
Morphological Evolution and Extinction of Eodiscids and Agnostoid Arthropods


Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Geometric Morphometric Analyses
3.2. Morphospace Visualization and Morphological Disparity
4. Discussion
4.1. Response to the Cambrian Geological Events
4.2. Response to the GOBE
4.3. Extinction Mechanism
5. Conclusions
- (1)
- Our data indicate that anoxia from the BTE, ROECE, and SPICE events [16,20,77] may be the main reason for the periodic decline of the morphospace occupation of eodiscids and agnostoid arthropods, as anoxic events seemingly eliminated a large number of morphological types and reduced the overall morphospace of the clade.
- (2)
- During the Ordovician, abiotic factors such as temperature, as well as biotic factors such as competition, may have led to a reduction in morphospace occupation for the agnostinids. There was no observable increase in agnostinids morphological disparity during the GOBE, unlike for many groups.
- (3)
- After the Cambrian Series 2, the repeated and periodic decline in morphological disparity of eodiscids and agnostoid arthropods is consistent with multiple geological events during the Cambrian–Ordovician. The extinction of eodiscids and agnostoid arthropods is likely the result of a stepwise decrease in the total morphological occupancy, rather than a single, sudden event, and accordingly cannot be tied to a single cause, either abiotic or biotic.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Whittington, H.B.; Chatterton, B.D.E.; Speyer, S.E.; Fortey, R.A.; Owens, R.M.; Chang, W.T.; Dean, W.T.; Jell, P.A.; Laurie, J.R.; Palmer, A.R.; et al. Trilobita: Introduction, Order Agnostida, Order Redlichiida. In Treatise on Invertebrate Paleontology: Part O. Arthropoda 1 (Revised); Kaesler, R.L., Ed.; Geological Society of America & University of Kansas: Lawrence, KS, USA, 1997; pp. 1–530. [Google Scholar]
- Peng, S.C.; Robision, R.A. Agnostoid biostratigraphy across the Middle–Upper Cambrian boundary in Hunan, China. J. Paleontol. 2000, 74, 1–104. [Google Scholar] [CrossRef]
- Ogg, J.G.; Ogg, G.M.; Gradstein, F.M. A Concise Geologic Time Scale: 2016; Elsevier: Amsterdam, The Netherlands, 2016; pp. 41–55. [Google Scholar] [CrossRef]
- Babcock, L.E.; Peng, S.C.; Ahlberg, P. Cambrian trilobite biostratigraphy and its role in developing an integrated history of the Earth system. Lethaia 2017, 50, 381–399. [Google Scholar] [CrossRef]
- Dai, T.; Zhang, X.L. Morphology and ontogeny of the eodiscoid trilobite Sinodiscus changyangensis from the lower Cambrian of South China. Palaeontology 2013, 56, 411–420. [Google Scholar] [CrossRef]
- Müller, K.J.; Waloszek, D. Morphology, ontogeny, and life habit of Agnostus pisiformis from the Upper Cambrian of Sweden. In Fossils and Strata; Bengtson, S., Ed.; Universitetsforlaget: Oslo, Norway, 1987; pp. 1–124. [Google Scholar]
- Adrain, J.M. Class Trilobita Walch, 1771. In Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness; Zhang, Z.Q., Ed.; Zootaxa: Auckland, NZ, 2011; pp. 104–109. [Google Scholar]
- Ramsköld, L.; Edgecombe, G.D. Trilobite monophyly revisited. Hist. Biol. 1990, 4, 267–283. [Google Scholar] [CrossRef]
- Vannier, J.; Aria, C.; Taylor, R.S.; Caron, J.-B. Waptia fieldensis Walcott, a mandibulate arthropod from the middle Cambrian Burgess Shale. R. Soc. Open Sci. 2018, 5, 172206. [Google Scholar] [CrossRef]
- Bergström, J.; Hou, X.G. Early Palaeozoic non-lamellipedian arthropods. In Crustacea and Arthropod Relationships; Koenemann, S., Jenner, R.A., Eds.; Taylor & Francis: Boca Raton, FL, USA, 2005; pp. 73–93. [Google Scholar]
