
The Sequence Characteristics and Binding Properties of the Odorant-Binding Protein SvelOBP1 from Sympiezomias velatus (Coleoptera: Curculionidae) to Jujube Volatiles

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Culture, Body Part Collection and RNA Isolation
2.2. cDNA Synthesis and Gene Cloning
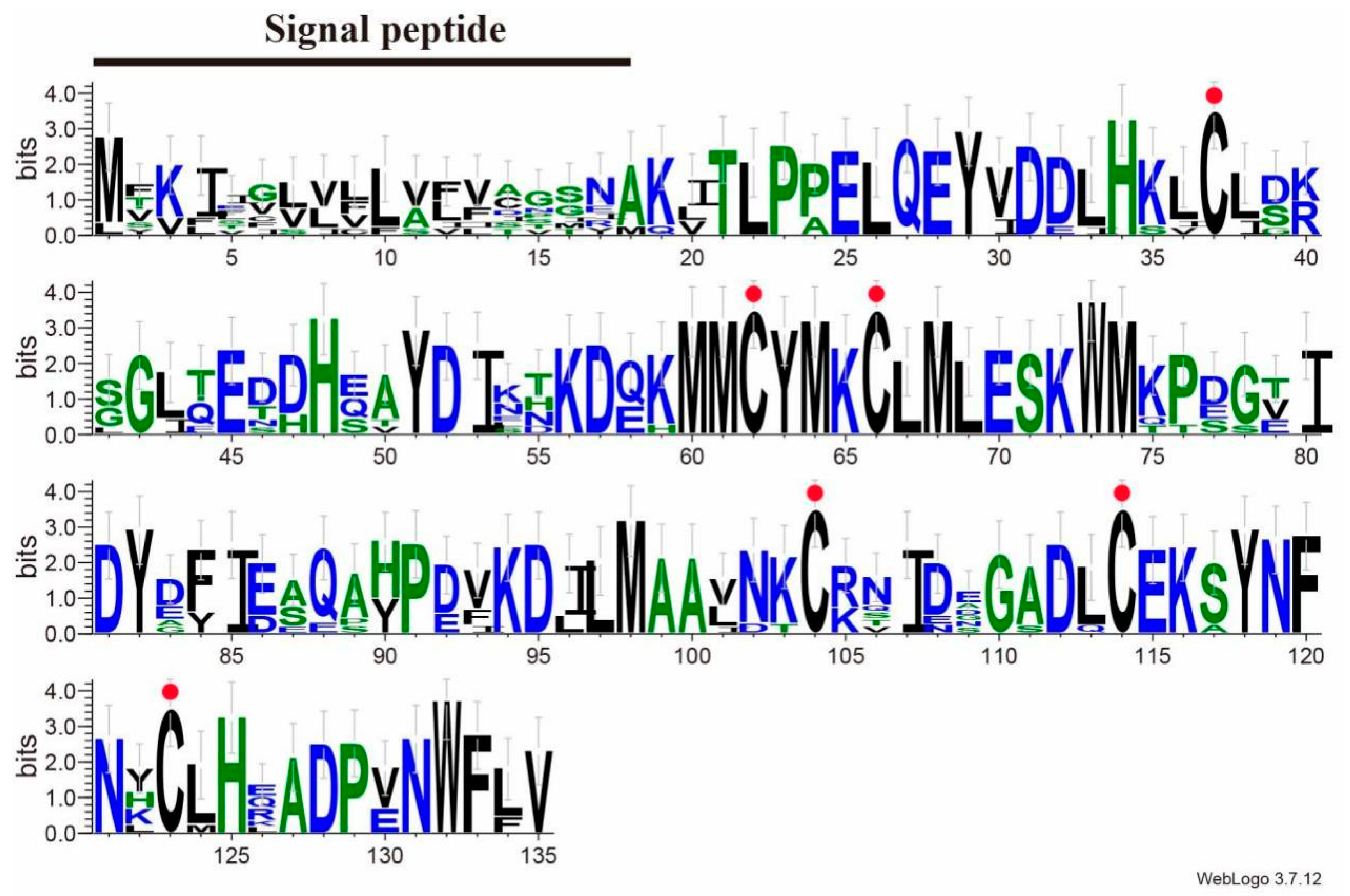
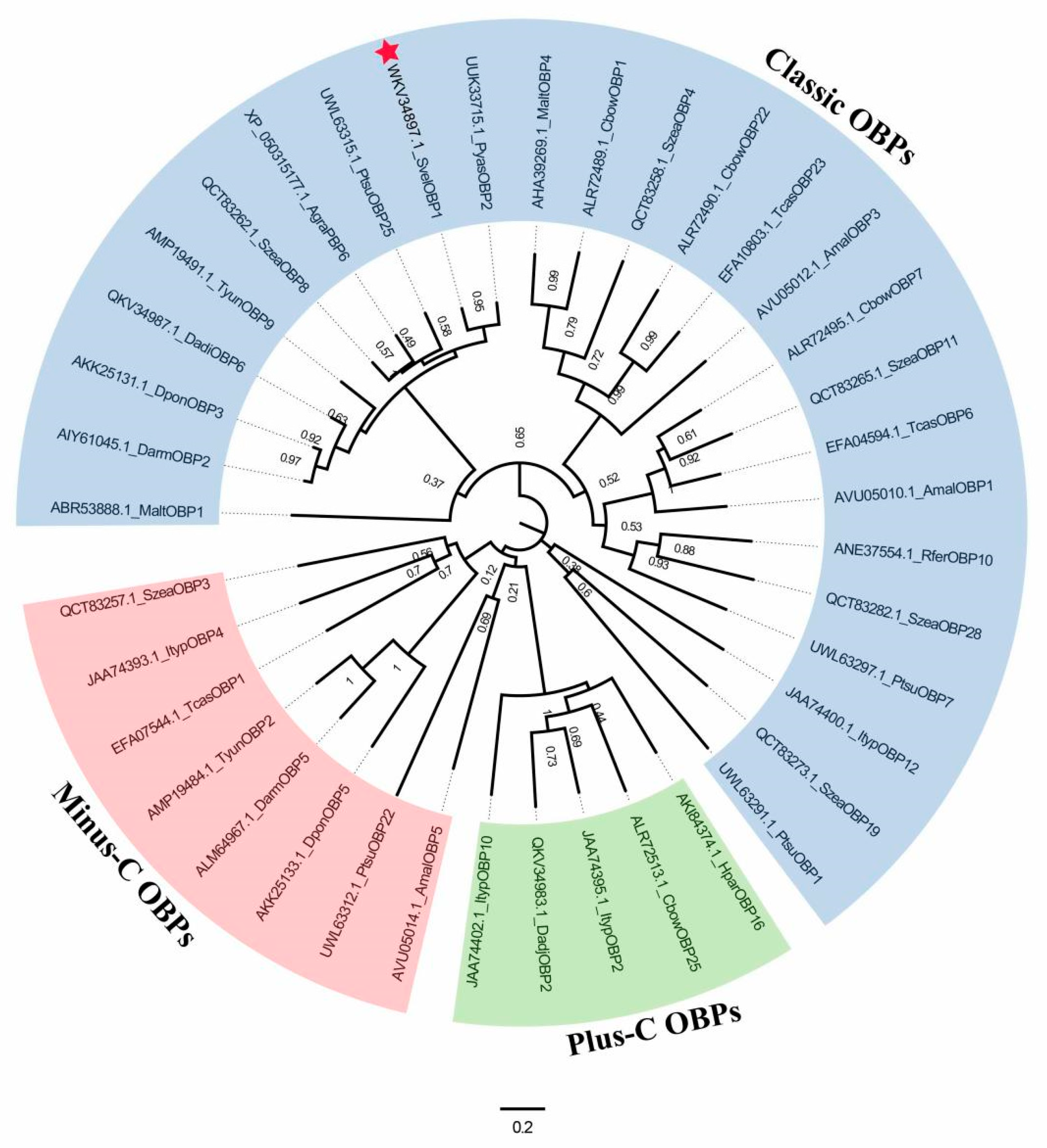
2.3. Sequence Alignment and Phylogenetic Analyses
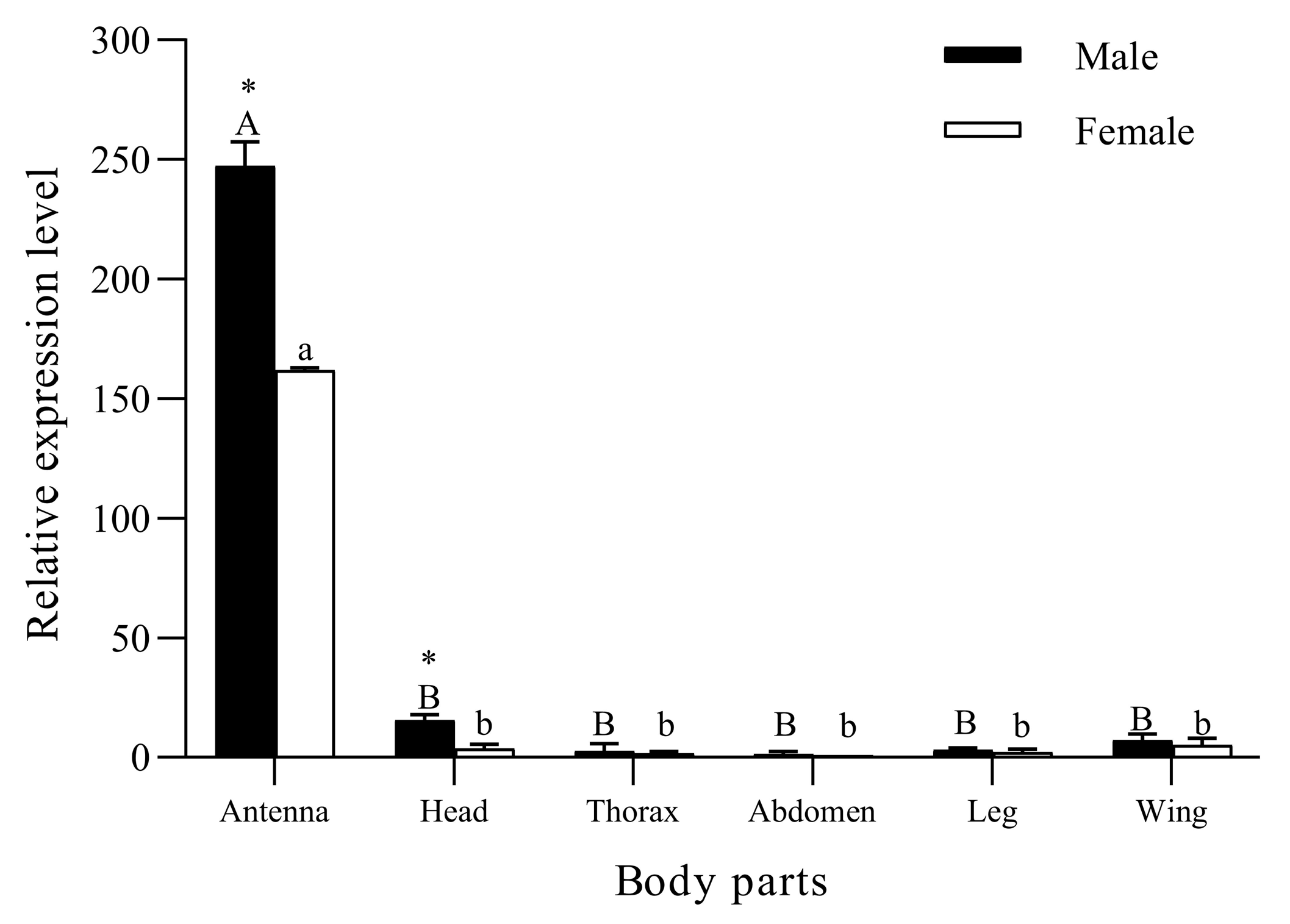
2.4. RT-qPCR Detection for Body Part Expression of SvelOBP1
2.5. Recombinant Plasmid Construction for E. coli Expression
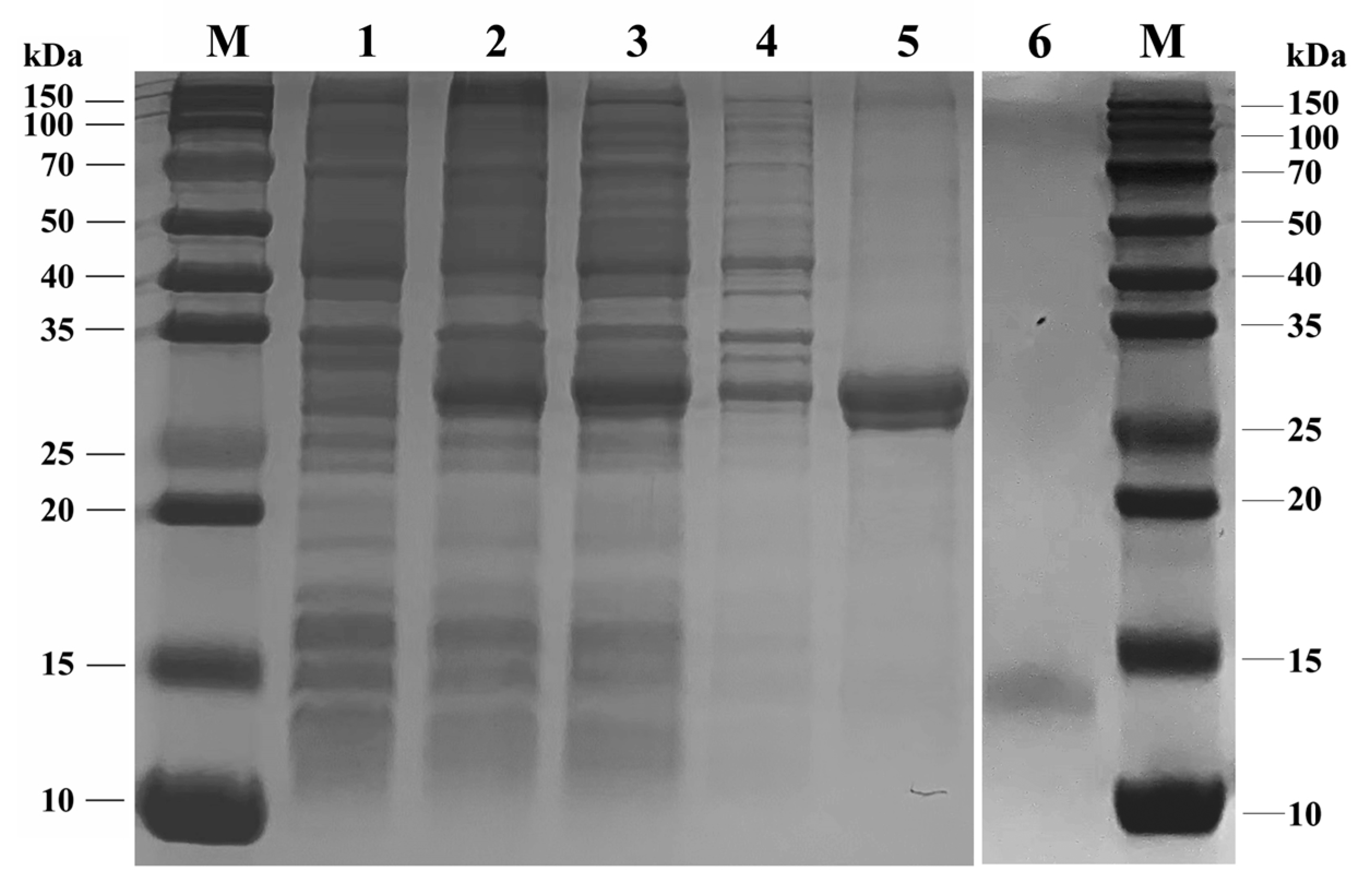
2.6. Expression and Purification of Recombinant SvelOBP1 Protein
2.7. Competitive Fluorescence Binding Assay
2.8. Structure Modeling and Molecular Docking
3. Results
3.1. Characterization of SvelOBP1 cDNA
3.2. Body Part Expression Profiles of SvelOBP1
3.3. Expression and Purification of SvelOBP1
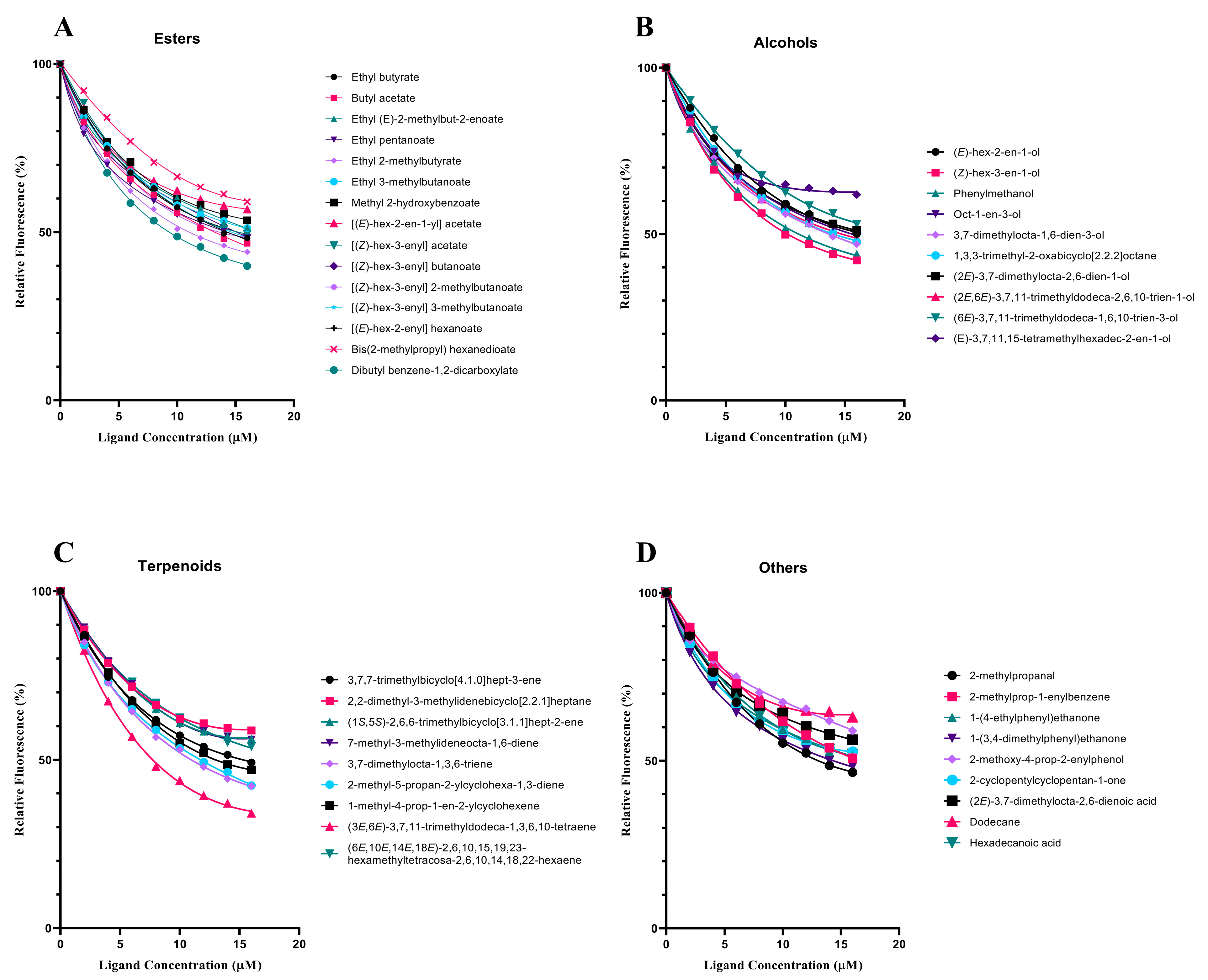
3.4. Fluorescence Competitive Binding Analyses of SvelOBP1
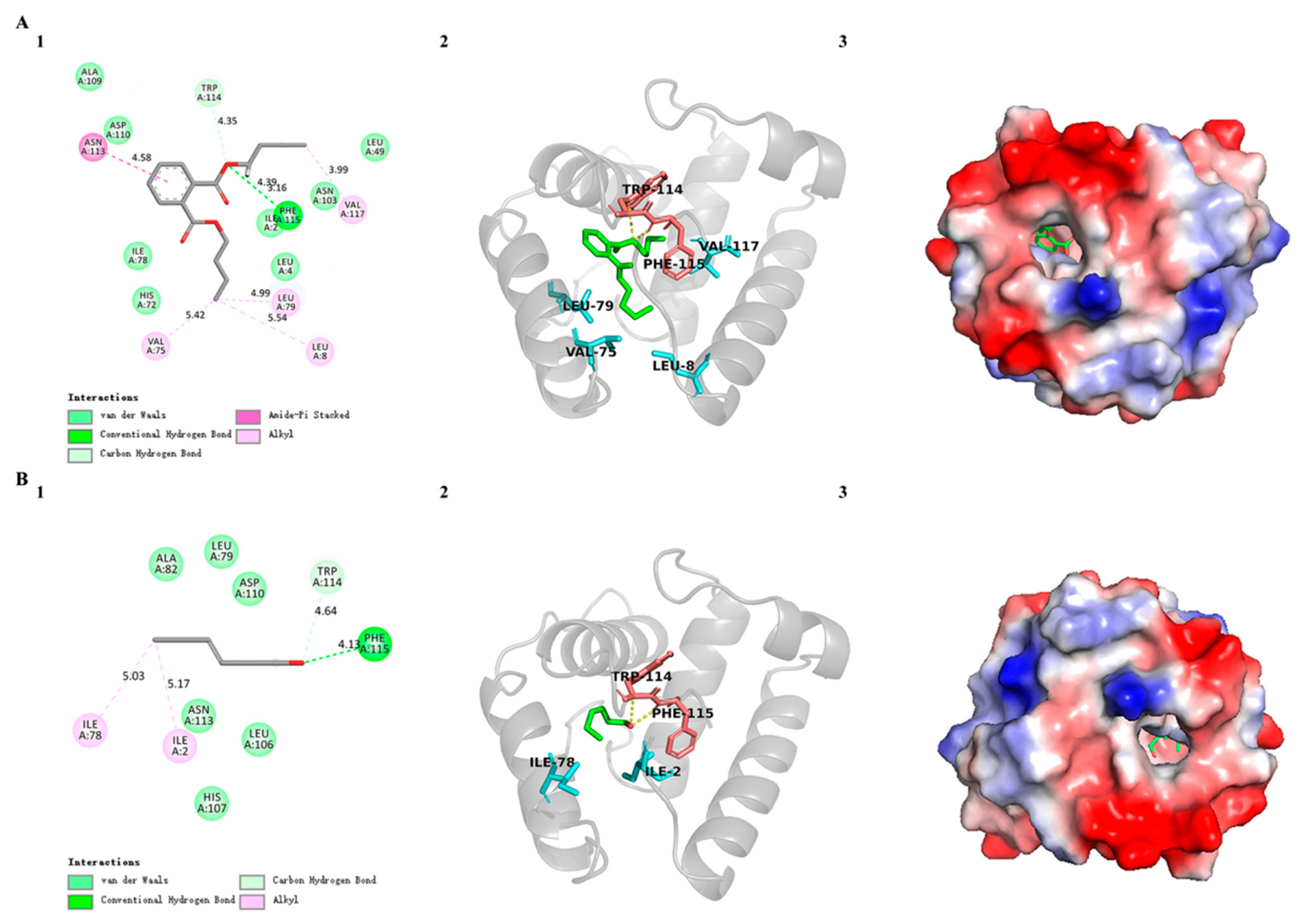
3.5. Three-Dimensional Modeling and Molecular Docking of SvelOBP1
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, Y.C.; Chen, Y.Q. Economic Insect Fauna of China: Coleoptera, Curculionidae(Ι); Science Press: Beijing, China, 1980; pp. 88–90. [Google Scholar]
- Yi, J. Biological characteristics and control of Sympiezomias velatus Chevrolat in Liaoning Province. Prot. For. Sci. Technol. 2015, 7, 105+107. [Google Scholar]
- Qu, Z.C. Integrated pest and disease control technology for pollution-free ecological jujube orchards. Parct. Forest. Tech. 2008, 10, 33–34. [Google Scholar]
- Wang, P. Experiment on the control effect of Sympiezomias velatus. Mod. Agri. Sci. Tech. 2022, 6, 68–69. [Google Scholar]
- Bruyne, M.; Baker, T.C. Odor detection in insects: Volatile codes. J. Chem. Ecol. 2008, 34, 882–897. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.W.; Sun, X.J.; Shi, Z.; An, X.K.; Khashaveh, A.; Li, Y.; Gu, S.H.; Zhang, Y.J. Identification and functional analysis of odorant-binding proteins provide new control strategies for Apolygus lucorum. Int. J. Biol. Macromol. 2023, 224, 1129–1141. [Google Scholar] [CrossRef]
- Sánchez-Gracia, A.; Vieira, F.G.; Rozas, J. Molecular Evolution of the Major Chemosensory Gene Families in Insects. Heredity 2009, 103, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Vosshall, L.B.; Amrein, H.; Morozov, P.S.; Rzhetsky, A.; Axel, R. A Spatial Map of Olfactory Receptor Expression in the Drosophila Antenna. Cell 1999, 96, 725–736. [Google Scholar] [CrossRef]
