Abstract
The skin’s recognised functions may undergo physiological alterations due to ageing, manifesting as varying degrees of facial wrinkles, diminished tautness, density, and volume. Additionally, these functions can be disrupted (patho)physiologically through various physical and chemical injuries, including surgical trauma, accidents, or chronic conditions like ulcers associated with diabetes mellitus, venous insufficiency, or obesity. Advancements in therapeutic interventions that boost the skin’s innate regenerative abilities could significantly enhance patient care protocols. The application of Platelet-Rich Plasma (PRP) is widely recognized for its aesthetic and functional benefits to the skin. Yet, the endorsement of PRP’s advantages often borders on the dogmatic, with its efficacy commonly ascribed solely to the activation of fibroblasts by the factors contained within platelet granules. PRP therapy is a cornerstone of regenerative medicine which involves the autologous delivery of conditioned plasma enriched by platelets. This is achieved by centrifugation, removing erythrocytes while retaining platelets and their granules. Despite its widespread use, the precise sequences of cellular activation, the specific cellular players, and the molecular machinery that drive PRP-facilitated healing are still enigmatic. There is still a paucity of definitive and robust studies elucidating these mechanisms. In recent years, telocytes (TCs)—a unique dermal cell population—have shown promising potential for tissue regeneration in various organs, including the dermis. TCs’ participation in neo-angiogenesis, akin to that attributed to PRP, and their role in tissue remodelling and repair processes within the interstitia of several organs (including the dermis), offer intriguing insights. Their potential to contribute to, or possibly orchestrate, the skin regeneration process following PRP treatment has elicited considerable interest. Therefore, pursuing a comprehensive understanding of the cellular and molecular mechanisms at work, particularly those involving TCs, their temporal involvement in structural recovery following injury, and the interconnected biological events in skin wound healing and regeneration represents a compelling field of study.
1. Skin Functionality, Tissue Aggression, and Soft Tissue Lesions
The skin, the body’s largest organ, is composed of three primary layers: the epidermis, the dermis, and the subcutaneous tissue [1,2]. The epidermis—the outermost layer—acts as a protective waterproof barrier and also contributes to skin colour. The epidermis of thick skin areas (the palms and soles) consists of five sublayers: the stratum basale, stratum spinosum, stratum granulosum, stratum lucidum, and stratum corneum. The epidermis elsewhere on the body typically consists of four layers, lacking the stratum lucidum. Beneath the epidermis, the dermis consists of connective tissue and hosts a variety of structures including hair follicles, blood vessels, lymphatic vessels, and sweat glands. The dermis itself is divided into two distinct layers: the upper papillary dermis and the lower reticular dermis. The subcutaneous tissue (hypodermis, or fascia subcutanea) is the deepest layer of the skin, primarily made up of fat and connective tissue. This layer plays a crucial role in the thermal and mechanical insulation of the body, thus protecting underlying muscles and other internal structures.
However, the skin is a versatile organ, essential for maintaining overall health and well-being. It serves as a protective barrier against external threats such as microorganisms, dehydration, ultraviolet light, and physical injury, thus acting as the body’s first line of defence. Its cutaneous sensory functions, including the perception of pain, temperature, touch, and deep pressure, enable the body to respond to external stimuli. The skin also supports mobility, facilitating smooth movement and accommodating various degrees of stretching and bending. Additionally, it plays a key role in endocrine functions, notably in synthesizing vitamin D, essential for calcium absorption and normal bone metabolism. The skin’s exocrine functions involve releasing substances like water, urea, and ammonia and producing sebum, sweat, and pheromones. It also has immunologic roles, releasing bioactive substances like cytokines, and contributes to the body’s immunity against various pathogens. Finally, the skin is vital in thermoregulation, helping to either conserve or release heat as needed, and it plays a crucial role in maintaining the body’s water balance and overall homeostasis. The skin wound healing process is a remarkable interplay of cellular events and signalling molecules that participate in tissue remodelling. A profound understanding of this process could have significant implications for clinical medicine, particularly in those conditions that feature impaired healing. Depending on the body segment involved, the severity and the extent of the involved area, soft tissue healing can be a long, a slow, and sometimes incomplete (but constantly complicated) process, mainly regulated by the secretion and concentrations of the local growth factors (GFs). The development of new treatments such as PRP injections can enhance the body’s natural repair mechanisms and eventually help in speeding up wound healing.
Skin wound healing involves a series of intricate steps and sequences of multi-cellular interplays, in which secreted cytokines and chemokines ensure efficient tissue repair and regeneration (Figure 1). Usually, the wound healing process falls into four consecutive phases [3].
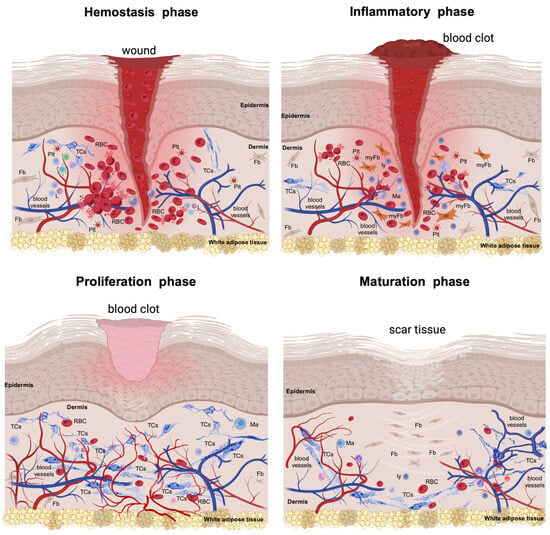
Figure 1.
The injury of the skin triggers a series of physiologic and morphological events for repairing the injured structures. This cascade of healing falls into four overlapping phases. The hemostasis phase (primarily about coagulation) is acute and begins with injury, followed by the formation of the blood clot that dams the bloodshed; the platelets become activated by their contact with basement membranes and collagen fibres and begin aggreging. Thrombin initiates the formation of the fibrin mesh. The inflammatory phase (which is mainly about removing the cellular debris) is orchestrated by neutrophils and macrophages and their variegated secretory panel. Clinically, it lasts about 6 days and the Celsus cardinal signs feature this phase. The proliferative phase (which is about filling and covering the skin defect) structurally features the formation of the granular tissue and neoangiogenesis and wound covering by the neighbouring epithelial cells. All these intricate processes can last about 3 weeks. In both the inflammatory and proliferative phases, the formerly destroyed network of interstitial cells (including telocytes, TCs) is restored to its initial parameters. The maturation phase (dominated by structural remodelling and reorganization processes) is the longest-lasting and is dominated by the rearrangement and maturation of the collagens, increasing the tensile strength. Thus, the cellular architects of the phase are the fibroblasts (Fbs) and fibrocytes. RBC—red blood cell; Plt—platelets; L—leukocytes; myFb—myofibroblast; ly—lymphocytes. The image was created with BioRender.com.
1.1. The Haemostasis Phase
This phase comes briefly after injury and is an immediate response that stops bleeding by constricting the damaged blood vessels. This process is swiftly followed by platelet activation, leading to the formation of a fibrin clot [4]. Platelets are activated, thus adhering to exposed collagen and other matrix elements, leading to the aggregation and formation of the platelet plug, stabilised by fibrin strands [5]. This clot serves a dual purpose: it halts blood loss and creates a structural framework for the next arriving inflammatory cells.
1.2. The Inflammatory Phase
This phase comes next and is marked by cell migration (mainly of neutrophils and macrophages). Neutrophils are the first responders, chemotactically attracted to the wound, and warding off any potential bacterial infections (mainly by phagocytosing debris and bacteria) [6]. Between 48 to 96 h post-injury, in the wound site the dispatched monocytes are differentiating into tissue-activated macrophages [7]. These cells release cytokines that further modulate the inflammatory response [8]. Additionally, the innate immune response of the adaptive immune system of the body comes into play. The key cellular players from this system (e.g., Langerhans cells, dermal dendritic cells, and T cells) are triggered into action to tackle both self- and foreign antigens present at the wound site. Recently, there has been keen scientific interest in discerning which exact cellular subset(s) is majorly dominating the cellular debris removal wound scene, or which cells are more focused on quelling infections [9]. As the inflammatory phase wanes, neo-angiogenesis takes centre stage. This sequence is marked by the proliferation and migration of endothelial cells, forming new blood vessels. Simultaneously, local pericytes provide the scaffolds and structural integrity for the burgeoning endothelial cells [10,11]. Current studies are attributing their enhanced adaptability to pericytes, considering them mesenchymal stromal cells [12]. Moreover, the formation of new blood vessels is also attributed to progenitor cells from the bone marrow [13,14,15].
1.3. The Proliferative Phase
This is marked by fibroblasts’ migration and the deposition of the extracellular matrix (ECM), promoting a developing cellular microenvironment. The epithelialization process permits keratinocytes to proliferate and migrate over the wound base [16]. In this stage, the dedifferentiation into the interfollicular epithelium of the Gata6+ cells originating from the sebaceous duct allows them to acquire the self-renewal capacity and further differentiate, more than they normally would, into numerous cell lineages [17]. An important aspect of the epidermal layer restoration is the regeneration of skin appendages [11,18]. It is worth noting that these epidermal stem cells predominantly display unipotency under regular conditions. This episodic plasticity enables them to differentiate into a variety of cell types, facilitating epidermal repair.
The local native fibroblast population is thriving in this phase, and they are being transferred to the newly formed clot, thus participating in the formation of the contractile granulation tissue. Certain fibroblasts undergo differentiation into myofibroblasts, playing a pivotal role in contracting the wound edges [19,20].
1.4. The Maturation Phase
This final phase is dominated by the remodelling, rearrangement, cross-linking, and maturation of the collagen fibres, increasing the collagen tensile strength, and strengthening the scar’s physical resistance. The wound continues to contract as the myofibroblasts remain active. Usually, unemployed cells, including fibroblasts, myofibroblasts, and inflammatory cells, undergo programmed cell death [21].
