

Mechanisms of Epigenetic Inheritance in Post-Traumatic Stress Disorder


Abstract
1. Introduction
2. Epigenetic Mechanisms
2.1. DNA Methylation
2.2. Histone Modification
2.3. Non-Coding RNAs (ncRNA)
3. Identifying Epigenetic Markers of PTSD
4. Intergenerational and Transgenerational Inheritance of Epigenetic Markers
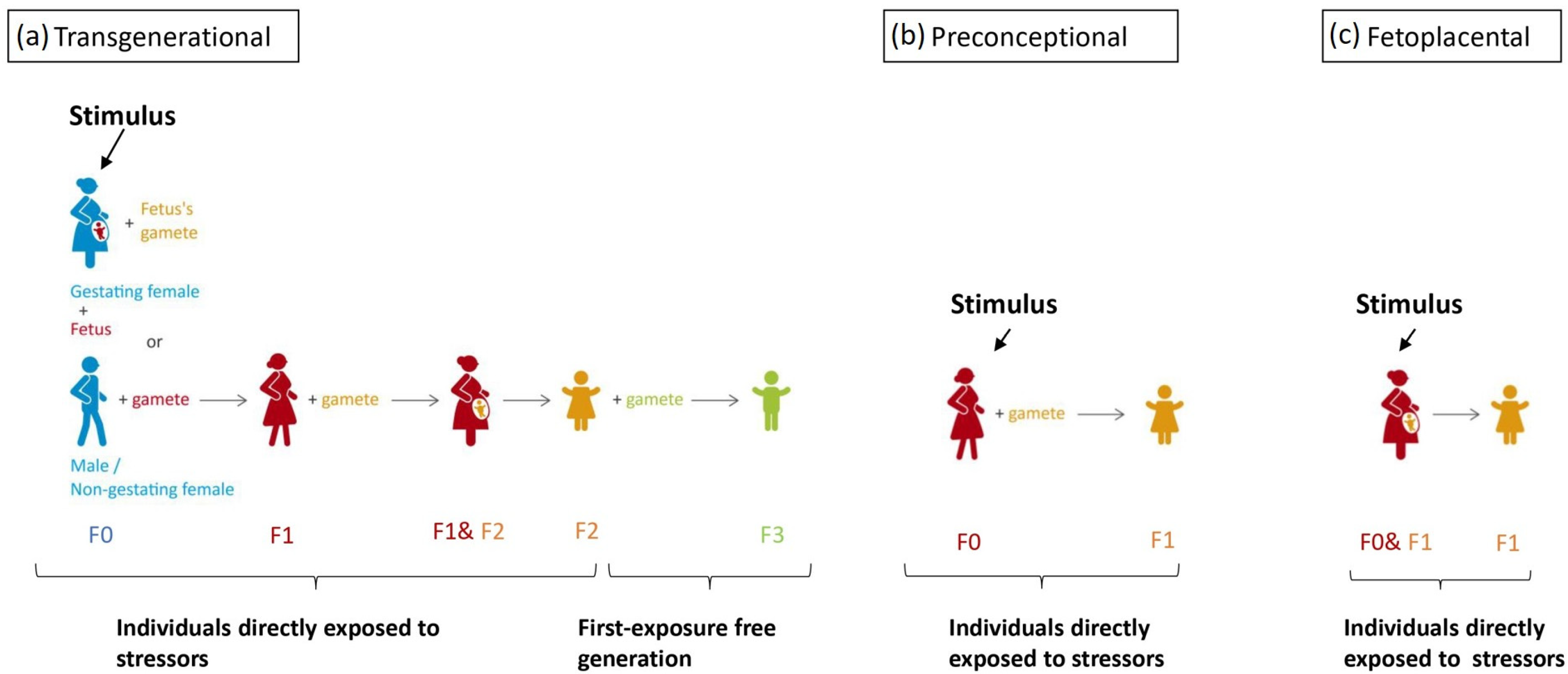
5. Different Timings of Trauma Exposure and Intergenerational Inheritance Studies
5.1. Post-Natal Exposure
5.2. In Utero Exposure
5.3. Preconceptional Inheritance and Transgenerational Inheritance
5.3.1. Preconceptional Inheritance
5.3.2. Transgenerational Inheritance
6. Cultural Level PTSD
7. Future Directions and Use
8. Closing Statements
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Varela, R.B.; Cararo, J.H.; Tye, S.J.; Carvalho, A.F.; Valvassori, S.S.; Fries, G.R.; Quevedo, J. Contributions of epigenetic inheritance to the predisposition of major psychiatric disorders: Theoretical framework, evidence, and implications. Neurosci. Biobehav. Rev. 2022, 135, 104579. [Google Scholar] [CrossRef]
- American Psychiatric Association. Desk Reference to the Diagnostic Criteria from DSM-5; American Psychiatric Association: Arlington, TX, USA, 2013. [Google Scholar]
- Campbell-Sills, L.; Sun, X.; Kessler, R.C.; Ursano, R.J.; Jain, S.; Stein, M.B. Exposure to Bullying or Hazing During Deployment and Mental Health Outcomes Among US Army Soldiers. JAMA Netw. Open 2023, 6, e2252109. [Google Scholar] [CrossRef] [PubMed]
- El-Khoury, F.; Rieckmann, A.; Bengtsson, J.; Melchior, M.; Rod, N.H. Childhood adversity trajectories and PTSD in young adulthood: A nationwide Danish register-based cohort study of more than one million individuals. J. Psychiatr. Res. 2021, 136, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.K.; Feng, M.; Fang, Y.; Lv, L.; Sun, G.L.; Yang, S.L.; Guo, P.; Cheng, S.F.; Qian, M.C.; Chen, H.X. Psychological trauma, posttraumatic stress disorder and trauma-related depression: A mini-review. World J. Psychiatry 2023, 13, 331–339. [Google Scholar] [CrossRef] [PubMed]
- John, R.; Rougelle, C. Developmental epigenetics, phenotype and the flexible epigenome. Front. Cell Dev. Biol. 2018, 6, 130. [Google Scholar] [CrossRef] [PubMed]
- Heijmans, B.T.; Tobi, E.W.; Stein, A.D.; Putter, H.; Blauw, G.J.; Susser, E.S.; Slagboom, P.E.; Lumey, L.H. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 17046–17049. [Google Scholar] [CrossRef] [PubMed]
- Yehuda, R.; Daskalakis, N.P.; Bierer, L.M.; Bader, H.N.; Klengel, T.; Holsboer, F.; Binder, E.B. Holocaust Exposure Induced Intergenerational Effects on FKBP5 Methylation. Biol. Psychiatry 2016, 80, 372–380. [Google Scholar] [CrossRef]
- Yehuda, R.; Lehrner, A. Intergenerational transmission of trauma effects: Putative role of epigenetic mechanisms. World Psychiatry 2018, 17, 243–257. [Google Scholar] [CrossRef]
