

Clock Proteins Have the Potential to Improve Term Delivery Date Prediction: A Proof-of-Concept Study

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Study Design and Participants
2.3. Proteomic Analysis
2.4. Statistical Analyses
3. Results
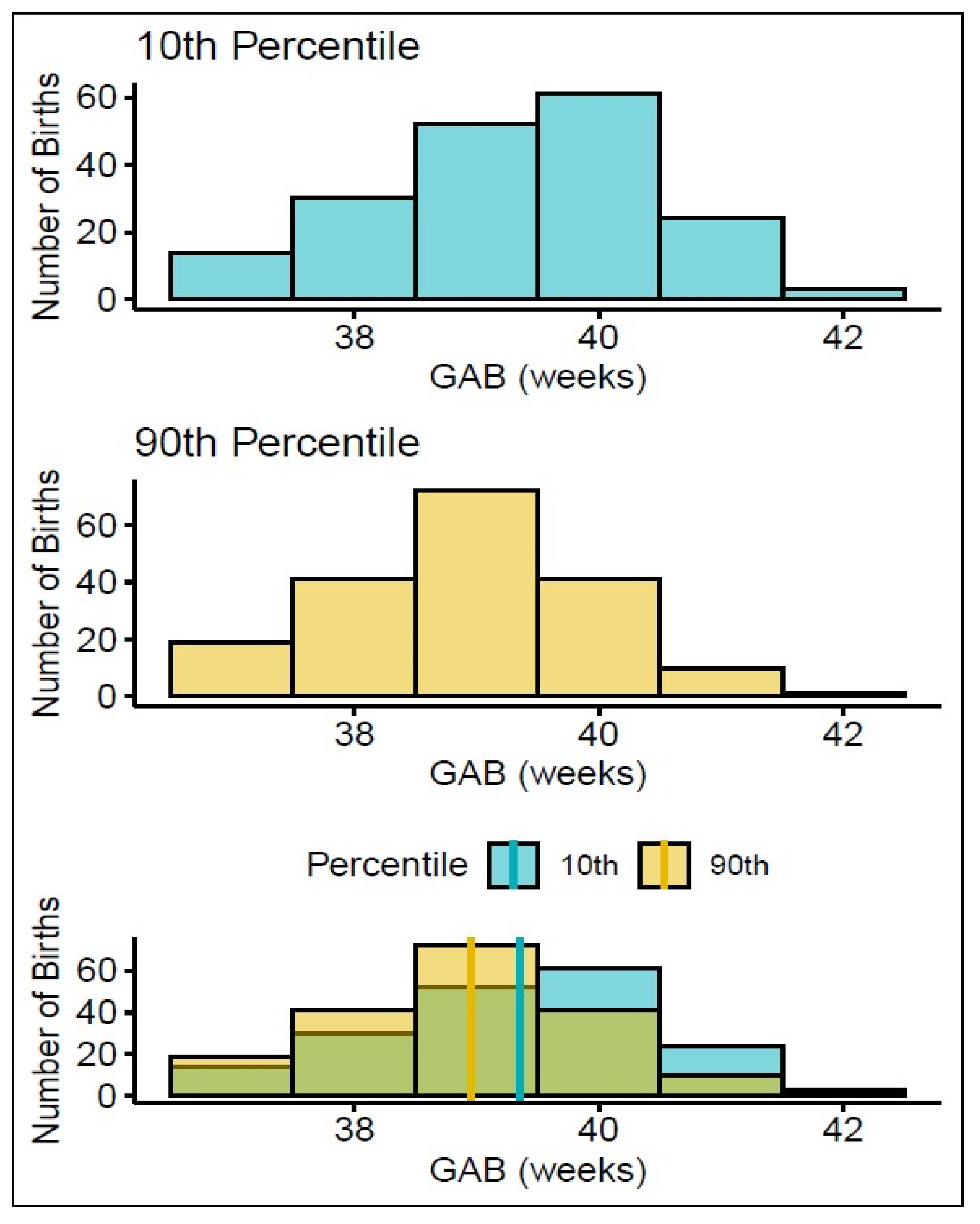
3.1. Patient Characteristics
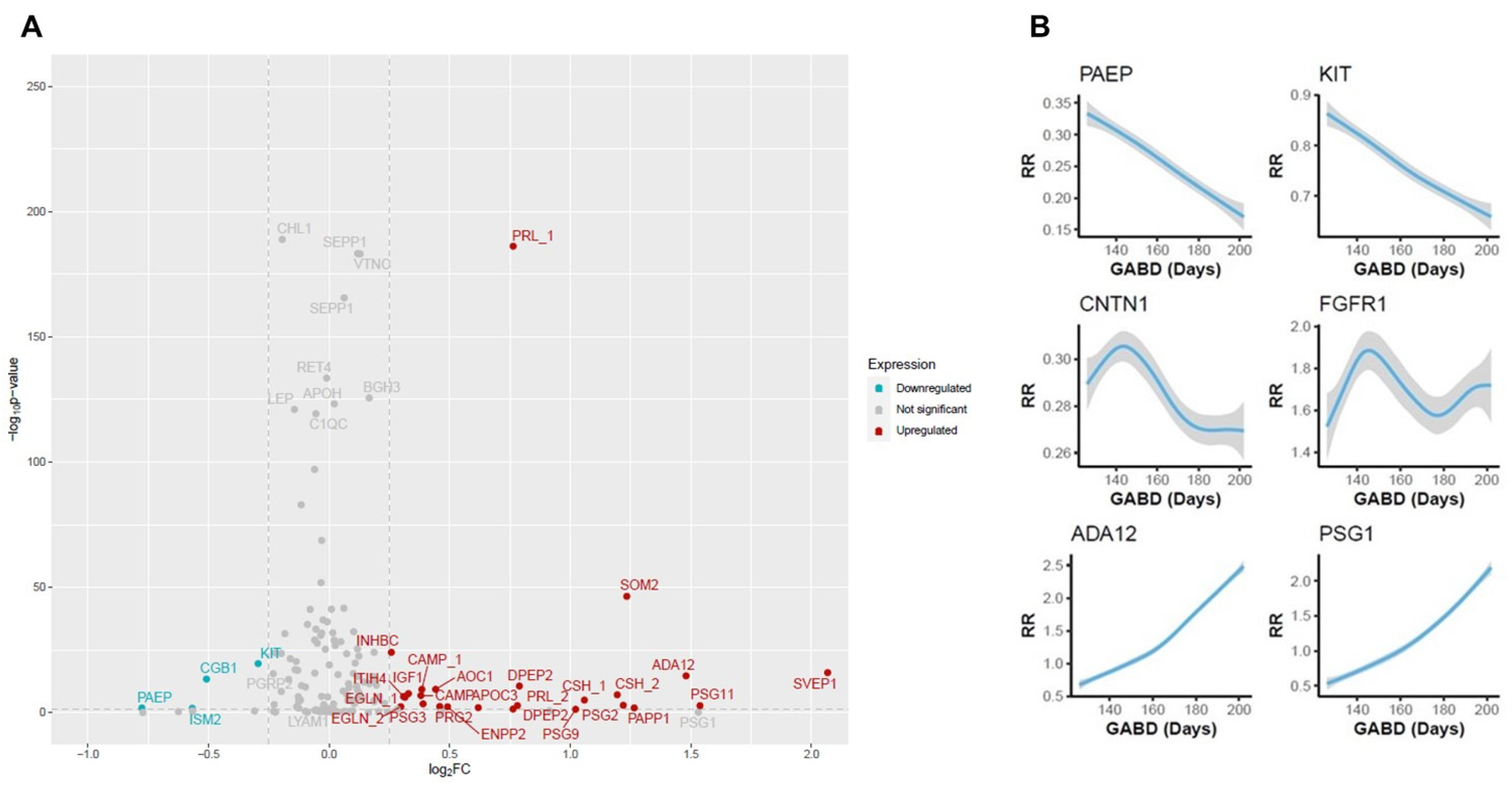
3.2. Clock Protein Expression
3.3. Association Between Proteins and Time to Birth
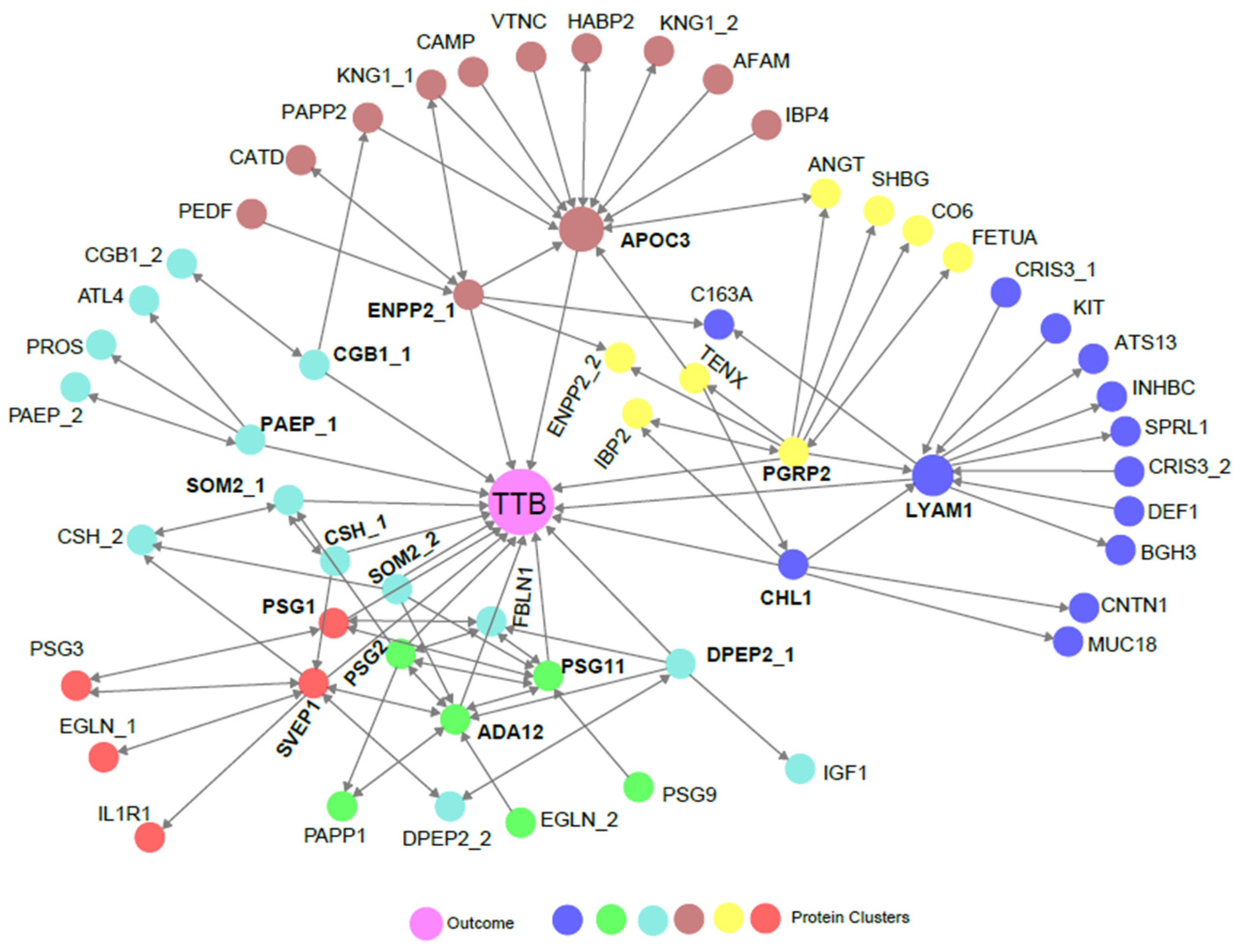
3.4. Biological Association of Biomarkers with TTB
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ADA12 | Disintegrin and metalloproteinase domain-containing protein 12 |
| AFAM | Afamin |
| ANGT | Angiotensinogen |
| AOC1 | Amiloride-sensitive amine oxidase [copper-containing] |
| APOC3 | Apolipoprotein C-III |
| APOH | Beta-2-glycoprotein 1 |
| ATL4 | ADAMTS-like protein 4 |
| ATS13 | A disintegrin and metalloproteinase with thrombospondin motifs 13 |
| BGH3 | Transforming growth factor-beta-induced protein ig-h3 |
| BMI | Body mass index |
| C163A | Scavenger receptor cysteine-rich type 1 protein M130 |
| C1QC | Complement C1q subcomponent subunit C |
| CAMP | Cathelicidin antimicrobial peptide |
| CAP | College of American Pathologists |
| CATD | Cathepsin D |
| CGB1 | Choriogonadotropin subunit beta variant 1 |
| CHL1 | Neural cell adhesion molecule L1-like protein |
| CLIA | Clinical Laboratory Improvement Amendment |
| CNTN1 | Contactin-1 |
| CO6 | Complement component C6 |
| CRIS3 | Cysteine-rich secretory protein 3 |
| CSH | Chorionic somatomammotropin hormone 1 |
| CSH | Chorionic somatomammotropin hormone 2 |
| DEF1 | Neutrophil defensin 1 |
| DPEP2 | Dipeptidase 2 |
| EDD | Estimated delivery date |
| EGLN | Endoglin |
| ENPP2 | Ectonucleotide pyrophosphatase/phosphodiesterase family member 2 |
| FBLN1 | Fibulin-1 |
| FETUA | Alpha-2-HS-glycoprotein |
| HABP2 | Hyaluronan-binding protein 2 |
| GAB | Gestational age at birth |
| GABD | Gestational age at blood draw |
| IBP2 | Insulin-like growth factor-binding protein 2 |
| IBP4 | Insulin-like growth factor-binding protein 4 |
| IGF1 | Insulin-like growth factor I |
| IL1R | Interleukin-1 receptor type 1 |
| INHBC | Inhibin beta C chain |
| ISM2 | Isthmin-2 |
| ITIH4 | Inter-alpha-trypsin inhibitor heavy chain H4 |
| KIT | Mast/stem cell growth factor receptor Kit |
| KNG1 | Kininogen-1 |
| LC-MRM-MS | Liquid chromatography–multiple reaction monitoring–mass spectrometry |
| LEP | Leptin |
| LYAM1 | L-selectin |
| MARS | Multiple affinity removal system |
| MR | Mendelian randomization |
| MRPC | Mendelian randomization-based machine learning algorithm |
| MRM | Multiple reaction monitoring |
| MUC18 | Cell surface glycoprotein MUC18 |
| PAEP | Glycodelin |
| PAPP1 | Pappalysin-1 |
| PAPP2 | Pappalysin-2 |
| PEDF | Pigment epithelium-derived factor |
| PGRP2 | n-acetylmuramoyl-L-alanine amidase |
| PRG2 | Bone marrow proteoglycan |
| PRL | Prolactin |
| PROS | Vitamin K-dependent protein S |
| PSG1 | Pregnancy-specific beta-1-glycoprotein 1 |
| PSG11 | Pregnancy-specific beta-1-glycoprotein 11 |
| PSG2 | Pregnancy-specific beta-1-glycoprotein 2 |
| PSG3 | Pregnancy-specific beta-1-glycoprotein 3 |
| PSG9 | Pregnancy-specific beta-1-glycoprotein 9 |
| RET4 | Retinol-binding protein 4 |
| RR | Response ratio |
| SEPP1 | Selenoprotein P |
| SHBG | Sex hormone-binding globulin |
| SOM2 | Growth hormone variant |
| SPRL1 | SPARC-like protein 1 |