A compromised wound healing process features the evolution of chronic wounds [22]. Such wounds are predominantly seen in individuals with diabetes, vascular disorders, hemoglobinopathies, neuropathies, and in the elderly. This wound mismanaging is usually marked by recurrences, or can even require dramatic solutions (e.g., limb amputation), while severe cases may prove fatal. Metabolic diseases (e.g., obesity or type 2 diabetes), are marked by inflammatory cells within the granular tissue layer gaining immunomodulatory significance, which can further modify the wound healing course [23]. However, in different scenarios, the healing process efficiently restores the skin’s barrier function and nearly reinstates its original tensile strength. However, there is a distinction between prenatal and adult wound healing. While prenatal wound healing is mainly regenerative (mirroring the original structure of the skin), the latter often culminates in a fibrous scar, acting as a swift fix for the wound [24]. Exuberant scarring and collagen overproduction can tip the balance and frequently causing hypertrophic scars, or even keloids [25]. Emerging research suggests that scarring outcomes might be influenced by varied cellular reactions to the mechanical stress experienced by the healing skin [26,27].
2. Platelet-Rich Plasma (PRP): Its Introduction in Tissue Repair
Lesion progression in skin ageing and in senescent skin transformation appears due to either endogenous/(epi)genetic factors and/or exogenous factors, all of these inducing progressive accumulations of (ultra)structural skin defects secondary to their action [28]. Since photoaging impedes the normal biosynthesis of collagen and elastic fibres within a spectrum of the also-generated dermal inflammatory context, the UV radiation increases the metalloproteinases’ (MMPs) activity due to dermal extracellular matrix degradation (this including the drop-off in collagen production) [29].
The usage of Platelet-Rich Plasma (PRP) is a regenerative medicine procedure that stimulates the healing response of the tissue (after injury) using products from the blood of the patient, collected after centrifugation of a whole blood sample. Apparently, PRP is the simplest, least invasive, and most effective procedure that locally supplements the GF through an autologous transplant of GFs, thus boosting the healing processes. The PRP products have heterogeneous biological effects and a regenerative capacity, known as orthobiologics when used, for example, in treating musculoskeletal diseases [30]. The PRP targets different cells and tissue environments, modulating cell signalling and/or having a paracrine effect [31]. The main advantages of autologous PRP are, with the proper ratio of GF, the lack of immunological problems and transmitting diseases, and its ability to form a gel when injected. This low viscosity gel state is acquired within tissue, permitting platelets to locally deliver and to maintaining high concentrations of GF [28]. The anti-inflammatory effect of PRP seems to be induced by the suppression of cyclooxygenase and production of prostaglandins and by increasing the intracellular expression of inflammation-controlling cytokines [32].
PRP has been applied in human clinical settings since the 1970s because of its therapeutic properties due to the autologous GF and secretory proteins it contains and their great potential to amplify the cellular healing processes [33]. Moreover, PRP promotes the attraction, multiplication, and transformation of those cells essential for tissue repair/regeneration [34]. Products associated with PRP are sometimes referred to as a platelet-rich concentrate, platelet gel, GF-rich preparations, and platelet release, and have been previously examined into in vitro and in vivo, as add-on procedures to surgical interventions [35]. This diversity of PRP products should be considered when analysing and comparing the results of PRP transplants. Usually, these differences of various amplitudes are in accordance with the processing and leveraging of PRP products for their usage [36].
The composition of PRP can vary based on the equipment and methodology employed, leading to different levels of plasma, erythrocytes, white blood cells, and platelets in the final product. For a sample to be truly designated as “platelet-rich”, its platelet count should exceed the baseline concentration in whole blood by, as is commonly accepted, at least five-fold [37].
The PRP preparation obtained by centrifugation of the whole blood contains more than 1100 different proteins (including GFs), many of which have already documented functions in various biological processes (including regeneration/remodelling processes, or angiogenesis) [28]. PRP that boasts a platelet count of a minimum of 1,000,000 platelets/μL in a 5 mL plasma sample has been linked to improved healing [38]. PRP’s potential to accelerate healing arises from its delivery of a diverse array of GFs and cytokines, found in the α–granules within platelets. Notably, PRP exhibits an eight-fold surge in GF levels compared to whole blood [39]. Platelets degranulate within the first 10 min of participation in the clotting cascade or on contact with the basement membrane [36].
After the transplantation of PRP, the myriad of proteins and GFs either boost some normal homeostatic processes or create local premises and further scaffolds for the reparatory processes/remodelling processes [28]. However, previous observations have indicated that the efficient and short epithelialization of skin wounds is proportional to the density and synthetic activity of dermal fibroblasts [32].
In vitro studies have revealed a direct correlation between platelet concentrations and the proliferation of human mesenchymal stem cells, fibroblast growth, and the synthesis of type I collagen, thus amplifying the wound healing process [40]. Experimental animal studies have documented, mainly clinically, its efficacy in both soft and hard tissue repair [41,42].
Currently, PRP is successfully used, with remarkable results (especially in bone renewal, or soft tissue maturation) in orthopaedics, periodontics, maxillofacial surgery, urology, and plastic surgery. Yet, the medical community has different opinions regarding its efficacy, since there are studies indicating substantial upticks in bone development and maturation [35], while others found no discernible enhancements [43].
3. Potential Mechanisms Underlying PRP’s Effects
PRP is recognized for its youth-preserving effects in dermatology and skincare. Although the precise process by which PRP works is not fully understood, it is known that PRP injections trigger a cascade of cellular activities in aged skin. Compounds like cellulose, fibronectin, and vitronectin within PRP form complexes with growth factors at the site of injection [44]. These compounds may provide a framework to support the growth of new cells and tissues, thus aiding in the rejuvenation of aged skin through the stimulation of DNA synthesis and gene expression at the cellular level [28,45]. The application of PRP to aged skin has been shown to thicken the reticular dermis due to increased collagen and elastin fibre deposition. Additionally, laboratory studies suggest that PRP can mitigate skin damage from UV exposure by reducing MMP-1 and tyrosinase levels while boosting the production of tropoelastin and fibrillin [28].
It is recognized that, within PRP, the platelets are dormant until administered to a damaged area. Here, interaction with the tissue factor initiates a transformation in platelet form, leading to the formation of extensions known as pseudopods, which facilitate the clustering of platelets [46]. These activated platelets then secrete growth factors such as PDGF, VEGF, TGFβ, and EGF at the injury site, which contribute to a cascade of biological activities that stimulate new blood vessel formation, tissue and bone repair, cell multiplication, and overall healing [47].
PDGF is a type of glycoprotein released when platelets degranulate at an injury site. It binds to specific receptors on the membranes of target cells, leading to the creation of high-energy phosphate bonds. These bonds then activate signalling proteins that trigger the target cells’ specific functions such as cell division, the formation of new blood vessels, and the activation of macrophages [47]. TGFβ, another compound released by platelets and macrophages, serves as an inhibitor of cell multiplication in normal epithelial cells and functions both through paracrine and autocrine mechanisms [48]. Its primary target cells include fibroblasts, marrow stem cells, and pre-osteoblasts, which are integral to long-term healing, bone restoration, and skeletal remodelling. TGFβ also plays a role in preventing the formation of osteoclasts. VEGF, which was first identified as a factor that makes blood vessels more permeable, is a signalling protein that encourages the formation of new blood vessels and the development of vascular systems [49]. Lastly, EGF is a growth factor that promotes cellular growth, proliferation, and differentiation by interacting with its receptor, EGFR [50].
Platelets are small cellular fragments circulating in the blood which have a relatively short lifespan. A normal body produces around 100 billion new platelets daily [51]. In comparison, various cell types have much longer lifespans; fibroblasts can survive up to 270 days in lab conditions [52], osteoblasts range from 1 to 200 days, bone lining cells can last between 1 to 10 years, and osteocytes may endure for as long as 50 years [53]. These longer-lived bone cells may accumulate substantial cellular damage over time, contributing to senescence and the Senescence-Associated Secretory Phenotype. Conversely, the frequent renewal of platelets, occurring every 7–10 days, suggests they are less prone to the effects of senescence. The secretion of growth factors and cytokines from platelet granules is known to expedite tissue healing, a process that is augmented when interacting with the fibrinolytic system [54]. This could explain one of the pathways through which PRP exerts its regenerative and potentially anti-ageing effects.
4. Thrombocyte’s Genesis and Phenotype
Platelets represent fragments of the cytoplasm and cell membrane of the thrombocytogenic megakaryoblast. Platelets are the first blood figurative elements that arrive at the tissue lesion site, and are involved in the initial inflammatory phase of the healing process through their adherence, aggregation, participation in clot formation, and promotion of angiogenesis and tissue repair/regeneration by releasing proteins and growth factors [36]. Platelets produce and locally deliver antibacterial proteins, metalloproteases, coagulation factors, and membrane glycoproteins [55]. The α–granules from the platelets harbour diverse GFs, among them the platelet-derived GF (PDGF), transforming GF β1 (TGF-β 1), vascular endothelial GF (VEGF), basic fibroblast GF (βFGF), and epidermal GF (EGF). The main body of granular GFs is mainly released within the first hour, but their release continues for the next 7 days after their activation at the clot site. The dense granules of platelets contain ADP, ATP, calcium ions, histamine, serotonin, and dopamine, which are active in tissue modulation and regeneration [56].