- Cao-Lei, L.; Massart, R.; Suderman, M.J.; Machnes, Z.; Elgbeili, G.; Laplante, D.P.; Szyf, M.; King, S. DNA methylation signatures triggered by prenatal maternal stress exposure to a natural disaster: Project Ice Storm. PLoS ONE 2014, 9, e107653. [Google Scholar] [CrossRef]
- Yehuda, R.; Engel, S.M.; Brand, S.R.; Seckl, J.; Marcus, S.M.; Berkowitz, G.S. Transgenerational effects of posttraumatic stress disorder in babies of mothers exposed to the World Trade Center attacks during pregnancy. J. Clin. Endocrinol. Metab. 2005, 90, 4115–4118. [Google Scholar] [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Youssef, N.A. Potential Societal and Cultural Implications of Transgenerational Epigenetic Methylation of Trauma and PTSD: Pathology or Resilience? Yale J. Biol. Med. 2022, 95, 171–174. [Google Scholar] [PubMed]
- Wu, H.; Sun, Y.E. Epigenetic regulation of stem cell differentiation. Pediatr. Res. 2006, 59 Pt 2, 21R–25R. [Google Scholar] [CrossRef]
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, C. Coding or Noncoding, the Converging Concepts of RNAs. Front. Genet. 2019, 10, 496. [Google Scholar] [CrossRef] [PubMed]
- Kaikkonen, M.U.; Lam, M.T.; Glass, C.K. Non-coding RNAs as regulators of gene expression and epigenetics. Cardiovasc. Res. 2011, 90, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Shlyakhtina, Y.; Moran, K.L.; Portal, M.M. Asymmetric Inheritance of Cell Fate Determinants: Focus on RNA. Noncoding RNA 2019, 5, 38. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, J.M. Epigenome-wide association studies (EWAS): Past, present, and future. Methods Mol. Biol. 2015, 1238, 51–63. [Google Scholar]
- Mill, J.; Heijmans, B.T. From promises to practical strategies in epigenetic epidemiology. Nat. Rev. Genet. 2013, 14, 585–594. [Google Scholar] [CrossRef]
- Beck, S.; Rakyan, V.K. The methylome: Approaches for global DNA methylation profiling. Trends Genet. 2008, 24, 231–237. [Google Scholar] [CrossRef]
- Michels, K.B.; Binder, A.M.; Dedeurwaerder, S.; Epstein, C.B.; Greally, J.M.; Gut, I.; Houseman, E.A.; Izzi, B.; Kelsey, K.T.; Meissner, A.; et al. Recommendations for the design and analysis of epigenome-wide association studies. Nat. Methods 2013, 10, 949–955. [Google Scholar] [CrossRef] [PubMed]
- Tanić, M. Epigenome-wide association study (EWAS): Methods and applications. In Epigenetics Methods; Elsevier: Amsterdam, The Netherlands, 2020; pp. 591–613. [Google Scholar]
- Miller, O.; Shakespeare-Finch, J.; Bruenig, D.; Mehta, D. DNA methylation of NR3C1 and FKBP5 is associated with posttraumatic stress disorder, posttraumatic growth, and resilience. Psychol. Trauma Theory Res. Pract. Policy 2020, 12, 750. [Google Scholar] [CrossRef]