| SIS | Stable isotope standard |
| SVEP1 | Sushi, von Willebrand factor type A, EGF and pentraxin domain-containing protein 1 |
| TENX | Tenascin-X |
| TTB | Time to birth |
| UHPLC | Ultra-high-performance liquid chromatography |
| VTNC | Vitronectin |
References
- Engle, W.A.; Kominiarek, M.A. Late preterm infants, early term infants, and timing of elective deliveries. Clin. Perinatol. 2008, 35, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Atkinson, J.; Roddy Mitchell, A.; Tong, S.; Walker, S.P.; Middleton, A.; Lindquist, A.; Hastie, R. Comparison of Maternal Labor-Related Complications and Neonatal Outcomes Following Elective Induction of Labor at 39 Weeks of Gestation vs Expectant Management: A Systematic Review and Meta-analysis. JAMA Netw. Open 2023, 6, e2313162. [Google Scholar] [CrossRef]
- Khasawneh, W.; Alyousef, R.; Akawi, Z.; Al-Dhoon, A.; Odat, A. Maternal and Perinatal Determinants of Late Hospital Discharge Among Late Preterm Infants; A 5-Year Cross-Sectional Analysis. Front. Pediatr. 2021, 9, 685016. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.; Siebert, K.; Carlson, S.; Ratner, N. Patient perspectives on loss of local obstetrical services in rural northern Minnesota. Birth 2018, 45, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Sonenberg, A.; Mason, D.J. Maternity Care Deserts in the US. JAMA Health Forum 2023, 4, e225541. [Google Scholar] [CrossRef] [PubMed]
- Tanne, J.H. Nearly six million women in the US live in maternity care deserts. BMJ 2023, 382, 1878. [Google Scholar] [CrossRef] [PubMed]
- Vos, S.C.; Anthony, K.E.; O’Hair, H.D. Constructing the uncertainty of due dates. Health Commun. 2014, 29, 866–876. [Google Scholar] [CrossRef] [PubMed]
- Naidu, K.; Fredlund, K.L. Gestational Age Assessment. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2025. [Google Scholar]
- Committee on Obstetric Practice American Institute of Ultrasound in Medicine Society for Maternal–Fetal Medicine. Committee Opinion No 700: Methods for Estimating the Due Date. Obstet Gynecol 2017, 129, e150–e154. [Google Scholar] [CrossRef]
- Osterman, M.J.K.; Hamilton, B.E.; Martin, J.A.; Driscoll, A.K.; Valenzuela, C.P. Births: Final Data for 2022. Natl. Vital Stat. Rep. 2024, 73, 1–56. [Google Scholar] [PubMed]
- Menon, R.; Bonney, E.A.; Condon, J.; Mesiano, S.; Taylor, R.N. Novel concepts on pregnancy clocks and alarms: Redundancy and synergy in human parturition. Hum. Reprod. Update 2016, 22, 535–560. [Google Scholar] [CrossRef]
- McLean, M.; Bisits, A.; Davies, J.; Woods, R.; Lowry, P.; Smith, R. A placental clock controlling the length of human pregnancy. Nat. Med. 1995, 1, 460–463. [Google Scholar] [CrossRef] [PubMed]
- Norwitz, E.R.; Bonney, E.A.; Snegovskikh, V.V.; Williams, M.A.; Phillippe, M.; Park, J.S.; Abrahams, V.M. Molecular Regulation of Parturition: The Role of the Decidual Clock. Cold Spring Harb. Perspect. Med. 2015, 5, a023143. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.S.; Stelzer, I.A.; Tsai, A.S.; Ghaemi, M.S.; Han, X.; Ando, K.; Winn, V.D.; Martinez, N.R.; Contrepois, K.; Moufarrej, M.N.; et al. Multiomic immune clockworks of pregnancy. Semin. Immunopathol. 2020, 42, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Aghaeepour, N.; Ganio, E.A.; McIlwain, D.; Tsai, A.S.; Tingle, M.; Van Gassen, S.; Gaudilliere, D.K.; Baca, Q.; McNeil, L.; Okada, R.; et al. An immune clock of human pregnancy. Sci. Immunol. 2017, 2, eaan2946. [Google Scholar] [CrossRef] [PubMed]