4.1. Development of Platelets
The development the hematopoietic stem cells is successively located in the yolk sac, liver, and spleen. During the whole process, stem cells lose their self-renewal capacity, and they transform into short-term self-renewal cells and pluripotent progenitor cells that will initiate lymphoid and myeloid haematopoiesis. Pluripotent hematopoietic progenitors found within the proximal osteoblastic niche of the bone, due to endomitosis after multiple DNA replications (but no cytokinesis), generate the committed megakaryocytes (MKs). Their differentiation into MKs is regulated by interleukins (IL-3, Il-6, IL-11) or the stem cell factor (SCF). At the level of bone marrow, MKs are in close vicinity to bone marrow sinusoids. MKs begin the rapid development of a very dense invaginated membrane system [57] and the agglomeration of proteins either in the cytoplasm or in the granules that will become essential for further functionality of the platelets—the α– and δ–granules [58]. The hematopoietic progenitor cells are positive for CD34 and CD41. However, an increased expression of CD41, along with CD42 and CD61, indicates a commitment to the MKs’ lineage [59,60].
On the other hand, switching off the chemotactic CXCR4 signalling pathway in the development of MKs permits MKs to leave the bone marrow, and to enter mainstream circulation [61]. Thrombocytogenic MKs represent less than 0.1% of the cells in bone marrow cell populations, but this number can be altered by chemokines, thrombopoietin, and other ligands of MKs receptors [62].
However, it is known that intact MKs can cross the sinusoids walls, enter the blood stream, and create blockages at the level of the pulmonary microcirculatory sieve (another acknowledged site of platelet production) [63]. Since more than 100 years ago, when MKs were identified in the pulmonary circulation as having migrated from the bone marrow into the bloodstream and having hitchhiked into the lung circulation, the production of platelets within the lungs has been documented (especially during foetal development) [64], even though the lungs are not the primary sites of platelets’ synthesis. It was previously acknowledged that 25 × 104 MKs are getting into lung tissue, hourly [60]. Already published morphometry data documented a presence of 14–65 MKs/cm2 in the lung (which were denser in the central and right upper lobes) [65]. However, MKs are almost 10 times more concentrated in the pulmonary artery than the aorta [66].
The development of MKs can be divided into three stages: the pro-megakaryoblast (first recognizable precursor), the megakaryoblast, and the promegakaryocyte (which, by endomitotic cell divisions, undergo DNA replication without cytokinesis, becoming the polyploid MK) [67]. Previously published studies have indicated a finite number of the endomitotic divisions of the mononuclear MKs (usually between 2–6 endomitotic cycles). Thus, many definitive MKs have a 16n of DNA content, thus indicating that the third endomitotic cycle is the most frequent [68,69,70]. The evaluation of the biological significance of polyploidisation indicated that 16n MKs will produce approximately 2000 platelets, 1 × 103 more platelets than a 2n MK; this ontogenic advantage creates more efficient platelet production [71]. MKs are producing platelets within the bone marrow, but they can migrate and produce platelets in the bloodstream and could even initiate platelet biogenesis in other tissues (e.g., the lungs) [72].
After endomitosis, the MKs’ cytoplasm will definitively mature by accumulating platelet-specific proteins and developing a prominent organelle compartment and an elaborate membrane system. This invaginated membrane system was proposed as the origin of MKs pseudopodia—the proplatelets [73]. Supplementarily, a sign of MKs’ maturation (including the maturation of the MKs’ cytoplasm) is the development of an extensive three-dimensional network of flattened cisternae and tubules of different lengths and calibres, delimiting the so-called “platelet territories”. These will serve as a membrane reserve for the additional proplatelets [74]. It is easy to understand that this invaginated membrane system represents the origin of the later mature platelets, of the open canalicular system—a network of intra-platelet surface-connected membrane channels along the dense tubule system [75].
Another essential change for platelet formation is the structural, ultrastructural, and biochemical changes within MKs, including the dimerizing α– and β– 1 tubulin and their polymerisation into microtubules organised in bundles with a cortical arrangement within MKs. The bundles of microtubules are dictating the length of the proplatelets, either through sliding antiparallel microtubules (by cytoplasmic dynein), or by the localised polymerization of microtubules within the proplatelets [76]. Moreover, the subsequent proplatelet branching is also dependent upon the actin filaments’ rearrangements (as actin is a MKs cytoskeleton component). Naturally, any defects of the cytoskeletal actin dynamics are also associated with impaired platelet production in mammals (including humans) [77].
MKs proplatelets protrude through bone marrow sinusoids walls and release fragments—the pre-platelets (that are usually larger than typical platelets)—within their lumens. The pre-platelets are larger discoid cellular fragments that, via forces applied across the microtubular system, are converted into barbell-shaped platelets, or can fragment directly into platelets [60]. However, the final stages of the maturation and recovery of the platelets occur outside the bone marrow microenvironment, but exclusively within the microcirculation [78]. The proplatelets can be separated into individual platelets by centrifugation of the blood, due to applied gravitational/shear forces [79].
The progression of platelets’ formation and maturation are marked by a series of changes in the organelles’ presence and distribution within platelets, perhaps due to cytoskeletal changes. Mitochondria are (almost) twice as numerous in platelet intermediates than resting platelets, mainly representing an energetic substrate for protein synthesis in the intermediate stages of platelet development [80]. Also, dense (δ–) and α–granules are scant within the cytoplasmic bridges of barbell-shaped proplatelets [81]. Published data have documented the different intermediate stages in platelet maturation, including the resting platelets, circular pre-platelets, oval-shaped pre-platelets, barbell-shaped pre-platelets, multibody pro-platelets [79].
Since platelets are only cellular fragments of MKs’ cytoplasm enwrapped in fragments of the MKs membrane, they have no nucleus, but only scarce mRNA and a few related organelles involved in protein production (endoplasmic reticulum/dense tubular system (DTS), Golgi apparatus, mitochondria, autophagosomes, or endosomes) [82,83,84,85]. Platelets maintain the protein synthesis ability (e.g., P-selectin, actin, etc.), or can incorporate proteins from the plasma or other sources, storing all these mainly into the α–granules [86,87]. Usually, the protein content of the platelets can be modulated by protein post-translational modification and is also regulated by different rates of protein degradation (an important process for platelet function) either by the ubiquitin-proteasome system or the autophagy-lysosome system [88,89]. The total number of proteins within platelets is known as the platelet proteome (mainly produced in MKs and transferred to platelets), and this proteome contains approximately 4000–5000 distinct proteins [90,91,92].
There are few accepted mechanisms of platelet production, specifically:
- Platelet budding from the megakaryocyte surface (apparently, these membrane blebs do not contain organelles) [93];
- Proplatelet formation, elongation, and branching is the process by which MKs form thin cellular prolongations with a moniliform aspect, typically presenting an alternation of dilated platelet-sized segments bounded by thin cytoplasmic bridges. The rupture of these cytoplasmic bridges releases fragments of the MKs’ cytoplasm (enwrapped in MK cell membrane) into circulation, also known as platelets [94];
- MKs cytoplasm fragmentation through the invagination membrane system (this functions as a reserve that is providing membranes for the proplatelets’ development) [95].
The in vitro production of platelets prompts reprogramming pluripotent stem cells to obtain a new expression phenotype for a few reprogramming transcription factors (Klf4, Sox2, Oct3/4, and c-Myc) [96]. It was shown that applying CD34-positive human cord blood cells to hydrogel scaffolds coated with fibronectin and thrombopoietin increases human platelet production [97].
Cultured MKs, in the presence of thrombopoietin, underwent a precise sequence of subcellular changes that finally lead to the pro-platelets’ production: a. the alignment/dilation of the demarcation membranes; b. the unfolding of cytoplasmic sheets and extensions of cell processes; c. the delimitation of pro-platelet-like territories; d. the appearance of the irregular constriction points and longitudinal axis of microtubules; e. the appearance and development of a central vacuole within the constriction zone centre; and f. platelet formation. Apparently, unlike thrombin-activated cultivated MKs, tissue MKs are surrounded by numerous shed microparticles of 100–300 nm which are strongly expressing GpIIb-IIIa, this fact supports the theory of cross-sectioned neighbouring cells processes [63,68,98,99]. MKs’ proplatelet-like cellular prolongations could be influenced by either by α–/β–tubulin polymerization inhibitors, or by low temperature [100]. Phase-contrast microscopy showed that within a single proplatelet cellular process of MK, when a more proximal segment is budding, several-times-larger platelets (about 10 mm) are obtained (in comparison with distal segment budding) [101].
Circulating platelets have a lifespan of about 8–10 days in the bloodstream. They are involved in regulating innate immunity and neo-angiogenesis. To maintain a constant count of platelets, almost 1 × 1011 of platelets are produced daily. They respond to blood vessel lesions by modifying their appearance, excreting their granular content, and participating in blood clot formation [62].
4.2. The Platelet’s Structure and Ultrastructure
Platelets are abundantly present within peripheral blood circulation and are traditionally responsible for maintaining the equilibrium between thrombosis and haemostasis. There are two functional states of normal platelets: the non-activated (resting) platelets, which maintain their discoid shape, and the activated platelets, which have morphological changes, mainly mediated by their cytoskeleton components, and which are ultrastructurally more amorphous [102,103,104]. In light microscopy, platelets demonstrate a discoid shape with dimensions 2–3 μm, but the diameter and the shape of the platelets are variegated. They usually contain α–granules, δ–granules, an open canalicular system, and microtubules, usually located on a side of the platelets, or transversely (corresponding to the constriction points of the proplatelets) [105]. The location of the microtubules (shaped like a coil) close to the constriction points could support the idea of the increased local rigidity of the membrane with implications in the clefting progression and further detaching of the platelets [106,107]. As cellular fragments, platelets are naturally marked by a plasma membrane enclosing a small aliquot of cytoplasm (Figure 2). Ultrastructurally, the plasma membrane is harbouring the apertures of an open canalicular system [108].