- Benton, R.; Vannice, K.S.; Gomez-Diaz, C.; Vosshall, L.B. Variant ionotropic glutamate receptors as chemosensory receptors in Drosophila. Cell 2009, 136, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Liu, L.; Fang, Y.; Feng, J. Expression of a sensory neuron membrane protein SNMP2 in olfactory sensilla of codling moth cydia pomonella (Lepidoptera: Tortricidae). J. Econ. Entomol. 2016, 109, 1907–1913. [Google Scholar] [CrossRef] [PubMed]
- Steinbrecht, R.A.; Laue, M.; Ziegelberger, G. Immunolocalization of pheromone-binding protein and general odorant-binding protein in olfactory sensilla of the silk moths Antheraea and Bombyx. Cell Tissue Res. 1995, 282, 203–217. [Google Scholar] [CrossRef]
- Leal, W.S. Odorant Reception in Insects: Roles of Receptors, Binding Proteins, and Degrading Enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef]
- Pelosi, P.; Zhou, J.J.; Ban, L.P.; Calvello, M. Soluble proteins in insect chemical communication. Cell. Mol. Life Sci. 2006, 63, 1658–1676. [Google Scholar] [CrossRef]
- Brito, N.F.; Moreira, M.F.; Melo, A.C. A look inside odorant-binding proteins in insect chemoreception. J. Insect Physiol. 2016, 95, 51–65. [Google Scholar] [CrossRef]
- Li, L.L.; Huang, J.R.; Xu, J.W.; Yao, W.C.; Yang, H.H.; Shao, L.; Zhang, H.R.; Dewer, Y.; Zhu, X.Y.; Zhang, Y.N. Ligand-binding properties of odorant-binding protein 6 in Athetis lepigone to sex pheromones and maize volatiles. Pest. Manag. Sci. 2022, 78, 52–62. [Google Scholar] [CrossRef]
- Hekmat-Scafe, D.S.; Scafe, C.R.; McKinney, A.J.; Tanouye, M.A. Genome-wide Analysis of the Odorant-Binding Protein Gene Family in Drosophila melanogaster. Genome Res. 2002, 12, 1357–1369. [Google Scholar] [CrossRef]
- Zhou, J.J. Odorant-binding proteins in insects. Vitam. Horm. 2010, 83, 241–272. [Google Scholar]
- Vogt, R.G.; Riddiford, L.M. Pheromone Binding and Inactivation by Moth Antennae. Nature 1981, 293, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Lun, X.Y.; Xu, X.X.; Zhang, Y.; Zhang, R.R.; Cao, Y.; Zhang, X.Z.; Jin, M.N.; Zhang, Z.Q.; Zhao, Y.H. An Antennae-Enriched Odorant-Binding Protein EonuOBP43 Mediate the Behavioral Response of the Tea Green Leafhopper, Empoasca onukii Matsuda to the Host and Nonhost Volatiles. J. Agric. Food Chem. 2023, 71, 20000–20010. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhang, L.; Qiu, Y.L.; Li, H.L. Research progress of olfactory binding proteins in insects. Aata Entomol. Sin. 2021, 64, 523–535. [Google Scholar]