- Aghaeepour, N.; Lehallier, B.; Baca, Q.; Ganio, E.A.; Wong, R.J.; Ghaemi, M.S.; Culos, A.; El-Sayed, Y.Y.; Blumenfeld, Y.J.; Druzin, M.L.; et al. A proteomic clock of human pregnancy. Am. J. Obstet. Gynecol. 2018, 218, 347.e1–347.e14. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Rasmussen, M.H.; Piening, B.; Shen, X.; Chen, S.; Rost, H.; Snyder, J.K.; Tibshirani, R.; Skotte, L.; Lee, N.C.; et al. Metabolic Dynamics and Prediction of Gestational Age and Time to Delivery in Pregnant Women. Cell 2020, 181, 1680–1692.e15. [Google Scholar] [CrossRef] [PubMed]
- Ngo, T.T.M.; Moufarrej, M.N.; Rasmussen, M.H.; Camunas-Soler, J.; Pan, W.; Okamoto, J.; Neff, N.F.; Liu, K.; Wong, R.J.; Downes, K.; et al. Noninvasive blood tests for fetal development predict gestational age and preterm delivery. Science 2018, 360, 1133–1136. [Google Scholar] [CrossRef]
- Mitao, M.; Mwita, W.C.; Antony, C.; Adinan, H.; Shayo, B.; Amour, C.; Mboya, I.B.; Mahande, M.J. Recurrence of post-term pregnancy and associated factors among women who delivered at Kilimanjaro Christian Medical Centre in northern Tanzania: A retrospective cohort study. PLoS ONE 2023, 18, e0282078. [Google Scholar] [CrossRef]
- Lancaster, E.E.; Lapato, D.M.; Jackson-Cook, C.; Strauss, J.F., 3rd; Roberson-Nay, R.; York, T.P. Maternal biological age assessed in early pregnancy is associated with gestational age at birth. Sci. Rep. 2021, 11, 15440. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.M.; McPherson, N.O. Influence of increased paternal BMI on pregnancy and child health outcomes independent of maternal effects: A systematic review and meta-analysis. Obes. Res. Clin. Pract. 2019, 13, 511–521. [Google Scholar] [CrossRef]
- Jukic, A.M.; Baird, D.D.; Weinberg, C.R.; McConnaughey, D.R.; Wilcox, A.J. Length of human pregnancy and contributors to its natural variation. Hum. Reprod. 2013, 28, 2848–2855. [Google Scholar] [CrossRef]
- Mekonen, M.W.; Bayew, A.T.; Lakew, T.J. Factors affecting the duration of gestation among women taking prenatal care at Gondar referral hospital, Ethiopia. Health Sci. Rep. 2022, 5, e676. [Google Scholar] [CrossRef] [PubMed]
- Saade, G.R.; Boggess, K.A.; Sullivan, S.A.; Markenson, G.R.; Iams, J.D.; Coonrod, D.V.; Pereira, L.M.; Esplin, M.S.; Cousins, L.M.; Lam, G.K.; et al. Development and validation of a spontaneous preterm delivery predictor in asymptomatic women. Am. J. Obstet. Gynecol. 2016, 214, 633.e1–633.e24. [Google Scholar] [CrossRef] [PubMed]
- Bradford, C.; Severinsen, R.; Pugmire, T.; Rasmussen, M.; Stoddard, K.; Uemura, Y.; Wheelwright, S.; Mentinova, M.; Chelsky, D.; Hunsucker, S.W.; et al. Analytical validation of protein biomarkers for risk of spontaneous preterm birth. Clin. Mass Spectrom. 2017, 3, 25–38. [Google Scholar] [CrossRef]
- Davey Smith, G.; Hemani, G. Mendelian randomization: Genetic anchors for causal inference in epidemiological studies. Hum. Mol. Genet. 2014, 23, R89–R98. [Google Scholar] [CrossRef]
- Badsha, M.B.; Fu, A.Q. Learning Causal Biological Networks With the Principle of Mendelian Randomization. Front. Genet. 2019, 10, 460. [Google Scholar] [CrossRef]
- Badsha, M.B.; Martin, E.A.; Fu, A.Q. MRPC: An R Package for Inference of Causal Graphs. Front. Genet. 2021, 12, 651812. [Google Scholar] [CrossRef] [PubMed]