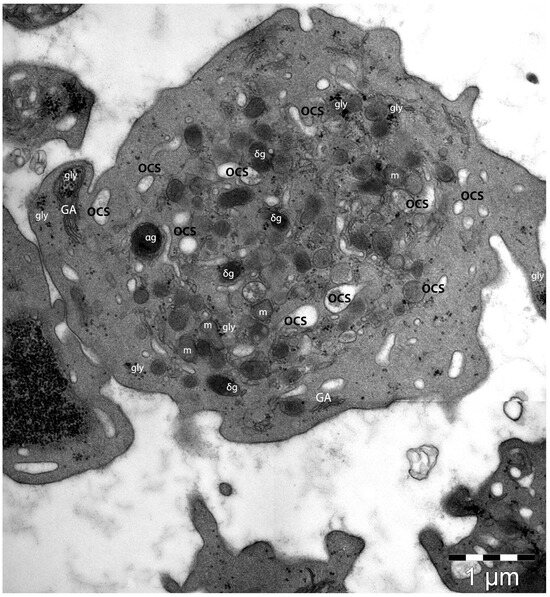
Figure 2.
Transmission electron microscopy image of a platelet. Under the plasma membrane normal platelet specific organelles are present. The entire volume presents scarce α–granules (αg) and δ-granules (δg) among several mitochondria (m) and multiple granules of glycogen (gly). The elements of the open canalicular system (OCS) indicate its three-dimensional distribution within the entire platelet volume. A scant distribution of Golgi apparatus (GA) elements is also visible. (Courtesy of Dr. E.T. Fertig, “Victor Babeş” National Institute of Pathology, Bucharest, Romania).
Biochemically, the platelet’s plasma membrane has the same composition as the MKs’ plasma membrane. Calcium release and the plasma membrane proteins participate in maintaining sphingolipids’ asymmetry with implications of their roles in thrombin generation by providing an optimum membrane surface for coagulation cascade complexes [109,110,111]. This process is involved in the coagulation cascade. The lipidomics of the platelet’s plasma membrane indicated the presence of the phospholipids (phosphatidylserine, phosphatidylethanolamine, phosphatidylcholine), sphingomyelin, and cholesterol, with this composition being crucial for the normal function of the platelets, with implications in their activation, degranulation, and exocytosis [108]. Previously published data has indicated that particular changes in lipid fractions at the level of the platelets’ plasma membrane have affected the platelets’ functioning in several studied pathologies (hypertension, lung cancer, or alcoholic liver disease) [112], or drug-related activities (aspirin, ticagrelor, or few statins intake) [113,114].
The submembranous region is one of the most important ultrastructural elements for proper platelet function, since at this level the intracellular domains of the transcellular receptor proteins interact with the proteins in platelets’ signalling cascades [115,116].
The cytoskeleton of the resting platelets is made of actin, tubulin, and spectrin; all these three conjugated proteins provide shape and support for the platelet’s morphology within circulation. This cytoskeletal architecture also permits a rapid response of the platelets to vascular damage [79]. The membrane skeleton-associated spectrin is connected to actin filaments via filamin, thus is creating a continuous ultrastructural element that promotes similar sizes for the forming platelets [117].
Microtubules are polymers of α– and β–tubulin subunits and they are dynamic and polarised structures, circumferentially organised in bundles of about 20 microtubules (Figure 3). The microtubules are located below the spectrin membrane skeleton. This microtubular loop plays an important role in realising and maintaining the shape and size of the resting platelets [118]. In early platelets, the microtubules are organised longitudinally, favouring their elongated shape and increased flexibility, differently from older round platelets, whose appearance is marked by circumferentially organised bundles of microtubules [100,119].
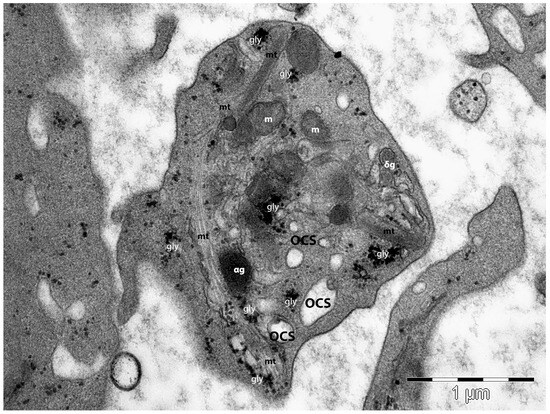
Figure 3.
Transmission electron microscopy image of a platelet. The peripheral distribution of the microtubular (mt) loop is obvious and demonstrates its implication in platelet morphology and shape maintenance. Three-dimensionally open canalicular system (OCS) elements are present, along with glycogen granules (gly), mitochondria (m), α–granules (αg), and δ-granules (δg). (Courtesy of Dr. E.T. Fertig, “Victor Babeş” National Institute of Pathology, Bucharest, Romania).
The actin filaments are organised in a meshwork and are also dynamic and polarised fibrillar structures, whose assembly is regulated by many actin-binding proteins. The constituted platelet actin cytoskeleton provides structural support for the platelets, particularly when they are subjected to shearing forces in circulation [120,121,122]
Elements of the cytoskeleton are involved in the regulation of the platelet’s granule secretion, since, in the platelet resting state, the actin functions as a barrier to secretion but supports the granule sorting and exocytosis in the activated platelets [123,124,125,126,127].
The surface-connected canalicular system represents a three-dimensional labyrinthine system of randomly organised platelet-internal canaliculi, which originates from the plasma membrane [62,75,128,129,130,131,132,133]. The origin of this open canalicular system is related to the demarcation membrane system of MKs, and its biochemical structure is like that of the plasma membrane [134,135,136]
Below the plasma membrane, the platelets’ organelles are represented by all three types of secretory granules within (α–, δ–, and lysosomes), which are produced by the Golgi apparatus of the MKs, a dense tubular system (an inner smooth endoplasmic reticulum membrane system which initiates and modulate platelet activation), and the open canalicular system (a surface-connected extension of the plasma membrane), which helps to convey and distribute external substances into the platelets or helps to discharge α–granules [137].
However, the most prominent organelles are the α–granules. They are produced synthetically (by the trans-Golgi network), or by endocytosis (by MKs’ endocytic abilities) [138]. Ultrastructural studies indicated a heterogeneous population of α–granules within one platelet [137], even though scanning electron microscopy suggests they are homogenous ovoid alpha granules of about 200–500 nm [139]. The alpha granules have a different cargo load, which is released by platelets in a kinetically heterogeneous way [140,141,142]. Proteomic studies (including mass spectrometry) have identified several hundred (generally accepted to be over 300 proteins) soluble proteins within the α–granules, including the α–granules’ specific proteins [138,143,144,145]. Among them are some mediators of blood coagulation; the von Willebrand factor, fibrinogen, factors V, XI, and XIII, prothrombin, plasminogen activator inhibitor-1 (PAI-1), thrombospondin (TSP1), etc. [138,144,146,147]. Thus, the α–granules are involved in maintaining the blood homeostatic balance, as they are involved in regulating bleeding diathesis via the pro- and anticoagulant proteins stored within their granules [148]. Moreover, the involvement of α–granules within inflammatory processes was previously documented. The α–granules contain P-selectin and several chemokines (e.g., CXCL1, CXCL4, CXCL7) that either mediate the interaction between platelets and endothelial cells or are involved in platelets’ activation or granular content secretion, or maintain the inflammatory state.
Notably, the α–granules contain a few very important proangiogenic molecules (e.g., angiopoietin, CXCL12/SDF-1α, and matrix metalloproteinases (MMP-1, -2, and -9)), but also angiogenic inhibitors (Thrombospondin-1) that inhibit endothelial cells’ proliferation, promoting their apoptosis [149,150,151,152].
The dense granules (δ–granules) are platelet-specific organelles. They are smaller (approximately 150 nm) and numerically less abundant than the α–granules [153,154]. The δ–granules arise from the early endosomal network, concomitantly with the α–granules [155], and contain serotonin, histamine, uracil, guanine nucleotides, serotonin, calcium, potassium, and polyphosphates [156]. The δ–granules’ proteome contains 40 proteins, including regulatory proteins [157,158,159]. The molecules stored within dense granules are involved in thrombosis and blood homeostasis. The δ–granules’ deficiency usually affects bleeding, by affecting platelet aggregation and thrombus formation (thus increasing the bleeding tendency) [153,160,161]. Moreover, serotonin is marked by both vasodilator and vasoconstrictor properties, and is an important player in vascular tone regulation in humans [162]. However, δ–granules may be involved in promoting vascular permeability in inflammation [163,164,165].
The platelet’s lysosomes are heterogenous and, usually, their diameter ranges between 200 and 250 nm, similar to those in other cells. The lysosomal membrane has positivity for LAMP-1, LAMP-2, and CD63, and their content is embodied by degradative enzymes (the acid hydrolases) belonging to proteins or carbohydrate-degrading enzymes or phosphate esters cleavers [166,167,168,169].
To secrete their granules, the lipid membranes of the granules and the platelet plasma membrane must fuse (by means of the lipids and phospholipids), and these fusions are governed by the Soluble N-ethylmaleimide-Sensitive Factor Attachment Proteins Receptors (SNAREs) [170]. However, the granule secretion is supported by the elevation of intracellular Ca2+ and regulated by proteins like calmodulin and calcyclin [171,172].
Platelets are produced within hematopoietic bone marrow from MKs that have a characteristic immunopositivity for thrombopoietin, (also known as Mpl ligand, Mpl-l, or the MK growth and development factor, MGDF), CD34, and CD38 [67,173,174,175,176]. Flow cytometry confirmed the presence of microparticles and small CD41-positive cellular fragments in the vicinity of MKs’ platelet-producing cellular prolongations. They are presumed to be related to platelet activation and/or their physiology [177,178]. Platelets consist of glycoproteins (GpIb, GpIIb-IIIa, P-selectin) that are characteristically distributed at the level of the platelet plasma membrane or platelet organelles [179,180].
4.3. Roles of Platelets
Within their granules, platelets store proteins but mainly growth factors, and the degranulation of these promote local tissue homoeostasis, through platelets’ membrane adhesion and subsequently their aggregation, clot formation, and their granular content release to promote tissue repair/regeneration [36].