- Moysiuk, J.; Caron, J.B. Burgess Shale fossils shed light on the agnostid problem. Proc. R. Soc. B 2019, 286, 20182314. [Google Scholar] [CrossRef]
- Jell, P.A. Phylogeny of early Cambrian trilobites. Spec. Pap. Palaeontol. 2003, 70, 45–57. [Google Scholar]
- Cotton, T.J.; Fortey, R.A. Comparative morphology and relationships of the Agnostida. In Crustacea and Arthropod Relationships; Koenemann, S., Jenner, R.A., Eds.; Taylor & Francis: Boca Raton, FL, USA, 2005; pp. 95–123. [Google Scholar]
- Peng, S.C. Illustrations of Cambrian Stratigraphy and Index Fossils of China, Trilobites; Zhejiang University Press: Hangzhou, China, 2019; pp. 54–151. [Google Scholar]
- Signor, P.W. Taxonomic diversity and faunal turnover in the Early Cambrian: Did the most severe mass extinction of the Phanerozoic occur in the Botomian stage? Paleontol. Soc. Spec. Publ. 1992, 6, 272. [Google Scholar] [CrossRef]
- Zhuravlev, A.Y.; Wood, R.A. Anoxia as the cause of the mid-Early Cambrian (Botomian) extinction event. Geology 1996, 24, 311–314. [Google Scholar] [CrossRef]
- Zhuravlev, A.Y.; Wood, R.A. The two phases of the Cambrian Explosion. Sci. Rep. 2018, 8, 16656. [Google Scholar] [CrossRef]
- He, T.C.; Zhu, M.Y.; Mills, B.J.W.; Wynn, P.M.; Zhuravlev, A.Y.; Tostevin, R.; von Strandmann, P.A.E.P.; Yang, A.; Poulton, S.W.; Shields, G.A. Possible links between extreme oxygen perturbations and the Cambrian radiation of animals. Nat. Geosci. 2019, 12, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Hu, W.X.; Wang, X.L.; Yu, H.; Yang, A.H.; Cao, J.; Yao, S.P. Carbon isotope stratigraphy of the lower to middle Cambrian on the eastern Yangtze Platform, South China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 479, 90–101. [Google Scholar] [CrossRef]
- Zhu, M.Y.; Babcock, L.E.; Peng, S.C. Advances in Cambrian stratigraphy and paleontology: Integrating correlation techniques, paleobiology, taphonomy and paleoenvironmental reconstruction. Palaeoworld 2006, 15, 217–222. [Google Scholar] [CrossRef]
- Saltzman, M.R.; Ripperdan, R.L.; Brasier, M.D.; Lohmann, K.C.; Robison, R.A.; Chang, W.T.; Peng, S.C.; Ergaliev, E.K.; Runnegar, B. A global carbon isotope excursion (SPICE) during the Late Cambrian: Relation to trilobite extinctions, organic-matter burial and sea level. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 162, 211–223. [Google Scholar] [CrossRef]
- Wood, R.; Liu, A.G.; Bowyer, F.; Wilby, P.R.; Dunn, F.S.; Kenchington, C.G.; Cuthill, J.F.H.; Mitchell, E.G.; Penny, A. Integrated records of environmental change and evolution challenge the Cambrian Explosion. Nat. Ecol. Evol. 2019, 3, 528–538. [Google Scholar] [CrossRef]
- Bian, L.; Chappaz, A.; Schovsbo, N.H.; Wang, X.M.; Zhao, W.Z.; Sanei, H. A 20-million-year reconstruction to decipher the enigmatic Cambrian extinction–Ordovician biodiversification transition. Earth Planet. Sci. Lett. 2023, 612, 118170. [Google Scholar] [CrossRef]
- Servais, T.; Harper, D.A.T. The Great Ordovician Biodiversification Event (GOBE): Definition, concept and duration. Lethaia 2018, 51, 151–164. [Google Scholar] [CrossRef]
- Servais, T.; Cascales-Miñana, B.; Harper, D.A.T. The Great Ordovician Biodiversification Event (GOBE) is not a single event. Paleontol. Res. 2021, 25, 315–328. [Google Scholar] [CrossRef]
- Edwards, C.T.; Saltzman, M.R.; Royer, D.L. Oxygenation as a driver of the Great Ordovician Biodiversication Event. Nat. Geosci. 2016, 10, 925–929. [Google Scholar] [CrossRef]
- Gon, S.M., III. A Pictorial guide to the Orders of Trilobites. 1999. Available online: http://www.trilobites.info (accessed on 9 December 2022).