- Khanam, R.; Fleischer, T.C.; Boghossian, N.S.; Nisar, I.; Dhingra, U.; Rahman, S.; Fox, A.C.; Ilyas, M.; Dutta, A.; Naher, N.; et al. Performance of a validated spontaneous preterm delivery predictor in South Asian and Sub-Saharan African women: A nested case control study. J. Matern. Fetal Neonatal Med. 2022, 35, 8878–8886. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing, Version 4.4.2; R Foundation for Statistical Computing: Vienna, Austria, 2024. [Google Scholar]
- Chellakooty, M.; Vangsgaard, K.; Larsen, T.; Scheike, T.; Falck-Larsen, J.; Legarth, J.; Andersson, A.M.; Main, K.M.; Skakkebaek, N.E.; Juul, A. A longitudinal study of intrauterine growth and the placental growth hormone (GH)-insulin-like growth factor I axis in maternal circulation: Association between placental GH and fetal growth. J. Clin. Endocrinol. Metab. 2004, 89, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Hannibal, R.L.; Cardoso-Moreira, M.; Chetty, S.P.; Lau, J.; Qi, Z.; Gonzalez-Maldonado, E.; Cherry, A.M.; Yu, J.; Norton, M.E.; Baker, J.C. Investigating human placentation and pregnancy using first trimester chorionic villi. Placenta 2018, 65, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Korevaar, T.I.; Steegers, E.A.; de Rijke, Y.B.; Schalekamp-Timmermans, S.; Visser, W.E.; Hofman, A.; Jaddoe, V.W.; Tiemeier, H.; Visser, T.J.; Medici, M.; et al. Reference ranges and determinants of total hCG levels during pregnancy: The Generation R Study. Eur. J. Epidemiol. 2015, 30, 1057–1066. [Google Scholar] [CrossRef] [PubMed]
- Velegrakis, A.; Sfakiotaki, M.; Sifakis, S. Human placental growth hormone in normal and abnormal fetal growth. Biomed. Rep. 2017, 7, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Acar, A.; Altinbas, A.; Ozturk, M.; Kosar, A.; Kirazli, S. Selectins in normal pregnancy, pre-eclampsia and missed abortus. Haematologia 2001, 31, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Andres, F.; Wong, G.P.; Walker, S.P.; MacDonald, T.M.; Keenan, E.; Cannon, P.; Nguyen, T.V.; Hannan, N.J.; Tong, S.; Kaitu’u-Lino, T.J. A disintegrin and metalloproteinase 12 (ADAM12) is reduced at 36 weeks’ gestation in pregnancies destined to deliver small for gestational age infants. Placenta 2022, 117, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Arevalo, E.; Shanmugasundararaj, S.; Wilkemeyer, M.F.; Dou, X.; Chen, S.; Charness, M.E.; Miller, K.W. An alcohol binding site on the neural cell adhesion molecule L1. Proc. Natl. Acad. Sci. USA 2008, 105, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Bearer, C.F. Mechanisms of brain injury: L1 cell adhesion molecule as a target for ethanol-induced prenatal brain injury. Semin. Pediatr. Neurol. 2001, 8, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Erenel, H.; Yilmaz, N.; Cift, T.; Bulut, B.; Sozen, I.; Aslan Cetin, B.; Gezer, A.; Ekmekci, H.; Kaya, B.; Tuten, A. Maternal serum autotaxin levels in early- and late-onset preeclampsia. Hypertens. Pregnancy 2017, 36, 310–314. [Google Scholar] [CrossRef] [PubMed]
- Kremer, A.E.; Bolier, R.; Dixon, P.H.; Geenes, V.; Chambers, J.; Tolenaars, D.; Ris-Stalpers, C.; Kaess, B.M.; Rust, C.; van der Post, J.A.; et al. Autotaxin activity has a high accuracy to diagnose intrahepatic cholestasis of pregnancy. J. Hepatol. 2015, 62, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Laigaard, J.; Spencer, K.; Christiansen, M.; Cowans, N.J.; Larsen, S.O.; Pedersen, B.N.; Wewer, U.M. ADAM 12 as a first-trimester maternal serum marker in screening for Down syndrome. Prenat. Diagn. 2006, 26, 973–979. [Google Scholar] [CrossRef]