Due to the physical parameters of the blood, and their shape, platelets can be mainly found in proximity to the luminal surfaces of endothelial cells, where natural platelet inhibitors are present [75]. This localisation mediates the platelets’ tracking of endothelium integrity, allowing them to rapidly react to endothelial integrity issues through adherence, changing shape, aggregation, and the secretion of the growth factors they contain. In graft tissue, platelets can interact with the endothelium of blood vessels after the transplant, helping leukocytes to accumulate [181,182,183,184]. Through direct physical interaction, activated platelets can transfer chemokines to the atherosclerotic endothelium or to inflamed endothelial cells that can, after that, performantly attract leukocytes (mainly neutrophils) [185].
One physiologic role of platelets is to maintain blood homeostasis, as they are involved in the inflammatory response and adaptive immunity. The close interaction between the platelets and subsets of leukocytes (granular or agranular) has already been documented, as has the secondary secretion of immunomodulatory proteins [186]. The physical interaction between platelets and leukocytes is mainly mediated by P-selectin and PSGL-1; this interaction contributes to a performant leucocyte recruitment with the formation of circulating leukocytes–platelets conjugates that produce fibrin [187]. In the physical interaction of platelets with different types of leukocytes, the platelets’ P-selectin is responsible for the activation of different integrins: e.g., Mac-1 (αMβ2) and LFA-1 (αLβ2) on neutrophils, or β1 and β2 integrins on monocytes and lymphocytes. Moreover, P-selectin can produce a delayed response by triggering some gene expressions, and subsequently protein synthesis, producing, in the case of leucocytes, the acquiring of an inflammatory phenotype [188]. The modulatory effects of this interaction are reciprocal, mainly regulating cell cellular functions via proteases, oxygen radicals, or nitric oxide [189]. The platelet–leukocyte complex functions in the setting of the endothelial lesion, as this interaction is a prerequisite for the hyperplasia of the blood vessels intima after a vascular injury [190,191], or, for example, contributes to the progression of atherosclerosis or cardiac dysfunction [192].
The secretion of the granular cargo (especially from the α–granules) contains mainly mitogenic, coagulation, and angiogenic factors, as well as inflammatory mediators. The content of the platelet’s vesicular cargo may act either locally, influencing the activity of neighbouring cells, or systemically [193]. The platelet’s extracellular vesicles are small (100–250 nm) membranous structures produced and shedded by platelets under different stimuli (Figure 4). These extracellular vesicles (cargos) contain RNA, MMPs, antigens, and cytokines [194].
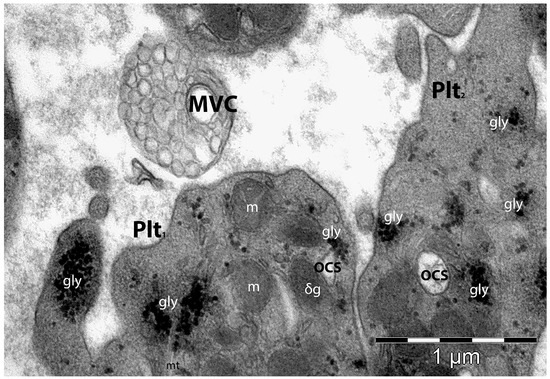
Figure 4.
Transmission electron microscopy image of two neighbouring platelets (Plt1 and Plt2). Plt1 is in close proximity to a just-shedded multivesicular cargo (MVC)—a complex group of platelet-derived membrane-bound structures that activate and regulate intercellular pathways. MVC are vectors of transport for nucleic acids, proteins, and lipids, regulating the local microenvironment (but also remotely) with implications in cellular functions’ modulation or tissue regeneration/repair. gly—glycogen; m—mitochondria; OCS—open canalicular system. (Courtesy of Dr. E.T. Fertig, “Victor Babeş” National Institute of Pathology, Bucharest, Romania).
However, platelets are essential for angiogenesis and maintaining blood vessel integrity via several tyrosine-based activation motifs (ITAM) that regulate cellular interactions [195]. The lipids from the structure of the platelet’s membrane (e.g., phosphatidic acid, lysophosphatidate, S1P) are cellular chemoattractants that are involved in angiogenesis [196].
Platelets can regulate blood vessel permeability. Thus, a severe depletion of platelet numbers affects the endothelium integrity, with the appearance of disruptions and fenestrations that permit red blood cells to cross into the surrounding tissue and to form petechiae [164]. The angiogenic role of the platelets is supported not only by their number, but also by their functionality, and their direct contact with endothelial cells, a process which is favoured by a series of adhesion molecules [197]. However, within platelets’ granules, among their angiogenic factors there are also anti-angiogenic factors (e.g., endostatin, thrombospondin-1, angiopoietin-1, angiostatin, etc.), contributing to the vascular homeostasis in the repairing tissue [198]. Upon platelet activation or due to oxidative stress, by membrane budding, platelets release microparticles that are mostly found at the site of angiogenesis [108,199]. Thus, they stimulate new blood vessel formation or increase vascular permeability. Platelet microparticles are also found frequently within solid tumour microenvironments, where they are involved in protein transfer or RNA/DNA transfer to endothelial cells [151].
At the level of vascular injury, platelets, via their secreted signalling molecules, are producing chemoattraction and the densification and numeric proliferation of the neighbouring smooth muscle cells, and with SDF-1α, differentiation into endothelial cells and vascular sprouting [200,201,202]. Previous data reported that there is an optimal concentration of platelets (1.5 × 106 platelets/μL) that induces angiogenesis in human endothelial cells; exceeding this concentration of thrombocytes could exert an inhibition of angiogenesis [203].
Platelets are also important in lymphangiogenesis since, similar to lymphatic endothelial cells (but different from blood vessels’ endothelial cells), they have a positive membrane expression for C-type lectin-like receptor (CLEC)-2, which ligates podoplanin [204,205]. Through this, they can bind podoplanin (a type I transmembrane sialomucin-like glycoprotein), which can lead to platelet activation, but the deficiency of podoplanin impairs lymphatic vessel formation [206,207]. Platelets are also involved in the embryonic differential development of lymph vessels and blood vessels, while also maintaining this separation (e.g., by platelets’ plugs), or being involved in further lymphatic vessel-repairing processes [208,209,210].
In the process of endothelial cell differentiation, platelets act as recruiters for progenitor cells, which, after that, in an SDF-1/CXCR4 manner, become functional endothelial cells [211]. One layer of platelets could realise leukocyte recruitment or contribute (by granular P-selectin) to monocytes’ recruitment, thus it could be rational to highlight the task of platelets in supporting neointimal formation, or intimal hyperplasia [212,213,214].
Platelets also secrete numerous chemokines involved in tumour cells’ invasion, tumoral cell proliferation, or metastasis regulation [215]. Among the series of GFs that the granules contain, PDGF and TGFβ1 appear to be integral modulators [36]. Activated platelets, via the key molecules and GFs they are secreting, are contributing, along with other immune cells, to all four stages of tissue repair, specifically to haemostasis, the inflammatory/immune response, tissular proliferation, and tissue remodelling [216,217,218,219,220]. Within a lower pH micro-environment (that features the initial stages of wound healing) platelets seem to increase the concentration of PDGF and therefore stimulate the proliferation of Fb, since in the same pH range TGFbeta1 increases the production of collagen. Later, by releasing IL1, platelets mitigate inflammation, promoting dense scar formation [36].
5. A Concise Evolution of the PRP Therapy Concept
With comparative results to those hypothesized for stem cell therapies, PRP is quickly becoming a pillar of regenerative medicine. Its applications are expanding into diverse medical fields, such as aesthetic dermatology, orthopaedics, sports medicine, and surgical procedures.
Everything started more than a century ago, in 1842, with Donne’s ground-breaking revelation about blood containing structures beyond the erythrocytes and leukocytes [221]. It was Julius Bizzozero who initially coined the term “le piastrine del sangue” for platelets. In 1882, he documented the pivotal role of platelets in in vitro blood coagulation and their contribution to in vivo thrombosis. He also observed that the vascular wall could inhibit platelet adhesion [222,223]. Advancements in regenerative therapy were further made by Wright’s identification of MKs as platelets’ progenitors [224].
During the 1940s, medical professionals employed “embryonic extracts” (rich in growth factors and cytokines), to foster wound healing [225]. Successful surgical procedures rely, inter alia, on prompt and efficient wound healing, Thus, in April 1944 Eugen Cronkite et al. proposed the idea of using thrombin and fibrin in skin grafting [226,227]. By using these components, the firmness and stability achieved are integral to grafting surgeries.
At that time, the concept of platelets secreting growth factors in PRP and thus contributing to PRP’s efficacy emerged as a hypothesis. This theory was validated in the 1980s when it was established that the bioactive molecules (or GFs) discharged from platelets repair damaged tissues, such as skin ulcers [228,229]. Numerous studies have been published onto this topic since then. A prominent area of research in this domain has been the synergistic use of PRP with hyaluronic acid [230]. In 1962, Cohen identified the epidermal GF (EGF). This discovery paved the way for the subsequent finding of growth factors: platelet-derived GF (PDGF) in 1974 and vascular endothelial GF (VEGF) in 1989 [224]. Medical advancements, in a broader sense, have also spurred rapid progress in platelet applications. In 1972, Matras pioneered the use of platelets as sealants to maintain blood homeostasis during surgeries [231]. Later, in 1975, Oon and Hobbs were the pioneers in applying PRP in reconstructive treatments [232].
In 1986, Knighton and his colleagues pioneered the description of platelet concentrate protocols, coining the term “autologous platelet-derived wound healing factors” [233]. Following the establishment of these protocols, the technique started to become very popular in the field of aesthetic medicine [234]. Since the latter part of the 1980s, PRP has been a staple in the field of regenerative medicine and regenerative dermatology [229].