- Zhang, S.H.; Fan, J.X.; Morgan, C.A.; Henderson, C.M.; Shen, S.Z. Quantifying the middle–late Cambrian trilobite diversity pattern in South China. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2021, 570, 110361. [Google Scholar] [CrossRef]
- Jackson, I.S.C.; Budd, G.E. Intraspecific morphological variation of Agnostus pisiformis, a Cambrian Series 3 trilobite-like arthropod. Lethaia 2017, 50, 467–485. [Google Scholar] [CrossRef]
- Suárez, M.G.; Esteve, J. Morphological diversity and disparity in trilobite cephala and the evolution of trilobite enrolment throughout the Palaeozoic. Lethaia 2021, 54, 752–761. [Google Scholar] [CrossRef]
- Fischer, A.G. A carapace of the Ordovician trilobite Telephus. J. Paleontol. 1946, 20, 566–569. [Google Scholar]
- Glaessner, M.F. An Ordovician Trilobite and some later Paleozoic Crustacea in moulting positions. J. Paleontol. 1948, 22, 531. [Google Scholar]
- Busch, R.M.; Swartz, F.M. Molting and description of a new homalonotid trilobite from Pennsylvania. J. Paleontol. 1985, 59, 1062–1074. [Google Scholar]
- Brandt, D.S. Ecdysis in Flexicalymene meeki (Trilobita). J. Paleontol. 1993, 67, 999–1005. [Google Scholar] [CrossRef]
- McNamara, K.J. Techniques of exuviation in Australian species of the Cambrian trilobite Redlichia. Alcheringa 1986, 10, 403–412. [Google Scholar] [CrossRef]
- Wang, Y.F.; Peng, J.; Wang, Q.J.; Wen, R.Q.; Zhang, H.; Du, G.Y.; Shao, Y.B. Moulting in the Cambrian oryctocephalid trilobite Arthricocephalites xinzhaiheensis from Guizhou Province, South China. Lethaia 2021, 54, 211–228. [Google Scholar] [CrossRef]
- Drage, H.B.; Daley, A.C. Recognising moulting behaviour in trilobites by examining morphology, development, and preservation: Comment on Błażejowski et al. 2015. BioEssays 2016, 38, 981–990. [Google Scholar] [CrossRef]
- Drage, H.B.; Holmes, J.D.; Garcia-Bellido, D.C.; Daley, A.C. An exceptional record of Cambrian trilobite moulting behaviour preserved in the Emu Bay Shale, South Australia. Lethaia 2018, 51, 473–492. [Google Scholar] [CrossRef]
- Drage, H.B. Quantifying intra-and interspecific variability in trilobite moulting behaviour across the Palaeozoic. Palaeontol. Electron. 2019, 22.2.34A, 1–39. [Google Scholar] [CrossRef] [PubMed]
- Drage, H.B.; Legg, D.A.; Daley, A.C. Novel marrellomorph moulting behaviour preserved in the Lower Ordovician Fezouata Shale, Morocco. Front. Ecol. Evol. 2023, 11, 1226924. [Google Scholar] [CrossRef]