- Lizana, L.; Mutis, A.; Quiroz, A.; Venthur, H. Insights Into Chemosensory Proteins From Non-Model Insects: Advances and Perspectives in the Context of Pest Management. Front. Physiol. 2022, 13, 924750. [Google Scholar] [CrossRef]
- Cui, X.N.; Liu, D.G.; Sun, K.K.; He, Y.; Shi, X.Q. Expression Profiles and Functional Characterization of Two Odorant-Binding Proteins from the Apple Buprestid Beetle Agrilus mali (Coleoptera: Buprestidae). J. Econ. Entomol. 2018, 111, 1420–1432. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Y.J.; Wang, Q.; Sun, L.; Zhang, Y.J. The Adelphocoris lineolatus OBP4: Support for evolutionary and functional divergence of a mirid pheromone-binding protein from that found in lepidopteran moths. Insect Sci. 2022, 29, 151–161. [Google Scholar] [CrossRef]
- Zhu, J.Q.; Wang, F.; Zhang, Y.J.; Yang, Y.T.; Hua, D.K. Odorant-binding Protein 10 From Bradysia odoriphaga (Diptera: Sciaridae) Binds Volatile Host Plant Compounds. J. Insect Sci. 2023, 23, 7. [Google Scholar] [CrossRef]
- Jia, C.; Mohamed, A.; Cattaneo, A.M.; Huang, X.H.; Keyhani, N.O.; Gu, M.Q.; Zang, L.S.; Zhang, W. Odorant-Binding Proteins and Chemosensory Proteins in Spodoptera frugiperda: From Genome-Wide Identification and Developmental Stage-Related Expression Analysis to the Perception of Host Plant Odors, Sex Pheromones, and Insecticides. Int. J. Mol. Sci. 2023, 24, 5595. [Google Scholar] [CrossRef]
- Crava, C.M.; Bobkov, Y.V.; Sollai, G.; Anfora, G.; Crnjar, R.; Cattaneo, A.M. Chemosensory Receptors in the Larval Maxilla of Papilio hospiton. Front. Ecol. Evol. 2022, 9, 795994. [Google Scholar] [CrossRef]
- Sandler, B.H.; Nikonova, L.; Leal, W.S.; Clardy, J. Sexual attraction in the silkworm moth: Structure of the pheromone-binding-protein-bombykol complex. Chem. Biol. 2000, 7, 143–151. [Google Scholar] [CrossRef]
- Leite, N.R.; Krogh, R.; Xu, W.; Ishida, Y.; Iulek, J.; Leal, W.S.; Oliva, G. Structure of an odorant-binding protein from the mosquito Aedes aegypti suggests a binding pocket covered by a pH-sensitive “lid”. PLoS ONE 2009, 4, e8006. [Google Scholar] [CrossRef]