Beyond its applications in general and cardiac surgery, PRP found acclaim in maxillofacial surgery during the early 1990s. It was utilized to enhance graft integration during mandibular reconstructions [235]. The dental industry also recognized PRP’s potential. By the late 1990s, PRP began to be employed to support the integration of dental implants and support bone regeneration [236]. A related product, fibrin glue, was also introduced during this period [237]. A significant advancement in dental PRP applications came with Choukroun’s invention of platelet-rich fibrin (PRF), a unique platelet concentrate that eliminated the need for anticoagulants [233]. The 2000s witnessed a surge in PRF’s popularity, with its use spanning various dental treatments [238].
From 2010, and possibly even before, PRP has made notable inroads into cosmetic dermatology. Post-PRP injection, there is a marked rejuvenation of the skin, characterized by enhanced hydration, elasticity, and improved complexion [239]. An added application of PRP has been its use in augmenting hair growth [240]. The studies of Gentile et al. highlighted that hair density and count can be positively influenced by PRP injections [241]. Additionally, a pre-treatment with PRP prior to undergoing hair transplantation has been shown to boost hair growth and enhance the overall density of the hair [242,243].
Recent advancements in cosmetic dermatology show that integrating PRP with CO2 laser treatments leads to a pronounced decrease in acne scarring [244,245]. Moreover, the combination of PRP with micro-needling has been observed to produce more structured collagen formations in the skin, compared to using PRP by itself.
The application of PRP in medicine could be extensive, with notable contributions to various medical disciplines, including gynaecology [246], urology [247], and ophthalmology [248]. Overall, before mentioning the applications (and presumable benefits) of PRP in skin conditions, it is worth mentioning that neo-angiogenesis plays a primary role in the formation of the granular tissue and in maintaining keratinocytes’ survival rate. Thus, many prophylactic or therapeutic platelet transfusions are performed with activated platelets. When discussing PRP treatments, we need to differentiate between different platelet concentrates, based on their cell content and fibrin architecture, such as those presented in Table 1.

Table 1.
Classification proposed by Dohan Ehrenfest et al. for platelet concentrates based on their cell content and fibrin architecture [249,250].
A new coding system for PRP was designed by Kon E. et al. to simplify the description of PRP characteristics and ensure consistency and clarity in its preparation and use. Here is a breakdown of the PRP coding system [251]:
- N1N2—platelet composition:
- -
- The pair of digits relates to the concentration of platelets in the PRP compared to the basal levels in the blood.
- -
- “1” represents a concentration range of 100,000–200,000 platelets/μL.
- -
- “2” represents a concentration range of 200,000–300,000 platelets/μL.
- -
- So “12” would mean that the PRP has twice the basal concentration of platelets, and the PRP’s platelet concentration is between 200,000–400,000 platelets/μL.
- N3N4—purity:
- -
- N3 indicates the absence (0) or presence (1) of erythrocytes (RBCs).
- -
- N4 represents the concentration of leukocytes, with “0” indicating no leukocytes and increasing numbers indicating higher concentrations.
- -
- For example, “11” would indicate the presence of erythrocytes and a low concentration of leukocytes in the PRP.
- -
- Both leukocytes and erythrocytes can affect the results obtained after PRP injections due to side effects and interferences [252].
- N5N6—activation:
- -
- N5 specifies whether the PRP is endogenously activated (0) or whether it is activated externally, before PRP injection (1).
- -
- N6 indicates whether calcium was added for activation, with “0” meaning no calcium added and “1” meaning calcium was added.
- -
- For instance, “00” would mean that the PRP is endogenously activated, and no calcium has been added.
- -
- Previous research has indicated that the level of calcium can impact the cellular and tissue reactions elicited by PRP [253].
6. Utility of PRP in Dermatological Practice
6.1. PRP in Skin Burns
Skin burns are lesions caused by the action of high temperature agents over the skin, and can involve the epidermis only (superficial burns), the epidermis and superficial, papillary dermis (partial thickness burns), or deep skin burns (that involve the reticular dermis) [32]. The burn lesion has definitive stages of evolution: at 7 days of evolution it is characterised by acidophilic material covering the wound surface and dermal inflammation, with scarce CD34-positive cells; at 14 days, focal re-epithelization of the dermis and wound healing appear; and at 28 days a flawed epithelium with keratinocytes’ nuclear pyknosis cytoplasmic vacuolization is formed [254].
PRP plays a significant role in enhancing the healing and appearance of post-burn scars by improving pigmentation, skin texture, and alleviating associated neuropathic pain. This positive effect is primarily attributed to the growth factors in PRP, which promote neovascularization and neo-collagenogenesis, thereby aiding the healing process in burn wounds [255,256]. The application of PRP can be achieved using various injection techniques (the “linear retrograde and fanning” method, the “tunnelling” technique, etc.). Published data indicate that monthly PRP sessions over three months of treatment produced remarkable enhancements in terms of itch relief, scar pigmentation, and suppleness. The improvement in pain is believed to result from nerve growth factors present in the PRP, promoting nerve recovery and regeneration [257].
The PRP product injected in the skin burn wound, at 7 days after injection, determined the covering of the wound area by migrated spindle-shaped cells from the wound periphery with heterogeneous disorganised dermal collagen fibres [258,259]. Fourteen days after the PRP injection, the epidermis indicated a normal appearance but the dermis was the site of a prolific synthetic fibroblast-like infiltrate with an increased density of blood vessels [260]. However, the lesion injected with PRP had an increased number of CD34-positive cells at 14 and 28 days after the injection [261].
Overall, the beneficial effects of PRP administration are epithelization, endothelial cell migration, and neovascularization, increasing the production of collagen fibres and dermal matrix proteins, and improving dermal regeneration [262]. PRP upregulates the dermal expression of collagen I and downregulates the matrix metalloproteinase proteins 1 and 9 (MMP-1 and MMP-9) [263].
6.2. PRP in Alopecia
Hair on the scalp significantly contributes to an individual’s self-perception. Individuals with alopecia experience noticeable physical changes, leading to potential psychological distress and reduced self-confidence [264]. Hair loss, also called alopecia, stands as a prevalent concern among those seeking dermatological advice, and its duration can range from temporary to permanent [265]. Alopecia can be broadly categorised into non-cicatricial and cicatricial types. Androgenetic alopecia, telogen effluvium, alopecia areata, trichotillomania, and anagen effluvium are examples of non-cicatricial alopecies. On the other hand, conditions like lichen planopilaris, frontal fibrosing alopecia, folliculitis decalvans, and cutaneous discoid lupus erythematosus contribute to cicatricial alopecia [266].
For an effective alopecia treatment, it is crucial to conduct a thorough assessment, including an in-depth review of dietary habits, ensuring tailored patient care [267]. Clinicians have a variety of traditional treatment methods at their disposal to address alopecia. For instance, androgenetic alopecia treatments comprise topical minoxidil, oral medications like finasteride and dutasteride, peptides, and hair transplant procedures [268]. To address telogen effluvium, approaches such as high-protein diets, iron supplements, avoiding catagen-inducing medications like beta-blockers and retinoids, and treating underlying conditions that promote catagen (like thyroid issues or hyperandrogenism) are recommended [269].
With increasing interest in regenerative therapies, PRP has emerged as a promising treatment option for various types of alopecia, attributed to its high concentration of growth factors and cytokines. As it is derived from the patient’s own blood (autologous), PRP eliminates the potential adverse reactions often associated with other therapeutic methods, thus enhancing patient compliance. Moreover, because PRP operates differently from other treatments, it could further enhance results when used alongside treatments like minoxidil and finasteride [270]. PRP is derived from autologous blood and is concentrated with platelets that release various GFs, PDGF, TGF-β, VEGF, and EGF, which play a pivotal role in the hair cycle. They enhance the proliferation of dermal papilla cells, stimulate angiogenesis, and prolong the anagen (growth) phase of the hair cycle [242].
6.2.1. Androgenetic Alopecia (AGA)
Commonly referred to as male and female pattern baldness, androgenic alopecia results from genetic predisposition and hormonal influences. Studies have shown that a PRP treatment increases hair density and promotes thicker hair shafts in androgenic alopecia patients. The growth factors in PRP likely counteract the effects of dihydrotestosterone (DHT), the primary hormone responsible for androgenic alopecia [271].
Singhal et al. reported, after three months of PRP treatment, that all ten patients with androgenetic alopecia displayed significant hair growth, witnessing an average 65% decrease in hair loss. Remarkably, six patients saw new hair growth in just 7 days, while the rest observed it within 15 days [272]. In another study involving 11 AGA patients undergoing PRP treatment, the hair pull test was negative for 9 patients after four PRP sessions held bi-weekly. Also, a noticeable improvement in hair thickness and fullness was reported [273]. Furthermore, Greco et al. conducted a study with 10 AGA patients—5 treated with PRP and 5 untreated. Their findings suggest that using PRP as mesotherapy in AGA results in a significant boost in both hair thickness and density [274,275].
Uebel et al. noted a marked increase in hair density and growth when follicular units were pre-treated with platelet plasma growth factors prior to implantation. A noticeable difference in the success rate of follicular units was seen when comparing treated and controlled scalp areas [243]. A post-slitting intra-operative PRP treatment was found by Garg et al. to boost hair density swiftly, decrease transplanted hair’s catagen loss, speed up skin recovery, and activate dormant follicles in follicular unit extraction transplant subjects compared to a placebo [276].
Generally, hair growth necessitates performant vascular support and the development of this could be influenced by angiogenesis due to the production of VEGF by keratinocytes and papillary fibroblasts [277]. PRP induces follicular stem cells’ differentiation, and the anagen phase’s prolongation, stimulating hair growth via upregulations of the FGF-7 and β-catenin signalling pathways [278] Previous studies reported positive therapeutic effects on hair loss (male/female pattern) after interfollicular injections of CD34-positive cells and PRP [279]. Autologous PRP seems to increase the proliferation of dermal papillary cells, while also increasing the expression of fibroblast growth factor 7 (FGF-7), β-catenin, extracellular signal-related kinase (ERK), and Akt signalling, further suggesting a mechanism for hair restoration [280,281]. Likewise, a systematic review by Leo et al. supported the beneficial effects of PRP in hair regrowth and highlighted its safety profile [282].