- Drage, H.B.; Holmes, J.D.; García-Bellido, D.C.; Paterson, J.R. Associations between trilobite intraspecific moulting variability and body proportions: Estaingia bilobata from the Cambrian Emu Bay Shale, Australia. Palaeontology 2023, 66, e12651. [Google Scholar] [CrossRef]
- Drage, H.B. Trilobite moulting behaviour variability had little association with morphometry. bioRxiv 2022. [Google Scholar] [CrossRef]
- Clarkson, E.; Levi-Setti, R.; Horváth, G. The eyes of trilobites: The oldest preserved visual system. Arthropod Struct. Dev. 2006, 35, 247–259. [Google Scholar] [CrossRef]
- Scholtz, G.; Staude, A.; Dunlop, J.A. Trilobite compound eyes with crystalline cones and rhabdoms show mandibulate affinities. Nat. Commun. 2019, 10, 2503. [Google Scholar] [CrossRef]
- Schoenemann, B. An overview on trilobite eyes and their functioning. Arthropod Struct. Dev. 2021, 61, 101032. [Google Scholar] [CrossRef]
- Vargas-Parra, E.E.; Hopkins, M.J. Modularity in the trilobite head consistent with the hypothesized segmental origin of the eyes. Evol. Dev. 2022, 24, 177–188. [Google Scholar] [CrossRef]
- Fortey, R.A.; Owens, R.M. Feeding habits in trilobites. Palaeontology 1999, 42, 429–465. [Google Scholar] [CrossRef]
- Hopkins, M.J.; Chen, F.Y.; Hu, S.X.; Zhang, Z.F. The oldest known digestive system consisting of both paired digestive glands and a crop from exceptionally preserved trilobites of the Guanshan Biota (Early Cambrian, China). PLoS ONE 2017, 12, e0184982. [Google Scholar] [CrossRef]
- Park, T.Y.S.; Kihm, J.H. Head segmentation of trilobites. Lethaia 2017, 50, 1–6. [Google Scholar] [CrossRef]
- Foote, M. Morphologic patterns of diversification: Examples from trilobites. Palaeontology 1991, 34, 461–485. [Google Scholar]
- Brusatte, S.L.; Butler, R.J.; Prieto-Márquez, A.; Norell, M.A. Dinosaur morphological diversity and the end-Cretaceous extinction. Nat. Commun. 2012, 3, 804. [Google Scholar] [CrossRef] [PubMed]
- Bault, V.; Crônier, C.; Monnet, C. Coupling of taxonomic diversity and morphological disparity in Devonian trilobites? Hist. Biol. 2023, 36, 473–484. [Google Scholar] [CrossRef]
- Naimark, E.B.; Pegel, T.V. Revision of the Cambrian Agnostina (Trilobita?) from Russia. Paleontol. J. 2017, 51, 1167–1248. [Google Scholar] [CrossRef]