- Yin, J.; Zhuang, X.; Wang, Q.; Cao, Y.; Zhang, S.; Xiao, C.; Li, K. Three amino acid residues of an odorant-binding protein are involved in binding odours in Loxostege sticticalis. Insect Mol. Biol. 2015, 24, 528–538. [Google Scholar] [CrossRef]
- Venthur, H.; Zhou, J.J. Odorant receptors and odorant-binding proteins as insect pest control targets: A comparative analysis. Front. Physiol. 2018, 9, 1163. [Google Scholar] [CrossRef]
- Li, X.; Li, W.J.; Sun, X.W.; Li, W.; Gao, H.Y.; Liu, T.X.; Qu, M.J. Candidate Chemosensory Genes Identified in the Adult Antennae of Sympiezomias velatus and Binding Property of Odorant-Binding Protein 15. Front. Physiol. 2022, 13, 907667. [Google Scholar] [CrossRef]
- Hong, B.; Chang, Q.; Zhai, Y.Y.; Ren, B.W.; Zhang, F. Functional Characterization of Odorant Binding Protein PyasOBP2 From the Jujube Bud Weevil, Pachyrhinus yasumatsui (Coleoptera: Curculionidae). Front. Physiol. 2022, 13, 900752. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Shen, C.; Xia, D.S.; Wang, J.; Tang, Q.F. Characterization of the Expression and Functions of Two Odorant-Binding Proteins of Sitophilus zeamais Motschulsky (Coleoptera: Curculionoidea). Insects 2019, 10, 409. [Google Scholar] [CrossRef]
- Metcalf, W.W.; Jiang, W.; Daniels, L.L.; Kim, S.K.; Haldimann, A.; Wanner, B.L. Conditionally replicative and conjugative plasmids carrying lacZ alpha for cloning, mutagenesis, and allele replacement in bacteria. Plasmid 1996, 35, 1–13. [Google Scholar] [CrossRef]
- Promega; Wagner, T. PCR Cloning with Blue/White Selection and Easy Insert Excision. 2018. Available online: https://www.protocols.io/view/pcr-cloning-with-blue-white-selection-and-easy-ins-t89erz6.html (accessed on 26 March 2023).
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.N.; Tang, B.W.; Zhou, T.L.; Yu, X.T.; Hu, M.F.; Dai, W. Involvement of Chemosensory Protein BodoCSP1 in Perception ofHost Plant Volatiles in Bradysia odoriphaga. J. Agric. Food Chem. 2021, 69, 10797–10806. [Google Scholar] [CrossRef]
- Zhang, L.W.; Zhao, M.X.; Aikeremu, F.; Huang, H.S.; You, M.S.; Zhao, Q. Involvement of three chemosensory proteins in perception of host plant volatiles in the tea green leafhopper, Empoasca onukii. Front. Physiol. 2023, 13, 1068543. [Google Scholar] [CrossRef]
- Yan, X.F.; Liu, Y.H.; Li, G.; Qiang, D.H.; Xu, Y.P. EAG and Olfactory Behavioral Responses of Scythropus yasumatsui to Volatiles from the Zizyphus jujube. Chin. J. Appl. Entomol. 2017, 54, 621–628. [Google Scholar]
