–215. [Google Scholar] [CrossRef]
- Freudenthal, M.; van den Hoek Ostende, L.W.; Martín-Suárez, E. When and How Did the Mikrotia Fauna Reach Gargano (Apulia, Italy)? Geobios 2013, 46, 105–109. [Google Scholar] [CrossRef]
- Seguenza, G. Nuovi Resti Di Mammiferi Pontici Di Gravitelli Presso Messina. Boll. Della Soc. Geogr. Ital. 1907, 26, 89–122. [Google Scholar]
- Seguenza, L. I Vertebrati Fossili Della Provincia Di Messina: Parte II. Mammiferi e Geologia Del Piano Pontico. Boll. Della Soc. Geol. Ital. 1902, 21, 115–172. [Google Scholar]
- Iannucci, A. A Reappraisal of the Lost Suids from the Late Miocene of Gravitelli (Sicily, Italy) and Paleobiogeographical Implications. Palaeoworld 2024, 33, 469–491. [Google Scholar] [CrossRef]
- Gallai, G.; Rook, L. Propotamochoerus sp. (Suidae, Mammalia) from the Late Miocene of Gravitelli (Messina, Sicily, Italy) Rediscovered. Riv. Ital. Paleontol. Stratigr. 2006, 112, 317. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specimen | Locality | Inventory Number |
|---|---|---|
| Samotherium boissieri | ||
| Ulna—left proximal epiphysis | Malopara | MuRi-870 |
| Radius—left distal epiphysis | Malopara | MuRi-895 |
| Radius—right distal epiphysis | Malopara | MuRi-866 |
| Scaphoid—left | Malopara | MuRi-878 |
| Intermedium—left | Malopara | MuRi-879 |
| Intermedium—right | Malopara | MuRi-885 |
| Ulnare—left | Malopara | MuRi-880 |
| Ulnare—right | Malopara | MuRi-884 |
| Magnotrapezoid—left | Malopara | MuRi-882 |
| Unicinatum—left | Malopara | MuRi-881 |
| Unicinatum—right | Malopara | MuRi-886 |
| Sesamoids and pisiform | Malopara | MuRi-883 |
| Metacarpal—left | Malopara | MuRi-865 |
| Metacarpal—right | Malopara | MuRi-864 |
| Metacarpal. distal end—left | Cessaniti | MuRi-875 |
| First phalanx, III digit—right | Malopara | MuRi-888 |
| First phalanx, IV digit—right | Malopara | MuRi-887 |
| Second phalanx, III digit—right | Malopara | MuRi-890 |
| Second phalanx, IV digit—right | Malopara | MuRi-899 |
| Third phalanx, III digit—right | Malopara | MuRi-892 |
| Third phalanx, IV digit—right | Malopara | MuRi-891 |
| Third phalanges III and IV digit | Cessaniti ? | MuRi-894 |
| Astragalus—right | Cessaniti | ? GPT-MuMe |
| Metatarsal—left | Cessaniti | SiMU-CMA07 |
| Metatarsal—right | Cessaniti | SiMU-CMA06 |
| Bohlinia attica | ||
| Radius—left | Cessaniti | MuRi-874 |
| Metacarpal—left | Cessaniti | MuRi-972 |
| Astragalus—left | Cessaniti | SiMU-CMA02 |
| Astragalus—right | Cessaniti | Muri-877 |
| Inventory | L | TDp | APDp | TDdia | CD | APDdia | TDd | APDd |
|---|---|---|---|---|---|---|---|---|
| MuRi-865 | 373 | 75.7 | 48.2 | 43.7 | 136.5 | 36.5 | 84.1 | 48.2 |
| MuRi-864 | 373 | 74.4 | 49.2 | 44 | 136.5 | 36.3 | 82.8 | 48.0 |
| MuRi-875 | - | - | - | - | - | - | 83.4 | 48.0 |
| MuRi-972 | - | 90.8 | 53.1 | - | - | - | 88.0 | 49.9 |
| Specimen | Inventory | L | TDp | SD | TDd |
|---|---|---|---|---|---|
| First phalanx, III digit | MuRi-888 | 100.9 | 41.1 | 32.9 | 32.2 |
| First phalanx, IV digit | MuRi-887 | 98.0 | 40.1 | 32.7 | 31.4 |
| Second phalanx, III digit | MuRi-890 | 65.2 | 37.7 | 30.6 | 32.2 |
| Second phalanx, IV digit | MuRi-899 | 61.3 | 36.9 | 30.9 | 31.0 |
| DLS | Ld | MBs | |||
| Third phalanx, III digit | MuRi-892 | 80.7 | 59.1 | 30.5 | |
| Third phalanx, IV digit | MuRi-891 | 80.5 | 61.7 | 29.3 |
| Inventory | L | TDp | APDp | TDdia | CD | APDdia | TDd | APDd |
|---|---|---|---|---|---|---|---|---|
| SiMU-CMA06 | 395 | 69.9 | 65.9 | 41.0 | 15.0 | 33.6 | 75.8 | 46.0 |
| SiMU-CMA07 | 405.0 | 78.0 | 72.0 | 43.0 | 15.5 | 37.0 | 91.3 | 50.3 |
| Species | Ref. | Llat | Lmed | TD | APDd |
|---|---|---|---|---|---|