- Li, Y.T.; Chen, J.S.; Jian, W.; He, Y.D.; Li, N.; Xie, Y.N.; Wang, J.; Zhang, V.W.; Huang, W.R.; Jiang, F.M.; et al. L1CAM mutations in three fetuses diagnosed by medical exome sequencing. Taiwan. J. Obstet. Gynecol. 2020, 59, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Tuzluoglu, S.; Ustunyurt, E.; Karasin, S.S.; Karasin, Z.T. Investigation of Serum Pregnancy-Specific Beta-1-Glycoprotein and Relationship with Fetal Growth Restriction. JBRA Assist. Reprod. 2022, 26, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Ramanjaneya, M.; Butler, A.E.; Bashir, M.; Bettahi, I.; Moin, A.S.M.; Ahmed, L.; Elrayess, M.A.; Hunt, S.C.; Atkin, S.L.; Abou-Samra, A.B. apoA2 correlates to gestational age with decreased apolipoproteins A2, C1, C3 and E in gestational diabetes. BMJ Open Diabetes Res. Care 2021, 9, e001925. [Google Scholar] [CrossRef] [PubMed]
- Dou, X.; Menkari, C.; Mitsuyama, R.; Foroud, T.; Wetherill, L.; Hammond, P.; Suttie, M.; Chen, X.; Chen, S.Y.; Charness, M.E.; et al. L1 coupling to ankyrin and the spectrin-actin cytoskeleton modulates ethanol inhibition of L1 adhesion and ethanol teratogenesis. FASEB J. 2018, 32, 1364–1374. [Google Scholar] [CrossRef]
- The Human Protein Atlas. Available online: https://www.proteinatlas.org/ (accessed on 10 December 2024).
- Kasimis, C.; Evangelinakis, N.; Rotas, M.; Georgitsi, M.; Pelekanos, N.; Kassanos, D. Predictive value of biochemical marker ADAM-12 at first trimester of pregnancy for hypertension and intrauterine growth restriction. Clin. Exp. Obstet. Gynecol. 2016, 43, 43–47. [Google Scholar] [CrossRef]
- Toprak, K.; Yildiz, Z.; Akdemir, S.; Esen, K.; Kada, R.; Can Gulec, N.; Omar, B.; Bicer, A.; Demirbag, R. Low pregnancy-specific beta-1-glycoprotein is associated with nondipper hypertension and increased risk of preeclampsia in pregnant women with newly diagnosed chronic hypertension. Scand. J. Clin. Lab. Investig. 2023, 83, 479–488. [Google Scholar] [CrossRef]
- Ilicic, M.; Zakar, T.; Paul, J.W. The Regulation of Uterine Function During Parturition: An Update and Recent Advances. Reprod. Sci. 2020, 27, 3–28. [Google Scholar] [CrossRef] [PubMed]
- Bonney, E.A. Demystifying animal models of adverse pregnancy outcomes: Touching bench and bedside. Am. J. Reprod. Immunol. 2013, 69, 567–584. [Google Scholar] [CrossRef]
- Wegienka, G.; Baird, D.D. A comparison of recalled date of last menstrual period with prospectively recorded dates. J. Womens Health 2005, 14, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Barr, W.B.; Pecci, C.C. Last menstrual period versus ultrasound for pregnancy dating. Int. J. Gynaecol. Obstet. 2004, 87, 38–39. [Google Scholar] [CrossRef] [PubMed]
- Christians, J.K.; Beristain, A.G. ADAM12 and PAPP-A: Candidate regulators of trophoblast invasion and first trimester markers of healthy trophoblasts. Cell Adhes. Migr. 2016, 10, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.E.; Thomas, G.; Tuytten, R.; Van Herrewege, Y.; Djiokep, R.O.; Roberts, C.T.; Kenny, L.C.; Simpson, N.A.; North, R.A.; Baker, P.N. Mid-trimester maternal ADAM12 levels differ according to fetal gender in pregnancies complicated by preeclampsia. Reprod. Sci. 2015, 22, 235–241. [Google Scholar] [CrossRef]
- Gaccioli, F.; Aye, I.; Sovio, U.; Charnock-Jones, D.S.; Smith, G.C.S. Screening for fetal growth restriction using fetal biometry combined with maternal biomarkers. Am. J. Obstet. Gynecol. 2018, 218, S725–S737. [Google Scholar] [CrossRef]