- Yan, X.F.; Liu, Y.H.; Wang, Y.W.; Li, G.; Jing, R.; Yang, Y.J. EAG and Behavioral Responses of Scythropus yasumatsui (Coleoptera: Curculionidae) to Volatiles from the Common Jujube (Zizyphus jujuba). Acta Entomol. Sin. 2020, 63, 981–991. [Google Scholar]
- Li, L.L.; Xu, B.Q.; Li, C.Q.; Li, B.L.; Luo, K.; Li, G.W.; Chen, X.L. Functional disparity of four pheromone-binding proteins from the plum fruit moth Grapholita funebrana Treitscheke in detection of sex pheromone components. Int. J. Biol. Macromol. 2023, 225, 1267–1279. [Google Scholar] [CrossRef]
- Zhou, X.; Zheng, W.; Cui, G.C.; Du, Z.M.; Qian, Y.L.; Yang, S.M.; Liu, M.H.; Guo, J.X. Binding Properties of Odorant-Binding Protein 4 of Tirathaba rufivena to Areca catechu Volatiles. Plants 2022, 11, 167. [Google Scholar] [CrossRef]
- Cousins, K.R. Computer Review of Chemdraw Ultra 12.0. J. Am. Chem. Soc. 2011, 133, 8388. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.N.; Shan, S.; Yu, G.Y.; Wang, H.; Dhillo, K.H.; Khashaveh, A.; Zhang, F.; Zhang, Y.J. Identification of odorant-binding proteins and functional analysis of antenna-specific AplaOBP1 in the emerald ash borer, Agrilus planipennis. J. Pest. Sci. 2020, 93, 853–865. [Google Scholar] [CrossRef]
- Khuhro, S.A.; Liao, H.; Dong, X.T.; Yu, Q.; Yan, Q.; Dong, S.L. Two general odorant binding proteins display high bindings to both host plant volatiles and sex pheromones in a pyralid moth Chilo suppressalis (Lepidoptera: Pyralidae). J. Asia Pac. Entomol. 2017, 20, 521–528. [Google Scholar] [CrossRef]
- Huang, G.Z.; Liu, J.T.; Zhou, J.J.; Wang, Q.; Dong, J.Z.; Zhang, Y.J.; Li, X.C.; Li, J.; Gu, S.H. Expressional and functional comparisons of two general odorant binding proteins in Agrotis ipsilon. Insect Biochem. Mol. Biol. 2018, 98, 34–47. [Google Scholar] [CrossRef]
- Yin, J.; Cao, Y.Z.; Luo, L.Z.; Hu, Y. Oviposition preference of the meadow moth, Loxostege sticticalis L., on different host plants and its chemical mechanism. Acta Ecol. Sin. 2005, 25, 1844–1852. [Google Scholar]
- Li, E.T.; Wu, H.J.; Qin, J.H.; Luo, J.; Li, K.B.; Gao, Y.Z.; Zhang, S.; Peng, Y.; Yin, J. Involvement of Holotrichia parallela odorant-binding protein 3 in the localization of oviposition sites. Int. J. Biol. Macromol. 2023, 242, 124744. [Google Scholar] [CrossRef]
- Abdukerim, R.; Cao, Y.Z.; Zhang, S.; Yin, J.; Li, X.; Li, K.B. Feeding preference and taxis behavior of adult Holotrichia oblita (Coleoptera: Scarabaeidae) on three plants. Acta Entomol. Sin. 2018, 61, 585–595. [Google Scholar]