| Cessaniti specimen | 90 | 80 | 58 | 54 | |
| Samotherium boissieri | |||||
| [14] | 90.08 | 78.5 | 57.6 | ||
| [14] | 96.5 | 82.3 | 59.7 | ||
| [14] | 97.5 | 85.1 | 58.4 | ||
| [14] | 89.2 | 78.6 | 59.3 | ||
| [14] | 89.0 | 79.2 | 58.0 | ||
| [14] | 88.4 | 78.0 | 59 | ||
| [14] | 86.8 | 73.7 | 56.4 | ||
| [14] | 85.6 | 75.0 | 56.8 | ||
| [14] | 89.3 | 76.2 | 61.0 | ||
| [14] | 84.4 | 76.5 | 53.5 | ||
| [14] | 88.8 | 77.6 | 55.0 | ||
| [14] | 89.0 | 75.8 | 59.0 | ||
| [14] | 86.0 | 77.2 | 55.7 | ||
| [14] | 90.0 | 76.6 | 56.0 | ||
| [14] | 92.2 | 81.0 | 62.3 | ||
| [37] | 88.1 | 80.95 | 56.16 | ||
| Bohlinia attica | |||||
| [10] | 89 | 78.6 | 59 | 55.6 | |
| [10] | 81 | 59 | |||
| [36] | 87.7–103.4 (min–max) | 76.5–89.6 (min–max) | 55–68 (min–max) | 46.5–61.1 (min–max) | |
| [36] | 103 | 99 | 76.6 | ||
| [36] | 105 | 94 | 70 | ||
| [36] | 105 | 91 | 71 | ||
| [36] | 108 | 94 | 73 | ||
| [42] | 83.8 | 59.3 | |||
| [42] | 87.6 | ||||
| [39] | 81 | 59 |
| Stegotetrabelodon syrticus | cf. Ceratotherium advenientis | Tragoportax cf. rugosifrons | Bohlinia attica | Samotherium boissieri | |
|---|---|---|---|---|---|
| Unit 2 | mandible; incisor; fragmentary molar; two fragmentary humeri *; right II metacarpal; incomplete femur | fragmentary skull; two fragmentary teeth; few postcranial bones | hemimandible; radius; astragalus; anterior phalanx; humerus *; anteriorphalanx *; metatarsal *; posterior phalanx * | upper molar row distal radius; metacarpal; two astragali; two cubonaviculars; | two metacarpals two metatarsals complete manus ** fragmentary ulna ** distal radius ** fragmentary humerus ** |
| Unit 1 | one worn DP4 |
| Locality | Country | Age (Ma) | Samotherium boissieri | Bohlinia attica | Tragoportax rugosifrons |
|---|---|---|---|---|---|
| Azmaka | Bulgaria | 7.2 | X | ||
| Gorna Susica | Bulgaria | 8.3–7.285 | X | ||
| Hadjidimovo | Bulgaria | 7.6–7.1 | X | X | |
| Kalimantsi | Bulgaria | 7.6–7.1 | X | ||
| Krodimovo | Bulgaria | 7.6–7.1 | X | ||
| Strumyani | Bulgaria | 7.6–7.1 | X | X | |
| Kirokuçuk | N Macedonia | 8.9–5.3 | X | ||
| Dolni Disan | N Macedonia | 8.9–5.3 | X | ||
| Karaslari | N Macedonia | 7.6–7.1 | X | X | |
| Ditiko | Greece | 7.1–5.3 | X | ||
| Kerassia | Greece | 8.9–7.1 | X | ||
| Nikiti | Greece | 9.9–8.9 | X | ||
| Pikermi * | Greece | 7.454–7.1 | X | ||
| Pyrgos Vassilissis | Greece | 7.212–7.14 | X | ||
| Ravin de la Pluie | Greece | 9.426–9.311 | X | ||
| Ravin Zouaves * | Greece | 9.9–8.1 | X | ||
| Samos * | Greece | 8.9–5.3 | X | X | |
| Vathylakkos * | Greece | 7.489–7.454 | X | ||
| Corakyerler | Turkey | 8.9–7.6 | X | X | |
| Esendere | Turkey | 8.9–7.6 | X | ||
| Küçükçekmece | Turkey | 8.9–5.3 | X | ||
| Gülpinar | Turkey | 7.6–7.1 | X | ||
| Karain | Turkey | 8.9–7.1 | X | ||
| Kavakdere | Turkey | 8.254–8.108 | X | ||
| Sinap * | Turkey | 11.2–7.1 | X | ||
| Injana | Iraq | 8.9–7.6 | X | X | |
| Maragheh * | Iran | 8.9–7.1 | X | X | |
| Novo-Elizavetovka | Ukraine | 8–9–7.1 | X | ||
| Taraklia | Moldova | 7.6–7.1 | X | X |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marra, A.C. Samotherium boissieri from the Late Miocene of Southern Italy. Life 2025, 15, 911. https://doi.org/10.3390/life15060911
Marra AC. Samotherium boissieri from the Late Miocene of Southern Italy. Life. 2025; 15(6):911. https://doi.org/10.3390/life15060911
Chicago/Turabian StyleMarra, Antonella Cinzia. 2025. "Samotherium boissieri from the Late Miocene of Southern Italy" Life 15, no. 6: 911. https://doi.org/10.3390/life15060911
APA StyleMarra, A. C. (2025). Samotherium boissieri from the Late Miocene of Southern Italy. Life, 15(6), 911. https://doi.org/10.3390/life15060911