- Cui, L.H.; Dai, T.; Zhang, X.L.; Peng, S.C. Morphology and ontogeny of the eodiscoid trilobite Pagetia vinusta from the Cambrian (Miaolingian) of South China. PalZ 2019, 93, 195–206. [Google Scholar] [CrossRef]
- Dai, T.; Zhang, X.L. Ontogeny of the eodiscoid trilobite Tsunyidiscus acutus from the lower Cambrian of South China. Palaeontology 2011, 54, 1279–1288. [Google Scholar] [CrossRef]
- Bault, V.; Crônier, C.; Monnet, C. Morphological disparity trends in Devonian trilobites from North Africa. Palaeontology 2022, 65, e12623. [Google Scholar] [CrossRef]
- Bault, V.; Crônier, C.; Monnet, C.; Balseiro, D.; Serra, F.; Waisfeld, B.; Bignon, A.; Rustán, J.J. Rise and fall of the phacopids: The morphological history of a successful trilobite family. Palaeontology 2023, 66, e12673. [Google Scholar] [CrossRef]
- Rohlf, F.J. The tps series of software. Hystrix 2015, 26, 9–12. [Google Scholar] [CrossRef]
- Rohlf, F.J. TpsUtil, File Utility Program, version 1.83; Department of Ecology & Evolution, State University of New York at Stony Brook: New York, NY, USA, 2011. [Google Scholar]
- Rohlf, F.J. TpsRelw, Relative Warps Analysis, version 1.75; Department of Ecology & Evolution, State University of New York at Stony Brook: New York, NY, USA, 2010. [Google Scholar]
- Abdi, H.; Williams, L.J. Principal component analysis. WIRES Comput. Stat. 2010, 2, 433–459. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- McKinney, W. Python for Data Analysis, 3rd ed.; O’Reilly Media, Inc.: Sebastopol, CA, USA, 2022. [Google Scholar]
- Ciampaglio, C.N.; Kemp, M.; McShea, D.W. Detecting changes in morphospace occupation patterns in the fossil record: Characterization and analysis of measures of disparity. Paleobiology 2001, 27, 695–715. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Guillerme, T. dispRity: A modular R package for measuring disparity. Methods Ecol. Evol. 2018, 9, 1755–1763. [Google Scholar] [CrossRef]
- Collyer, M.L.; Sekora, D.J.; Adams, D.C. A method for analysis of phenotypic change for phenotypes described by high-dimensional data. Heredity 2015, 115, 357–365. [Google Scholar] [CrossRef]
- Foote, M. Morphological disparity in Ordovician–Devonian crinoids and the early saturation of morphological space. Paleobiology 1994, 20, 320–344. [Google Scholar] [CrossRef]
- Hughes, M.; Gerber, S.; Wills, M.A. Clades reach highest morphological disparity early in their evolution. Proc. Natl. Acad. Sci. USA 2013, 110, 13875–13879. [Google Scholar] [CrossRef]