- Mehta, D.; Pelzer, E.S.; Bruenig, D.; Lawford, B.; McLeay, S.; Morris, C.P.; Gibson, J.N.; Young, R.M.; Voisey, J.; Initiative, P. DNA methylation from germline cells in veterans with PTSD. J. Psychiatr. Res. 2019, 116, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Labonte, B.; Azoulay, N.; Yerko, V.; Turecki, G.; Brunet, A. Epigenetic modulation of glucocorticoid receptors in posttraumatic stress disorder. Transl. Psychiatry 2014, 4, e368. [Google Scholar] [CrossRef] [PubMed]
- Blacker, C.J.; Frye, M.A.; Morava, E.; Kozicz, T.; Veldic, M. A Review of Epigenetics of PTSD in Comorbid Psychiatric Conditions. Genes 2019, 10, 140. [Google Scholar] [CrossRef] [PubMed]
- Howie, H.; Rijal, C.M.; Ressler, K.J. A review of epigenetic contributions to post-traumatic stress disorder. Dialogues Clin. Neurosci. 2022, 21, 417–428. [Google Scholar] [CrossRef]
- Logue, M.W.; Miller, M.W.; Wolf, E.J.; Huber, B.R.; Morrison, F.G.; Zhou, Z.; Zheng, Y.; Smith, A.K.; Daskalakis, N.P.; Ratanatharathorn, A. An epigenome-wide association study of posttraumatic stress disorder in US veterans implicates several new DNA methylation loci. Clin. Epigenetics 2020, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.K.; Ratanatharathorn, A.; Maihofer, A.X.; Naviaux, R.K.; Aiello, A.E.; Amstadter, A.B.; Ashley-Koch, A.E.; Baker, D.G.; Beckham, J.C.; Boks, M.P.; et al. Epigenome-wide meta-analysis of PTSD across 10 military and civilian cohorts identifies methylation changes in AHRR. Nat. Commun. 2020, 11, 5965. [Google Scholar] [CrossRef]
- Snijders, C.; Maihofer, A.X.; Ratanatharathorn, A.; Baker, D.G.; Boks, M.P.; Geuze, E.; Jain, S.; Kessler, R.C.; Pishva, E.; Risbrough, V.B.; et al. Longitudinal epigenome-wide association studies of three male military cohorts reveal multiple CpG sites associated with post-traumatic stress disorder. Clin. Epigenetics 2020, 12, 11. [Google Scholar] [CrossRef]
- Nothling, J.; Abrahams, N.; Toikumo, S.; Suderman, M.; Mhlongo, S.; Lombard, C.; Seedat, S.; Hemmings, S.M.J. Genome-wide differentially methylated genes associated with posttraumatic stress disorder and longitudinal change in methylation in rape survivors. Transl. Psychiatry 2021, 11, 594. [Google Scholar] [CrossRef]
- Hjort, L.; Rushiti, F.; Wang, S.J.; Fransquet, P.; PKrasniqi, S.; Çarkaxhiu, S.I.; Arifaj, D.; Xhemaili, V.D.; Salihu, M.; ALeku, N.; et al. Intergenerational effects of maternal post-traumatic stress disorder on offspring epigenetic patterns and cortisol levels. Epigenomics 2021, 13, 967–980. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, C.; D’Errico, N.; Stees, J.; Hughes, D. Methylation changes at NR3C1 in newborns associate with maternal prenatal stress exposure and newborn birth weight. Epigenetics 2012, 7, 853–857. [Google Scholar] [CrossRef] [PubMed]