- Wei, H.S.; Qin, J.H.; Cao, Y.Z.; Li, K.B.; Yin, J. Two classic OBPs modulate the responses of female Holotrichia oblita to three major ester host plant volatiles. Insect Mol. Biol. 2021, 30, 390–399. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, Q.; Li, H.; Sun, L.; Zhang, D.; Zhang, Y.J. Sensilla localization and sex pheromone recognition of odorant binding protein OBP4 in the mirid plant bug Adelphocoris lineolatus (Goeze). J. Insect Physiol. 2020, 121, 104012. [Google Scholar] [CrossRef]
- Liu, N.Y.; Yang, F.; Yang, K.; He, P.; Niu, X.H.; Xu, W.; Anderson, A.; Dong, S.L. Two subclasses of odorant-binding proteins in Spodoptera exigua display structural conservation and functional divergence. Insect Mol. Biol. 2015, 24, 167–182. [Google Scholar] [CrossRef]
- Pelosi, P.; Lovinella, L.; Zhu, J.; Wang, G.R.; Dani, F.R. Beyond chemoreception: Diverse tasks of soluble olfactory proteins in insects. Biol. Rev. 2018, 93, 184–200. [Google Scholar] [CrossRef]
- Mao, Y.; Xu, X.; Xu, W.; Ishida, Y.; Leal, W.S.; Ames, J.B.; Clardy, J. Crystal and solution structures of an odorant-binding protein from the southern house mosquito complexed with an oviposition pheromone. Proc. Natl. Acad. Sci. USA 2010, 107, 19102–19107. [Google Scholar] [CrossRef]
- Zhuang, X.; Wang, Q.; Wang, B.; Zhong, T.; Cao, Y.; Li, K.; Yin, J. Prediction of the key binding site of odorant-binding protein of Holotrichia oblita Faldermann (Coleoptera: Scarabaeida). Insect Mol. Biol. 2014, 23, 381–390. [Google Scholar] [CrossRef]
- Liu, X.Q.; Jiang, H.B.; Fan, J.Y.; Liu, T.Y.; Meng, L.M.; Liu, Y.; Yu, H.Z.; Dou, W.; Wang, J.J. An odorant-binding protein of Asian citrus psyllid, Diaphorina citri, participates in the response of host plant volatiles. Pest. Manag. Sci. 2021, 77, 3068–3079. [Google Scholar] [CrossRef]
- Qu, C.; Yang, Z.K.; Wang, S.; Zhao, H.P.; Li, F.Q.; Yang, X.L.; Luo, C. Binding affinity characterization of four antennae-enriched odorant-binding proteins from Harmonia axyridis (Coleoptera: Coccinellidae). Front. Physiol. 2022, 13, 188. [Google Scholar] [CrossRef]
- Yang, L.; Tian, X.L.; Gui, L.Y.; Wang, F.L.; Zhang, G.H. Key Amino Acid Residues Involved in Binding Interactions between Bactrocera minax Odorant-Binding Protein 3 (BminOBP3) and Undecanol. Insects 2023, 14, 745. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequences (5′-3′) | Fragment Length (bp) |