- Montagnana, M.; Trenti, T.; Aloe, R.; Cervellin, G.; Lippi, G. Human chorionic gonadotropin in pregnancy diagnostics. Clin. Chim. Acta 2011, 412, 1515–1520. [Google Scholar] [CrossRef] [PubMed]
- Blois, S.M.; Sulkowski, G.; Tirado-González, I.; Warren, J.; Freitag, N.; Klapp, B.F.; Rifkin, D.; Fuss, I.; Strober, W.; Dveksler, G.S. Pregnancy-specific glycoprotein 1 (PSG1) activates TGF-beta and prevents dextran sodium sulfate (DSS)-induced colitis in mice. Mucosal Immunol. 2014, 7, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Shanley, D.K.; Kiely, P.A.; Golla, K.; Allen, S.; Martin, K.; O’Riordan, R.; Ball, M.; Aplin, J.D.; Singer, B.B.; Caplice, N.; et al. Pregnancy-specific glycoproteins bind integrin alphaIIbbeta3 and inhibit the platelet-fibrinogen interaction. PLoS ONE 2013, 8, e57491. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.F.; Cervi, L.; Knubel, C.P.; Panzetta-Dutari, G.M.; Motran, C.C. The role of pregnancy-specific glycoprotein 1a (PSG1a) in regulating the innate and adaptive immune response. Am. J. Reprod. Immunol. 2013, 69, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.F.; Knubel, C.P.; Sanchez, M.C.; Cervi, L.; Motran, C.C. Pregnancy-specific glycoprotein 1a activates dendritic cells to provide signals for Th17-, Th2-, and Treg-cell polarization. Eur. J. Immunol. 2012, 42, 1573–1584. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.Q.; Baranov, V.; Zimmermann, W.; Grunert, F.; Erhard, B.; Mincheva-Nilsson, L.; Hammarstrom, S.; Thompson, J. Highly specific monoclonal antibody demonstrates that pregnancy-specific glycoprotein (PSG) is limited to syncytiotrophoblast in human early and term placenta. Placenta 1997, 18, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Sargent, I.L.; Borzychowski, A.M.; Redman, C.W. NK cells and human pregnancy—An inflammatory view. Trends Immunol. 2006, 27, 399–404. [Google Scholar] [CrossRef]
- Mahajan, D.; Sharma, N.R.; Kancharla, S.; Kolli, P.; Tripathy, A.; Sharma, A.K.; Singh, S.; Kumar, S.; Mohanty, A.K.; Jena, M.K. Role of Natural Killer Cells during Pregnancy and Related Complications. Biomolecules 2022, 12, 68. [Google Scholar] [CrossRef]
- Okamoto, N.; Uchida, A.; Takakura, K.; Kariya, Y.; Kanzaki, H.; Riittinen, L.; Koistinen, R.; Seppala, M.; Mori, T. Suppression by human placental protein 14 of natural killer cell activity. Am. J. Reprod. Immunol. 1991, 26, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, D.; Sundereshan, S.; Rao, C.; Karande, A.A. Placental protein 14 induces apoptosis in T cells but not in monocytes. J. Biol. Chem. 2001, 276, 28268–28273. [Google Scholar] [CrossRef]
- Licheri, V.; Brigman, J.L. Altering Cell-Cell Interaction in Prenatal Alcohol Exposure Models: Insight on Cell-Adhesion Molecules During Brain Development. Front. Mol. Neurosci. 2021, 14, 753537. [Google Scholar] [CrossRef]
- Cau, F.; Fanni, D.; Manchia, M.; Gerosa, C.; Piras, M.; Murru, R.; Paribello, P.; Congiu, T.; Coni, P.; Pichiri, G.; et al. Expression of L1 Cell Adhesion Molecule (L1CAM) in extracellular vesicles in the human spinal cord during development. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 6273–6282. [Google Scholar] [CrossRef] [PubMed]