- Rodney, N.C.; Mulligan, C.J. A biocultural study of the effects of maternal stress on mother and newborn health in the D emocratic R epublic of C ongo. Am. J. Phys. Anthropol. 2014, 155, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Vinkers, C.H.; Geuze, E.; van Rooij, S.J.H.; Kennis, M.; Schur, R.R.; Nispeling, D.M.; Smith, A.K.; Nievergelt, C.M.; Uddin, M.; Rutten, B.P.F.; et al. Successful treatment of post-traumatic stress disorder reverses DNA methylation marks. Mol. Psychiatry 2021, 26, 1264–1271. [Google Scholar] [CrossRef] [PubMed]
- Skinner, M.K. What is an epigenetic transgenerational phenotype?: F3 or F2. Reprod. Toxicol. 2008, 25, 2–6. [Google Scholar] [CrossRef]
- Lacal, I.; Ventura, R. Epigenetic Inheritance: Concepts, Mechanisms and Perspectives. Front. Mol. Neurosci. 2018, 11, 292. [Google Scholar] [CrossRef]
- Saab, B.J.; Mansuy, I.M. Neurobiological disease etiology and inheritance: An epigenetic perspective. J. Exp. Biol. 2014, 217 Pt 1, 94–101. [Google Scholar] [CrossRef]
- Heard, E.; Martienssen, R.A. Transgenerational epigenetic inheritance: Myths and mechanisms. Cell 2014, 157, 95–109. [Google Scholar] [CrossRef]
- Collins, N.; Roth, T.L. Intergenerational transmission of stress-related epigenetic regulation. In Developmental Human Behavioral Epigenetics; Elsevier: Amsterdam, The Netherlands, 2021; pp. 119–141. [Google Scholar]
- McGowan, P.O.; Sasaki, A.; D’Alessio, A.C.; Dymov, S.; Labonte, B.; Szyf, M.; Turecki, G.; Meaney, M.J. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat. Neurosci. 2009, 12, 342–348. [Google Scholar] [CrossRef]
- Jiang, S.; Postovit, L.; Cattaneo, A.; Binder, E.B.; Aitchison, K.J. Epigenetic Modifications in Stress Response Genes Associated With Childhood Trauma. Front. Psychiatry 2019, 10, 808. [Google Scholar] [CrossRef]
- Ullmann, E.; Licinio, J.; Barthel, A.; Petrowski, K.; Stalder, T.; Bornstein, S.R.; Kirschbaum, C. Persistent LHPA Activation in German Individuals Raised in an Overprotective Parental Behavior. Sci. Rep. 2017, 7, 2778. [Google Scholar] [CrossRef] [PubMed]
- Yahyavi, S.T.; Zarghami, M.; Marwah, U. A review on the evidence of transgenerational transmission of posttraumatic stress disorder vulnerability. Braz. J. Psychiatry 2014, 36, 89–94. [Google Scholar] [CrossRef]
- Perroud, N.; Rutembesa, E.; Paoloni-Giacobino, A.; Mutabaruka, J.; Mutesa, L.; Stenz, L.; Malafosse, A.; Karege, F. The Tutsi genocide and transgenerational transmission of maternal stress: Epigenetics and biology of the HPA axis. World J. Biol. Psychiatry 2014, 15, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J. Transgenerational Epigenetic Contributions to Stress Responses: Fact or Fiction? PLoS Biol. 2016, 14, e1002426. [Google Scholar] [CrossRef] [PubMed]