- Cooney, C.R.; Bright, J.A.; Capp, E.J.; Chira, A.M.; Hughes, E.C.; Christopher, J.A.; Moody, C.J.; Nouri, L.O.; Varley, Z.K.; Thomas, G.H. Mega-evolutionary dynamics of the adaptive radiation of birds. Nature 2017, 542, 344–347. [Google Scholar] [CrossRef]
- Budd, G.E. Morphospace. Curr. Biol. 2021, 31, R1181–R1185. [Google Scholar] [CrossRef]
- Erwin, D.H. Early introduction of major morphological innovations. Acta Palaeontol. Pol. 1994, 38, 281–294. [Google Scholar]
- Erwin, D.H. Disparity: Morphological pattern and developmental context. Palaeontology 2007, 50, 57–73. [Google Scholar] [CrossRef]
- Webster, M. A Cambrian peak in morphological variation within trilobite species. Science 2007, 317, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Oyston, J.W.; Hughes, M.; Wagner, P.J.; Gerber, S.; Wills, M.A. What limits the morphological disparity of clades? Interface Focus 2015, 5, 20150042. [Google Scholar] [CrossRef]
- Zhu, M.Y.; Yang, A.H.; Yuan, J.L.; Li, G.X.; Zhang, J.M.; Zhao, F.C.; Ahn, S.Y.; Miao, L.Y. Cambrian integrative stratigraphy and timescale of China. Sci. China Earth Sci. 2019, 62, 25–60. [Google Scholar] [CrossRef]
- Gill, B.C.; Lyons, T.W.; Young, S.A.; Kump, L.R.; Knoll, A.H.; Saltzman, M.R. Geochemical evidence for widespread euxinia in the Later Cambrian ocean. Nature 2011, 469, 80–83. [Google Scholar] [CrossRef]
- Saltzman, M.R.; Young, S.A.; Kump, L.R.; Gill, B.C.; Lyons, T.W.; Runnegar, B. Pulse of atmospheric oxygen during the late Cambrian. Proc. Natl. Acad. Sci. USA 2011, 108, 3876–3881. [Google Scholar] [CrossRef]
- Servais, T.; Perrier, V.; Danelian, T.; Klug, C.; Martin, R.; Munnecke, A.; Nowak, H.; Nützel, A.; Vandenbroucke, T.R.A.; Williams, M.; et al. The onset of the ‘Ordovician Plankton Revolution’ in the late Cambrian. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 458, 12–28. [Google Scholar] [CrossRef]
- Ontiveros, D.E.; Beaugrand, G.; Lefebvre, B.; Marcilly, C.M.; Servais, T.; Pohl, A. Impact of global climate cooling on Ordovician marine biodiversity. Nat. Commun. 2023, 14, 6098. [Google Scholar] [CrossRef]
- Trotter, J.A.; Williams, I.S.; Barnes, C.R.; Lécuyer, C.; Nicoll, R.S. Did cooling oceans trigger Ordovician biodiversification? Evidence from conodont thermometry. Science 2008, 321, 550–554. [Google Scholar] [CrossRef]