6.2.2. Alopecia Areata (AA)
AA is an autoimmune disorder causing patchy hair loss. Trink et al. demonstrated that PRP induced significant hair regrowth in androgenic alopecia patients, potentially due to the immunomodulatory effects of the GFs present in PRP [283]. Also, evidence suggests that PRP might offer therapeutic benefits for AA patients by reducing local inflammation and stimulating hair regrowth [284].
PRP was successfully used in AA [285,286]. Administering PRP to hair skin improves the skin ischaemic conditions by increasing the density of vascular structures within the hairy skin, thus promoting cellular proliferation and differentiation [287]. Greco et al. employed PRP as mesotherapy for a single patient with AA, yielding positive outcomes after 10 months [274]. However, a comprehensive study with 45 patients demonstrated PRP’s efficacy in treating AA, showcasing significant hair regrowth, reduced hair abnormalities, and minimised scalp discomfort [283]. Singh et al.’s research revealed that of 20 AA patients treated with PRP, only one experienced a relapse, and the treatment was well-received [288]. Donovan pointed out the potential of PRP in treating specific steroid-resistant forms of AA [289]. El Taieb et al. found PRP to be more effective than topical minoxidil 5% in treating AA based on their study involving 90 patients [290].
6.2.3. Telogen Effluvium (TE)
TE is often caused by stress, medications, or hormonal changes, and is characterised by diffuse excessive shedding. While fewer studies have focused on PRP’s efficacy in TE, preliminary reports suggest improvements in hair density and a reduction in hair shedding post-PRP treatment [291,292].
The efficacy of PRP in treating TE lacks concrete research evidence. It was previously documented that minoxidil increases the ratio of Bcl-2/Bax by activating ERK and Akt [293,294]. On the other hand, finasteride induces the activation and prolongation of the anagen phase of hair growth, which is responsible for the longitudinal and diametral growth of hairs, but it also induces the expression of apoptosis inhibitors and caspase [295].
Studies comparing PRP with other treatments, like minoxidil or finasteride in AA, suggest that while both treatments may offer benefits independently, the combination of PRP with traditional therapies often produces superior results [296]. Fakahany et al. observed that automated micro-needling combined with PRP effectively treats various hair loss conditions, including cicatricial alopecia [297]. The success of grafted follicles in cicatricial alopecia areas, which typically have a reduced blood flow, relies primarily on the vascular bed’s blood supply. PRP, laden with growth factors like IGF-1, βFGF, and VEGF, can ameliorate cutaneous ischaemic conditions, enhancing the hair follicles’ surrounding vascular structures [298]. Saxena et al. found that introducing 1 mL of PRP intradermally to a recipient area test patch just before grafting was effective in managing cicatricial lichen planus [299]. In cicatricial alopecia cases, the scarcity of ostia might suggest a limited potential for restoring destroyed hair follicles. Thus, new follicles from follicular unit extraction punch grafting might serve as a stem cell source, APM, and nerve bundle.
6.3. PRP in Skin Ageing
The relentless search for non-invasive rejuvenating therapies has brought PRP to the forefront of aesthetic dermatology and rejuvenating medicine. This autologous product from whole blood harnesses a concentrated cocktail of growth factors and cytokines, presenting a biocompatible and effective approach to combat skin ageing. The regenerative potential of PRP is mainly attributed to its rich milieu of growth factors, among them the PDGF that stimulates fibroblast growth and promotes collagen production, improving the skin’s texture and elasticity [300]. TGF-β enhances the synthesis of the extracellular matrix and plays a role in tissue remodelling [301]. VEGF boosts angiogenesis, ensuring the better oxygenation and nourishment of skin tissues [302]. EGF facilitates cell growth and differentiation, leading to epidermal and dermal regeneration [303]. FGF promotes fibroblast function and augments skin repair mechanisms [304].
All the aforementioned growth factors are only a few of those provided by the platelets within PRP. Together, they stimulate collagen synthesis, enhance tissue regeneration, reduce inflammation, and amplify skin hydration—the well-acknowledged hallmarks of youthful skin. PRP therapy has demonstrated efficacy in mitigating wrinkles and enhancing skin elasticity, possibly due to increased collagen production post-therapy [305]. PRP treatments can improve skin tone and firmness, making it a viable option for those with sagging skin [306]. With its capacity to stimulate neo-collagenesis, PRP can lead to smoother skin and a reduction in pore size [307]. PRP’s angiogenic properties can rejuvenate the skin’s microcirculation, leading to brighter, more radiant skin [308]. PRP emerges as a promising treatment modality for skin ageing, riding on its autologous nature and multiplicity of regenerative factors. However, the procedure’s success is influenced by factors like PRP preparation techniques, individual patient differences, and treatment protocols. While preliminary results are encouraging, larger-scale, randomised studies are essential to firmly establish PRP’s role in anti-ageing treatments.
Notable benefits of PRP’s administration on facial skin, like skin suppleness, texture, hydration, wrinkle reduction, and micropigmentation, were observed between 1 and 3 months post-treatment [305,309]. In one specific study, the outcomes persisted for 6 months after completing three PRP sessions, although there was a slow reversion to the original state [310]. Previously published data have indicated the successful administration of PRP as a single treatment for periorbital wrinkles and dark circles, an improvement of the perioral area, skin fitness enhancement, and a reduction in skin redness, revealed an improvement in skin texture and increased patient satisfaction for crow’s feet, and a notable enhancement in skin colour for infraorbital dark circles [305,306,309,311,312,313,314,315].
6.4. PRP in Acne Scarring
Acne scarring, a significant cosmetic concern, can severely impact an individual’s self-esteem. With the rise of regenerative medicine, PRP has shown promise as a treatment modality for acne scars. PRP, an autologous preparation derived from whole blood, offers a rich mix of growth factors that can foster tissue repair and regeneration. The PRP growth factors initiate collagen production, stimulate angiogenesis, and boost tissue repair, making PRP a potent tool against acne scars.
PRP has demonstrated the potential to enhance the texture and appearance of atrophic scars by amplifying collagen synthesis and remodelling [316]. PRP can be combined with other treatments like micro-needling and laser therapy to synergize their effects and achieve improved outcomes. Micro-needling with PRP has shown superior results in reducing scar depth and improving skin texture compared to micro-needling alone [317].
PRP’s anti-inflammatory properties can accelerate healing and reduce the downtime post resurfacing procedures like lasers or dermabrasion [318], thus emerging as an effective therapeutic option for acne scars, as can be seen in Table 2. Its autologous nature minimises the risk of allergic reactions or incompatibility. While initial results are encouraging, the optimal preparation techniques, concentrations, and treatment protocols are still subjects of ongoing research. Large-scale, randomised trials are warranted to conclusively establish PRP’s role in acne scar management.
Table 2.
Studies on acne scar treatments with PRP.
6.5. PRP in Melasma
Melasma is a persistent hyperpigmentation disorder characterised by an overproduction of melanin by melanocytes, resulting in yellow-brown patches, particularly on sun-exposed skin (centrofacial, malar, or mandibular) [328,329]. The exact cause remains unclear, but it is believed to be a combination of factors including UV exposure, hormonal changes, inflammation, and oxidative stress that influences skin cells, leading to melanogenesis [330]. The prevalence of melasma varies largely [331]. It predominantly affects women, with a 9:1 female-to-male ratio, and particularly those in their reproductive years, pregnant, or with skin phototypes III-V [329,332]. Common treatments encompass topicals like hydroquinone and azelaic acid, systemic treatments like glutathione, and procedures like bleaching and lasers, although these treatments can sometimes lead to inconsistent outcomes and skin reactions [328].
PRP’s potential in treating melasma stems from its wound healing properties and the angiogenic effects of its growth factors, PDGF, TGF-β1, and TGF-β2. Specifically, TGF-β1’s ability to inhibit tyrosinase—an enzyme crucial for melanin production—might be key. This inhibition might be due to its effects on transcription factors such as microphthalmia associated transcription factor (MITF) and through the prolonged activation of the extracellular signal-related kinase.
However, the current literature on PRP for melasma is limited, with only a few studies including a meta-analysis, a non-randomized clinical trial, and a prospective study available for review. Even though all these studies utilised the Melasma Area and Severity Index (MASI) to assess outcomes, the treatments varied—ranging from PRP injections and tranexamic acid to lasers and micro-needling. Overall, incorporating PRP in combination treatments like fat grafting and CO2 lasers has shown marked progress in post-burn scar appearance [333].
6.6. Limitations of PRP
PRP therapy shows encouraging results in treating various skin conditions. However, there is a pressing need for extensive, randomized, controlled trials. Most existing studies are limited by small sample sizes, inconsistent criteria for preparation protocols and outcome comparisons, and brief follow-up durations. Despite numerous studies, the impact of different parameters on PRP’s clinical effectiveness in aesthetic medicine remains unclear. A primary issue in evaluating PRP’s effectiveness across studies is the lack of standardization. Another significant challenge is the inconsistency and inadequacy of assessing outcomes. Further research is essential to establish the most effective methods and procedures for collecting, processing, and administering PRP.
The significant variability in PRP preparation protocols has been a recurrent topic of discussion [38,334,335]. One reason for this variation is the array of classification systems proposed. In 2009, Ehrenfest et al. [336] introduced the first such system, categorizing platelet-rich preparations based on two factors: the presence of leukocytes and the density of the fibrin network. Thus, we can divide PRP in four categories [296]: pure PRP/P-PRP (leukocyte-poor, low-density fibrin network), leukocyte-rich PRP/L-PRP (leukocyte-rich, low-density fibrin network), pure PRF/P-PRF (leukocyte-poor, high-density fibrin network), and leukocyte-rich PRF/L-PRF (leukocyte-rich, high-density fibrin network).