- Bale, T.L. Lifetime stress experience: Transgenerational epigenetics and germ cell programming. Dialogues Clin. Neurosci. 2014, 16, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Matthews, S.G.; Phillips, D.I. Minireview: Transgenerational inheritance of the stress response: A new frontier in stress research. Endocrinology 2010, 151, 7–13. [Google Scholar] [CrossRef]
- Tang, A.; Huang, Y.; Li, Z.; Wan, S.; Mou, L.; Yin, G.; Li, N.; Xie, J.; Xia, Y.; Li, X.; et al. Analysis of a four generation family reveals the widespread sequence-dependent maintenance of allelic DNA methylation in somatic and germ cells. Sci. Rep. 2016, 6, 19260. [Google Scholar] [CrossRef]
- Jenkins, T.G.; Carrell, D.T. The sperm epigenome and potential implications for the developing embryo. Reproduction 2012, 143, 727–734. [Google Scholar] [CrossRef]
- Yeshurun, S.; Hannan, A.J. Transgenerational epigenetic influences of paternal environmental exposures on brain function and predisposition to psychiatric disorders. Mol. Psychiatry 2019, 24, 536–548. [Google Scholar] [CrossRef]
- Krawetz, S.A.; Kruger, A.; Lalancette, C.; Tagett, R.; Anton, E.; Draghici, S.; Diamond, M.P. A survey of small RNAs in human sperm. Hum. Reprod. 2011, 26, 3401–3412. [Google Scholar] [CrossRef]
- Kretschmer, M.; Gapp, K. Deciphering the RNA universe in sperm in its role as a vertical information carrier. Environ. Epigenetics 2022, 8, dvac011. [Google Scholar] [CrossRef] [PubMed]
- Smeeth, D.; Beck, S.; Karam, E.G.; Pluess, M. The role of epigenetics in psychological resilience. Lancet Psychiatry 2021, 8, 620–629. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.T.; Printz, D.; Ching, T.; Wetterneck, C. Assessing PTSD in ethnic and racial minorities: Trauma and racial trauma. Dir. Psychiatry 2018, 38, 179–196. [Google Scholar]
- Zeitlin, S.B.; McNally, R.J. Implicit and explicit memory bias for threat in post-traumatic stress disorder. Behav. Res. Ther. 1991, 29, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Allsopp, K.; Read, J.; Corcoran, R.; Kinderman, P. Heterogeneity in psychiatric diagnostic classification. Psychiatry Res. 2019, 279, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Somoza, E.; Mossman, D. “Biological markers” and psychiatric diagnosis: Risk-benefit balancing using ROC analysis. Biol. Psychiatry 1991, 29, 811–826. [Google Scholar] [CrossRef]
- Sabeeha, S.; Hasnain, S.E. Forensic Epigenetic Analysis: The Path Ahead. Med. Princ. Pract. 2019, 28, 301–308. [Google Scholar] [CrossRef]
- Ramírez, C.G.; Queijeiro, A.V.; Morales, S.J.; López, D.B.; Miranda, A.H.; Chow, A.R.; Cárdenas, L.T.; Estrada, M.G. The NR3C1 gene expression is a potential surrogate biomarker for risk and diagnosis of posttraumatic stress disorder. Psychiatry Res. 2020, 284, 112797. [Google Scholar] [CrossRef]