|---|---|---|
| For gene cloning | ||
| SvelOBP1-F | ATGTCTAAAATTATAGGTCTGTTGG | 408 |
| SvelOBP1-R | TTASACRAAGAACCARTTCAC | |
| For qRT-PCR | ||
| SvelOBP1-qF | GCTCCAAGAATACGTCGATGA | 168 |
| SvelOBP1-qR | TCGATTTCACCACTGGGTTT | |
| EF1α-qF | AACCACCATACAGCGAGTCC | 128 |
| EF1α-qR | CATGTTGTCTCCATGCCATC | |
| βactin-qF | CTACGAAGGTTACGCTCTTCCC | 123 |
| βactin-qR | CTTTCAGCTGTGGTGGTGAAT | |
| For prokaryotic expression | ||
| SvelOBP1-eF | CGGGATCCAAGATAACTTTACCTCCTGAGCTCC | 374 |
| SvelOBP1-eR | CCGGAATTCCGGTTASACRAAGAACCAGTTCACAGG |
| Ligands | CAS No. | IC50 (μM) | Ki (μM) |
|---|---|---|---|
| Esters | |||
| Ethyl butyrate | 105-54-4 | 14.19 ± 0.16 | 11.79 ± 0.13 |
| Butyl acetate | 123-86-4 | 13.16 ± 0.47 | 10.94 ± 0.39 |
| Ethyl (E)-2-methylbut-2-enoate | 5837-78-5 | >16 | − |
| Ethyl pentanoate | 539-82-2 | 14.12 ± 0.31 | 11.73 ± 0.26 |
| Ethyl 2-methylbutyrate | 7452-79-1 | 11.28 ± 0.10 | 9.37 ± 0.08 |
| Ethyl 3-methylbutanoate | 108-64-5 | >16 | − |
| Methyl 2-hydroxybenzoate | 119-36-8 | >16 | − |
| [(E)-hex-2-en-1-yl] acetate | 2497-18-9 | >16 | − |
| [(Z)-hex-3-enyl] acetate | 3681-71-8 | 14.45 ± 0.11 | 12.01 ± 0.09 |
| [(Z)-hex-3-enyl] butanoate | 16491-36-4 | 15.52 ± 0.90 | 12.90 ± 0.75 |
| [(Z)-hex-3-enyl] 2-methylbutanoate | 53398-85-9 | 13.42 ± 0.20 | 11.15 ± 0.16 |
| [(Z)-hex-3-enyl] 3-methylbutanoate | 35154-45-1 | 14.37 ± 0.16 | 11.94 ± 0.13 |
| [(E)-hex-2-enyl] hexanoate | 53398-86-0 | 14.51 ± 0.06 | 12.06 ± 0.05 |
| Bis(2-methylpropyl) hexanedioate | 141-04-8 | >16 | − |
| Dibutyl benzene-1,2-dicarboxylate | 84-74-2 | 9.60 ± 0.09 | 6.66 ± 0.03 |
| Alcohols | |||
| (E)-hex-2-en-1-ol | 928-95-0 | 15.44 ± 0.14 | 12.83 ± 0.12 |
| (Z)-hex-3-en-1-ol | 928-96-1 | 10.51 ± 0.11 | 7.98 ± 0.08 |
| Phenylmethanol | 100-51-6 | 11.31 ± 0.27 | 9.40 ± 0.23 |
| Oct-1-en-3-ol | 3391-86-4 | 15.18 ± 0.13 | 12.61 ± 0.11 |
| 3,7-dimethylocta-1,6-dien-3-ol | 78-70-6 | 13.59 ± 0.16 | 11.29 ± 0.13 |
| 1,3,3-trimethyl-2-oxabicyclo [2.2.2]octane | 470-82-6 | 13.81 ± 0.13 | 11.48 ± 0.11 |
| (2E)-3,7-dimethylocta-2,6-dien-1-ol | 106-24-1 | >16 | − |
| (2E,6E)-3,7,11-trimethyldodeca-2,6,10-trien-1-ol | 4602-84-0 | 14.36 ± 0.17 | 11.93 ± 0.14 |
| (6E)-3,7,11-trimethyldodeca-1,6,10-trien-3-ol | 7212-44-4 | >16 | − |
| (E)-3,7,11,15-tetramethylhexadec-2-en-1-ol | 7541-49-3 | >16 | − |
| Terpenoids | |||
| 3,7,7-trimethylbicyclo [4.1.0]hept-3-ene | 13466-78-9 | 14.56 ± 0.07 | 12.10 ± 0.06 |