- Prakobphol, A.; Genbacev, O.; Gormley, M.; Kapidzic, M.; Fisher, S.J. A role for the L-selectin adhesion system in mediating cytotrophoblast emigration from the placenta. Dev. Biol. 2006, 298, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Ma, X.; Deng, L.; Yao, B.; Xiong, Y.; Wu, Y.; Wang, L.; Ma, Q.; Ma, F. Role of selectins and their ligands in human implantation stage. Glycobiology 2017, 27, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Genbacev, O.D.; Prakobphol, A.; Foulk, R.A.; Krtolica, A.R.; Ilic, D.; Singer, M.S.; Yang, Z.Q.; Kiessling, L.L.; Rosen, S.D.; Fisher, S.J. Trophoblast L-selectin-mediated adhesion at the maternal-fetal interface. Science 2003, 299, 405–408. [Google Scholar] [CrossRef]
- Gelius, E.; Persson, C.; Karlsson, J.; Steiner, H. A mammalian peptidoglycan recognition protein with N-acetylmuramoyl-L-alanine amidase activity. Biochem. Biophys. Res. Commun. 2003, 306, 988–994. [Google Scholar] [CrossRef]
- Lo, D.; Tynan, W.; Dickerson, J.; Mendy, J.; Chang, H.W.; Scharf, M.; Byrne, D.; Brayden, D.; Higgins, L.; Evans, C.; et al. Peptidoglycan recognition protein expression in mouse Peyer’s Patch follicle associated epithelium suggests functional specialization. Cell. Immunol. 2003, 224, 8–16. [Google Scholar] [CrossRef]
- Tosoni, G.; Conti, M.; Diaz Heijtz, R. Bacterial peptidoglycans as novel signaling molecules from microbiota to brain. Curr. Opin. Pharmacol. 2019, 48, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Arentsen, T.; Qian, Y.; Gkotzis, S.; Femenia, T.; Wang, T.; Udekwu, K.; Forssberg, H.; Diaz Heijtz, R. The bacterial peptidoglycan-sensing molecule Pglyrp2 modulates brain development and behavior. Mol. Psychiatry 2017, 22, 257–266. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Inclusion Criteria | Exclusion Criteria |
|---|---|
|
|
| Demographic/Clinical Variable | Value | All Subjects (n = 2648) |
|---|---|---|
| Maternal Age | Median (IQR) | 28 years (23–32) |
| Maternal Pre-pregnancy BMI | Median (IQR) | 25.90 (22.3–31.4) * |
| Gravida | Primigravida | 710 (26.8 %) |
| Multigravida | 1938 (73.2 %) | |
| Race | Black | 414 (15.6%) |
| White | 1998 (75.5%) | |
| Other | 236 (8.9%) | |
| Ethnicity | Hispanic | 942 (35.6%) |
| Not Hispanic | 1706 (64.4%) | |
| Gestational age at blood draw (GABD) | Median (IQR) | 166 days (141–184) |
| Gestational age at birth (GAB) | Median (IQR) | 274 days (270–280) |
| Diabetes ** | Gestational | 183 (6.9%) |
| Type I | 19 (0.7%) | |
| Type II | 55 (2.1%) | |
| None | 2391 (90.3%) | |
| Pre-eclampsia | No | 2516 (95.0%) |
| Yes | 132 (5.0%) | |
| Pregnancy-Induced Hypertension | No | 2580 (97.4%) |
| Yes | 68 (2.6%) | |
| Other Complications *** | No | 2455 (92.7%) |
| Yes | 193 (7.3%) | |
| Delivery | Cesarean Section: Primary | 405 (15.3%) |
| Cesarean Section: Repeat | 404 (15.3%) | |
| Vaginal | 1839 (69.4%) |
| Protein (UniProt ID) | Expression Change Between 18 and 20 Weeks of Gestation and 26 and 28 Weeks of Gestation, Wilcoxon Test | Placentally Expressed [46] | Protein Type | Role in Pregnancy | Previously Reported Associations with TTB or Pregnancy Complications |
|---|---|---|---|---|---|
| Peptidoglycan Recognition Protein 2 (PGRP2) | Downregulated NS | Yes | Hydrolase enzyme | Pattern recognition protein | None |
| Cell Adhesion Molecule L1 Like (CHL1) | Downregulated NS | No | Cell adhesion molecule | Negative regulator of proliferation | Brain malformation and neurodevelopmental delay [37,38,42] |
| Lymphocyte Adhesion Molecule 1 (LYAM1) | Downregulated NS | No | Cell adhesion molecule | Cell adhesion and migration | Preeclampsia [35] |