- Wilker, S.; Vukojevic, V.; Schneider, A.; Pfeiffer, A.; Inerle, S.; Pauly, M.; Elbert, T.; Papassotiropoulos, A.; de Quervain, D.; Kolassa, I.T. Epigenetics of traumatic stress: The association of NR3C1 methylation and posttraumatic stress disorder symptom changes in response to narrative exposure therapy. Transl. Psychiatry 2023, 13, 14. [Google Scholar] [CrossRef]
- Cao-Lei, L.; Saumier, D.; Fortin, J.; Brunet, A. A narrative review of the epigenetics of post-traumatic stress disorder and post-traumatic stress disorder treatment. Front. Psychiatry 2022, 13, 857087. [Google Scholar] [CrossRef]
- Yehuda, R.; Daskalakis, N.P.; Desarnaud, F.; Makotkine, I.; Lehrner, A.L.; Koch, E.; Flory, J.D.; Buxbaum, J.D.; Meaney, M.J.; Bierer, L.M. Epigenetic biomarkers as predictors and correlates of symptom improvement following psychotherapy in combat veterans with PTSD. Front. Psychiatry 2013, 4, 118. [Google Scholar] [CrossRef] [PubMed]
- Church, D.; Yount, G.; Rachlin, K.; Fox, L.; Nelms, J. Epigenetic Effects of PTSD Remediation in Veterans Using Clinical Emotional Freedom Techniques: A Randomized Controlled Pilot Study. Am. J. Health Promot. 2018, 32, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Bishop, J.R.; Lee, A.M.; Mills, L.J.; Thuras, P.D.; Eum, S.; Clancy, D.; Erbes, C.R.; Polusny, M.A.; Lamberty, G.J.; Lim, K.O. Methylation of FKBP5 and SLC6A4 in Relation to Treatment Response to Mindfulness Based Stress Reduction for Posttraumatic Stress Disorder. Front. Psychiatry 2018, 9, 418. [Google Scholar] [CrossRef] [PubMed]
- Carleial, S.; Natt, D.; Unternahrer, E.; Elbert, T.; Robjant, K.; Wilker, S.; Vukojevic, V.; Kolassa, I.T.; Zeller, A.C.; Koebach, A. DNA methylation changes following narrative exposure therapy in a randomized controlled trial with female former child soldiers. Sci. Rep. 2021, 11, 18493. [Google Scholar] [CrossRef]
- Xulu, K.R.; Womersley, J.S.; Sommer, J.; Hinsberger, M.; Elbert, T.; Weierstall, R.; Kaminer, D.; Malan-Muller, S.; Seedat, S.; Hemmings, S.M.J. DNA methylation and psychotherapy response in trauma-exposed men with appetitive aggression. Psychiatry Res. 2021, 295, 113608. [Google Scholar] [CrossRef]
- Pape, J.C.; Carrillo-Roa, T.; Rothbaum, B.O.; Nemeroff, C.B.; Czamara, D.; Zannas, A.S.; Iosifescu, D.; Mathew, S.J.; Neylan, T.C.; Mayberg, H.S.; et al. DNA methylation levels are associated with CRF(1) receptor antagonist treatment outcome in women with post-traumatic stress disorder. Clin. Epigenetics 2018, 10, 136. [Google Scholar] [CrossRef]
- Carver, R.B.; Castera, J.; Gericke, N.; Evangelista, N.A.; El-Hani, C.N. Young Adults’ Belief in Genetic Determinism, and Knowledge and Attitudes towards Modern Genetics and Genomics: The PUGGS Questionnaire. PLoS ONE 2017, 12, e0169808. [Google Scholar] [CrossRef]