- Fan, J.X.; Shen, S.Z.; Erwin, D.H.; Sadler, P.M.; MacLeod, N.; Cheng, Q.M.; Hou, X.D.; Yang, J.; Wang, X.D.; Wang, Y.; et al. A high-resolution summary of Cambrian to Early Triassic marine invertebrate biodiversity. Science 2020, 367, 272–277. [Google Scholar] [CrossRef]
- Fortey, R. The palaeoecology of trilobites. J. Zool 2014, 292, 250–259. [Google Scholar] [CrossRef]
- Adrain, J.M.; Fortey, R.A.; Westrop, S.R. Post-Cambrian trilobite diversity and evolutionary faunas. Science 1998, 280, 1922–1925. [Google Scholar] [CrossRef] [PubMed]
- Westrop, S.R.; Adrain, J.M. Trilobite alpha diversity and the reorganization of Ordovician benthic marine communities. Paleobiology 1998, 24, 1–16. [Google Scholar] [CrossRef]
- Rong, J.Y.; Huang, B. Study of Mass Extinction over the past thirty years: A synopsis. Sci. Sin. Terrae 2014, 44, 377–404. (In Chinese) [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
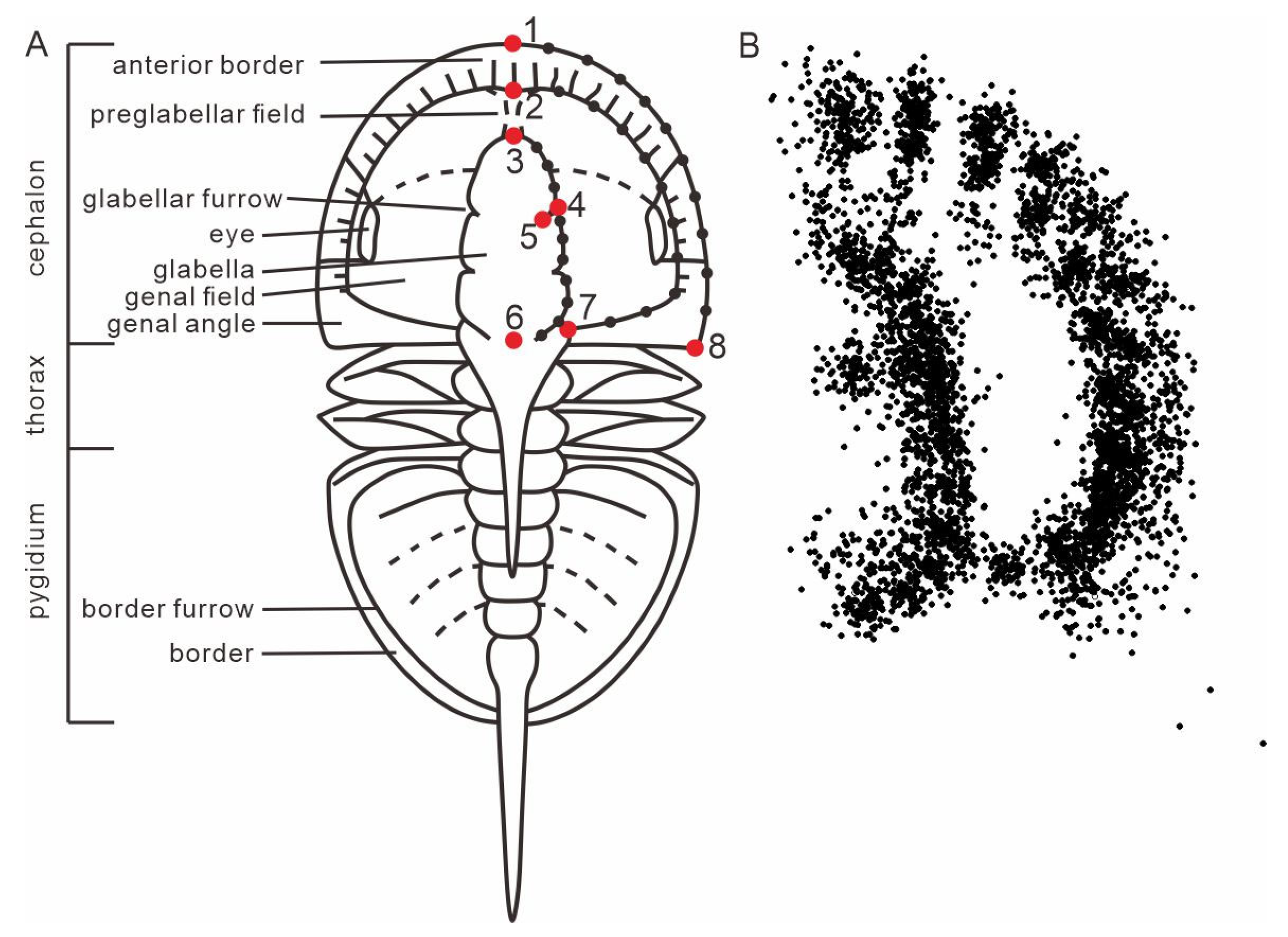
| Landmark Code | Description |
|---|---|
| 1 | Intersection between sagittal line and anterior cephalic margin |
| 2 | Intersection between sagittal line and anterior preglabellar field |
| 3 | Anterior-most point of sagittal glabellar length |
| 4 | Intersection between F3 and dorsal furrow |
| 5 | Point of the greatest extent medially of the F3 |
| 6 | Posterior-most point of sagittal glabellar length |
| 7 | Posterior-most end of genal field |
| 8 | Extremity of genal angle or spine |
| 9–18 | Sliders between landmarks 1 and 8, along the margin of cephalon |
| 19–28 | Sliders between landmarks 2 and 7, along the margin of genal field |
| 29–38 | Sliders between landmarks 3 and 6, along the margin of glabella |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Dai, T.; Chen, Y.; Xue, C.; Strotz, L.C. Morphological Evolution and Extinction of Eodiscids and Agnostoid Arthropods. Life 2025, 15, 38. https://doi.org/10.3390/life15010038
Li H, Dai T, Chen Y, Xue C, Strotz LC. Morphological Evolution and Extinction of Eodiscids and Agnostoid Arthropods. Life. 2025; 15(1):38. https://doi.org/10.3390/life15010038
Chicago/Turabian StyleLi, Huarui, Tao Dai, Yanlong Chen, Chunling Xue, and Luke C. Strotz. 2025. "Morphological Evolution and Extinction of Eodiscids and Agnostoid Arthropods" Life 15, no. 1: 38. https://doi.org/10.3390/life15010038
APA StyleLi, H., Dai, T., Chen, Y., Xue, C., & Strotz, L. C. (2025). Morphological Evolution and Extinction of Eodiscids and Agnostoid Arthropods. Life, 15(1), 38. https://doi.org/10.3390/life15010038