| Dipeptidase 2 (DPEP2) | Upregulated | Yes | Hydrolase enzyme | Cell differentiation | None |
| Growth Hormone 2 (SOM2) | Upregulated | Yes | Growth hormone | Cell differentiation and proliferation | Marker of gestational age [31] |
| Chorionic somatomammotropin (CSH) * | Upregulated | Yes | Growth hormone | Stimulates lactation and fetal growth | Marker of gestational age [16] |
| Progestogen-associated endometrial protein (PAEP) | Downregulated | Yes | Glycoprotein | Regulates uterine environment/immune cell inhibitor | None |
| Chorionic Gonadotropin Subunit Beta 1 (CGB1) | Downregulated | Yes | ꞵ-subunit of human chorionic gonadotropin | Stimulates uterine and fetal growth/immune modulator | Marker of gestational age [33] |
| Pregnancy-Specific ꞵ1 Glycoprotein 11 (PSG11) | Upregulated | Yes | Placental glycoprotein | Immune cell and angiogenesis modulator | None |
| Disintegrin and metalloproteinase domain-containing protein 12 (ADA12) | Upregulated | Yes | Placental glycoprotein | Placental growth and differentiation | Preterm birth, fetal growth restriction, preeclampsia, Down Syndrome, small for gestational age [36,41,47] |
| Pregnancy-Specific ꞵ1 Glycoprotein 2 (PSG2) | Upregulated | Yes | Placental glycoprotein | Immune cell and angiogenesis modulator | None |
| Sushi, von Willebrand Factor type A, EGF, and pentraxin domain-containing protein (SVEP1) | Upregulated | Yes | Cell adhesion molecule | Facilitates cell alignment and migration | Marker of gestational age [32] |
| Pregnancy-Specific ꞵ1 Glycoprotein 1 (PSG1) | Upregulated NS | Yes | Placental glycoprotein | Immune cell and angiogenesis modulator | Preeclampsia [48] |
| Ectonucleotide Pyrophosphatase/Phosphodiesterase 2 (ENPP2) | Upregulated | Yes | Phosphodiesterase and phospholipase | Cell proliferation and migration | Preeclampsia [39,40] |
| Apolipoprotein C-III (APOC3) | Upregulated | No | Lipid binding protein | Modulator of lipid metabolism | Gestational Diabetes Mellitus [44] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dufford, M.T.; Fleischer, T.C.; Sommerville, L.J.; Badsha, M.B.; Polpitiya, A.D.; Logan, J.; Fox, A.C.; Rust, S.R.; Cox, C.B.; Garite, T.J.; et al. Clock Proteins Have the Potential to Improve Term Delivery Date Prediction: A Proof-of-Concept Study. Life 2025, 15, 224. https://doi.org/10.3390/life15020224
Dufford MT, Fleischer TC, Sommerville LJ, Badsha MB, Polpitiya AD, Logan J, Fox AC, Rust SR, Cox CB, Garite TJ, et al. Clock Proteins Have the Potential to Improve Term Delivery Date Prediction: A Proof-of-Concept Study. Life. 2025; 15(2):224. https://doi.org/10.3390/life15020224
Chicago/Turabian StyleDufford, Max T., Tracey C. Fleischer, Laura J. Sommerville, Md. Bahadur Badsha, Ashoka D. Polpitiya, Jennifer Logan, Angela C. Fox, Sharon R. Rust, Charles B. Cox, Thomas J. Garite, and et al. 2025. "Clock Proteins Have the Potential to Improve Term Delivery Date Prediction: A Proof-of-Concept Study" Life 15, no. 2: 224. https://doi.org/10.3390/life15020224
APA StyleDufford, M. T., Fleischer, T. C., Sommerville, L. J., Badsha, M. B., Polpitiya, A. D., Logan, J., Fox, A. C., Rust, S. R., Cox, C. B., Garite, T. J., Boniface, J. J., & Kearney, P. E. (2025). Clock Proteins Have the Potential to Improve Term Delivery Date Prediction: A Proof-of-Concept Study. Life, 15(2), 224. https://doi.org/10.3390/life15020224