{kind=link}
| Gene | Gene Function | Modification | Modification Site | Reference |
|---|---|---|---|---|
| ACP5 | glycoprotein | Methylation | - | [27] |
| AHRR | xenobiotic metabolism | Methylation | - | [30] |
| AHRR | xenobiotic metabolism | Demethylation | cg05575921 and cg26703534 | [33] |
| ANXA2 | signal transduction and cellular growth | Methylation | - | [27] |
| APBA1/2 | protein transportation and synaptic function in brain tissue | Methylation | - | [29] |
| BDNF | neuroplasticity | Methylation | - | [27] |
| BRSK1 | tumor suppressor | Methylation | - | [27] |
| CCDC88C | coiled-coil domain containing 88C | Methylation | [25] | |
| CHST11 | extracellular signaling and neuronal plasticity in brain tissue | Methylation | - | [29] |
| CLEC9A | myeloid cells activation | Methylation | - | [27] |
| CXCL1 | chemoattractant of immune cells interacting with BDNF to regulate neuroplasticity | Methylation | - | [27] |
| DOCK2 | immune cells chemotaxis | Methylation | - | [27] |
| FKBP5 | co-chaperone inhibiting glucocorticoid receptor | Methylation | cg07485685 | [25,27,28] |
| G0S2 | lipid metabolism, downregulated in response to stress | Methylation | - | [29] |
| HGS | lysosome-dependent degradation | Methylation | - | [28] |
| HEXDC | enables beta-N-acetylhexosaminidase activity | Methylation | [31] | |
| LCN8 | ligand transportation | Methylation | - | [27] |
| MAD1L1 | component of the mitotic spindle-assembly checkpoint | Methylation | [31] | |
| MAN2C1 | apoptosis regulation | Methylation | - | [27] |
| NCR3C1 | glucocorticoid receptor | Methylation | cg19645279 | [25,34,35] |
| NRG1 | cell–cell signaling | Methylation | - | [28] |
| RNF39 | synaptic plasticity | Methylation | - | [28] |
| SPRY4 | associated with suicidal behavior | Long non-coding RNA | - | [29] |
| TLR8 | pathogen recognition | Methylation | - | [27] |
| TPR | cellular trafficking | Methylation | - | [27] |
| ZFP57 | transcriptional repressor | Methylation | - | [28,36] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chou, P.-C.; Huang, Y.-C.; Yu, S. Mechanisms of Epigenetic Inheritance in Post-Traumatic Stress Disorder. Life 2024, 14, 98. https://doi.org/10.3390/life14010098
Chou P-C, Huang Y-C, Yu S. Mechanisms of Epigenetic Inheritance in Post-Traumatic Stress Disorder. Life. 2024; 14(1):98. https://doi.org/10.3390/life14010098
Chicago/Turabian StyleChou, Pei-Chen, Yu-Chi Huang, and Sebastian Yu. 2024. "Mechanisms of Epigenetic Inheritance in Post-Traumatic Stress Disorder" Life 14, no. 1: 98. https://doi.org/10.3390/life14010098
APA StyleChou, P.-C., Huang, Y.-C., & Yu, S. (2024). Mechanisms of Epigenetic Inheritance in Post-Traumatic Stress Disorder. Life, 14(1), 98. https://doi.org/10.3390/life14010098