| 2,2-dimethyl-3-methylidenebicyclo [2.2.1]heptane | 79-92-5 | >16 | − |
| (1S,5S)-2,6,6-trimethylbicyclo [3.1.1]hept-2-ene | 7785-26-4 | >16 | − |
| 7-methyl-3-methylideneocta-1,6-diene | 123-35-3 | >16 | − |
| 3,7-dimethylocta-1,3,6-triene | 13877-91-3 | 11.13 ± 0.06 | 9.25 ± 0.05 |
| 2-methyl-5-propan-2-ylcyclohexa-1,3-diene | 99-83-2 | 11.69 ± 0.07 | 9.71 ± 0.06 |
| 1-methyl-4-prop-1-en-2-ylcyclohexene | 7705-14-8 | 13.11 ± 0.02 | 10.90 ± 0.01 |
| (3E,6E)-3,7,11-trimethyldodeca-1,3,6,10-tetraene | 502-61-4 | 8.01 ± 0.04 | 8.73 ± 0.09 |
| (6E,10E,14E,18E)-2,6,10,15,19,23-hexamethyltetracosa-2,6,10,14,18,22-hexaene | 111-02-4 | >16 | − |
| Others | |||
| 1-(4-ethylphenyl)ethanone | 937-30-4 | 13.13 ± 0.16 | 10.91 ± 0.13 |
| 1-(3,4-dimethylphenyl)ethanone | 3637-01-2 | >16 | − |
| (2E)-3,7-dimethylocta-2,6-dienoic acid | 459-80-3 | >16 | − |
| Hexadecanoic acid | 57-10-3 | 14.09 ± 0.33 | 11.71 ± 0.27 |
| 2-cyclopentylcyclopentan-1-one | 4884-24-6 | >16 | − |
| 2-methylpropanal | 78-84-2 | >16 | − |
| 2-methoxy-4-prop-2-enylphenol | 97-53-0 | >16 | − |
| Dodecane | 112-40-3 | >16 | − |
| 2-methylprop-1-enylbenzene | 768-49-0 | >16 | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhai, Y.; Zhang, F.; Tian, T.; Yang, Y.; Li, Y.; Ren, B.; Hong, B. The Sequence Characteristics and Binding Properties of the Odorant-Binding Protein SvelOBP1 from Sympiezomias velatus (Coleoptera: Curculionidae) to Jujube Volatiles. Life 2024, 14, 192. https://doi.org/10.3390/life14020192
Zhai Y, Zhang F, Tian T, Yang Y, Li Y, Ren B, Hong B. The Sequence Characteristics and Binding Properties of the Odorant-Binding Protein SvelOBP1 from Sympiezomias velatus (Coleoptera: Curculionidae) to Jujube Volatiles. Life. 2024; 14(2):192. https://doi.org/10.3390/life14020192
Chicago/Turabian StyleZhai, Yingyan, Feng Zhang, Tianqi Tian, Yiwei Yang, Yang Li, Bowen Ren, and Bo Hong. 2024. "The Sequence Characteristics and Binding Properties of the Odorant-Binding Protein SvelOBP1 from Sympiezomias velatus (Coleoptera: Curculionidae) to Jujube Volatiles" Life 14, no. 2: 192. https://doi.org/10.3390/life14020192
APA StyleZhai, Y., Zhang, F., Tian, T., Yang, Y., Li, Y., Ren, B., & Hong, B. (2024). The Sequence Characteristics and Binding Properties of the Odorant-Binding Protein SvelOBP1 from Sympiezomias velatus (Coleoptera: Curculionidae) to Jujube Volatiles. Life, 14(2), 192. https://doi.org/10.3390/life14020192