Additionally, some experts advocate for using the DEPA classification system by Magalon et al. [337], which categorizes different PRP products based on the dosage of injected platelets, the efficiency of platelet recovery from blood, the purity of the PRP (ratio of platelets to red and white blood cells), and the activation process [296].
7. Further Perspectives
It is already (clinically) acknowledged, in few different conditions (including dermatological disorders), that the administration of PRP has tremendous clinical or physiognomic benefits. However, we presume that the roles, the mechanisms of action, and the structural advantages of PRP delivery are still waiting to be approved and intimately described. It is our assessment that PRP’s involvement in regenerative dermatology is still at the beginning of the slope of enlightenment. The simplest acceptance that PRP is just a collagen stimulator is marked by prosaicism. There is still a need for more solid data to decipher the cellular and molecular machinery that explains PRP’s clinical benefits and reveals the complexity of the intricate reparatory/regeneration mechanisms that PRP is producing.
Since the first documentation of PRP’s clinical benefits on skin repairing/regeneration to the present day, much important data have been reported as well as a few essential changes in skin structural science (with respect to cellular and molecular dermatology). Presumably, some of the changes are still hiding those rational explanations for the PRP advantages that should be considered. Indubitably, the local application of trophic factors within the dermis promotes cell division and the synthesizing of cellular components of the dermal microenvironment, structurally and secondarily impacting the epidermis.
At least for skin science (but many times proved within the stroma of other organs), the presence of a new population of interstitial cells has already been identified—telocytes (TCs) [338,339,340]. A great body of already published data has (ultrastructurally and immunohistochemically) described skin TCs and also documented the involvement of TCs in neo-angiogenesis and in the tissular reparatory/recovery/remodelling/regeneration process [340,341,342]. However, to date, those solid structural and ultrastructural studies of tissue regeneration/repair after PRP administration are still insufficient. Thus far, the clinically acknowledged data on the PRP benefits in skin (patho)physiology include no correlation in between dermal-resident TCs and the beneficial PRP-induced potential for skin regeneration/repair. The growth factors and the substances that PRP locally provides could influence the local dermal TCs into generating the targeted tissue regenerative outcome. Moreover, the presumptive correlation between TCs and PRP could offer new insights into the skin adnexa regeneration processes, or reparatory processes in other organs (Figure 5). However, an interesting idea was promoted with regard to a cellular collaboration between the stem cells and TCs in fulfilling the regeneration process [343].
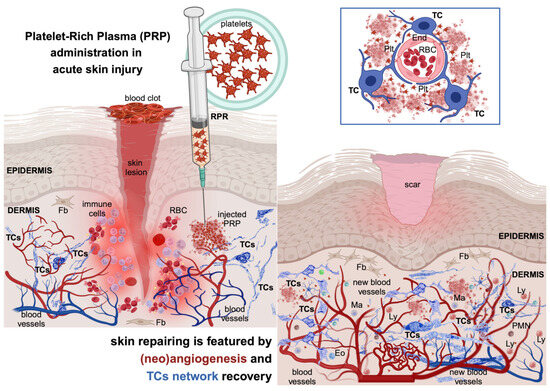
Figure 5.
Providing a platelet-rich plasma (PRP) treatment to the site of the skin injury helps the structural recovery in terms of speediness and the performant regeneration of affected cutaneous components. Considering the myriad of growth factors and cytokines, it is attractive to presume that PRP is eliciting the vascular recovery of telocytes (TCs), cells with a documented angiogenic potential within the interstitia of other organs. Inset: platelets could stimulate the involvement of TCs (by their cellular prolongations—telopodes) as nursing cells for the new-formed blood vessels. Plt—platelets; RBC—red blood cells; End—endothelial cell; Ma—macrophage; Fb—fibroblast; Ly—lymphocyte; Eo—eosinophil; PMN—neutrophil. The image was created with BioRender.com.
However, we presume that documenting any possible collaboration between TCs and PRP and the combined administration of PRP and interstitial cells for skin wound treatments, or the regeneration of skin defects, will be a provoking direction for research. Unpublished results have indicated that, in cardiac failure, the heart transplantation of interstitial cell suspensions (enriched in TCs) especially improved cardiac performance. Moreover, there is a current trend for the dermal administering of stimulators of skin’s natural collagen production that gradually contribute to a better-looking appearance of skin. These stimulators are in fact producing local dermal inflammation and probably changes in local dermal TCs that favour inflammation, immune cellular milieu, new blood vessel formation, and finally, collagen production.
Funding
This work was funded by the Ministry of Research, Innovation and Digitalization in Romania, under Program 1—The Improvement of the National System of Research and Development, and Subprogram 1.2—Institutional Excellence-Projects of Excellence Funding in RDI, Contract No. 31PFE/30.12.2021 and UEFISCDI project PN-III-P4-ID-PCE-2020-2300, grant no. PCE43/2021.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Someya, T.; Amagai, M. Toward a New Generation of Smart Skins. Nat. Biotechnol. 2019, 37, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Maranduca, M.; Branisteanu, D.; Serban, D.; Branisteanu, D.; Stoleriu, G.; Manolache, N.; Serban, I. Synthesis and Physiological Implications of Melanic Pigments (Review). Oncol. Lett. 2019, 17, 4183–4187. [Google Scholar] [CrossRef] [PubMed]
- Sorg, H.; Tilkorn, D.J.; Hager, S.; Hauser, J.; Mirastschijski, U. Skin Wound Healing: An Update on the Current Knowledge and Concepts. Eur. Surg. Res. 2017, 58, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.A.F. Fibrin Is a Many Splendored Thing. J. Investig. Dermatol. 2003, 121, xxi–xxii. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.; Kosaric, N.; Bonham, C.A.; Gurtner, G.C. Wound Healing: A Cellular Perspective. Physiol. Rev. 2019, 99, 665–706. [Google Scholar] [CrossRef]
- Wilgus, T.A.; Roy, S.; McDaniel, J.C. Neutrophils and Wound Repair: Positive Actions and Negative Reactions. Adv. Wound Care 2013, 2, 379–388. [Google Scholar] [CrossRef]
- Park, J.E.; Barbul, A. Understanding the Role of Immune Regulation in Wound Healing. Am. J. Surg. 2004, 187, S11–S16. [Google Scholar] [CrossRef]
- Koh, T.J.; DiPietro, L.A. Inflammation and Wound Healing: The Role of the Macrophage. Expert. Rev. Mol. Med. 2011, 13, e23. [Google Scholar] [CrossRef]
- Davies, L.C.; Jenkins, S.J.; Allen, J.E.; Taylor, P.R. Tissue-Resident Macrophages. Nat. Immunol. 2013, 14, 986–995. [Google Scholar] [CrossRef]
- Armulik, A.; Genové, G.; Betsholtz, C. Pericytes: Developmental, Physiological, and Pathological Perspectives, Problems, and Promises. Dev. Cell 2011, 21, 193–215. [Google Scholar] [CrossRef]
- Ansell, D.M.; Izeta, A. Pericytes in Wound Healing: Friend or Foe? Exp. Dermatol. 2015, 24, 833–834. [Google Scholar] [CrossRef] [PubMed]
- Crisan, M.; Yap, S.; Casteilla, L.; Chen, C.-W.; Corselli, M.; Park, T.S.; Andriolo, G.; Sun, B.; Zheng, B.; Zhang, L.; et al. A Perivascular Origin for Mesenchymal Stem Cells in Multiple Human Organs. Cell Stem Cell 2008, 3, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Asahara, T.; Murohara, T.; Sullivan, A.; Silver, M.; van der Zee, R.; Li, T.; Witzenbichler, B.; Schatteman, G.; Isner, J.M. Isolation of Putative Progenitor Endothelial Cells for Angiogenesis. Science 1997, 275, 964–966. [Google Scholar] [CrossRef] [PubMed]
- Ceradini, D.J.; Kulkarni, A.R.; Callaghan, M.J.; Tepper, O.M.; Bastidas, N.; Kleinman, M.E.; Capla, J.M.; Galiano, R.D.; Levine, J.P.; Gurtner, G.C. Progenitor Cell Trafficking Is Regulated by Hypoxic Gradients through HIF-1 Induction of SDF-1. Nat. Med. 2004, 10, 858–864. [Google Scholar] [CrossRef] [PubMed]
- Kosaraju, R.; Rennert, R.C.; Maan, Z.N.; Duscher, D.; Barrera, J.; Whittam, A.J.; Januszyk, M.; Rajadas, J.; Rodrigues, M.; Gurtner, G.C. Adipose-Derived Stem Cell-Seeded Hydrogels Increase Endogenous Progenitor Cell Recruitment and Neovascularization in Wounds. Tissue Eng. Part. A 2016, 22, 295–305. [Google Scholar] [CrossRef]
- Werner, S.; Krieg, T.; Smola, H. Keratinocyte–Fibroblast Interactions in Wound Healing. J. Investig. Dermatol. 2007, 127, 998–1008. [Google Scholar] [CrossRef]
- Donati, G.; Rognoni, E.; Hiratsuka, T.; Liakath-Ali, K.; Hoste, E.; Kar, G.; Kayikci, M.; Russell, R.; Kretzschmar, K.; Mulder, K.W.; et al. Wounding Induces Dedifferentiation of Epidermal Gata6+ Cells and Acquisition of Stem Cell Properties. Nat. Cell Biol. 2017, 19, 603–613. [Google Scholar] [CrossRef]
- Blanpain, C.; Horsley, V.; Fuchs, E. Epithelial Stem Cells: Turning over New Leaves. Cell 2007, 128, 445–458. [Google Scholar] [CrossRef]
- Martin, P. Wound Healing--Aiming for Perfect Skin Regeneration. Science 1997, 276, 75–81. [Google Scholar] [CrossRef]
- Freedman, B.R.; Hwang, C.; Talbot, S.; Hibler, B.; Matoori, S.; Mooney, D.J. Breakthrough Treatments for Accelerated Wound Healing. Sci. Adv. 2023, 9, eade7007. [Google Scholar] [CrossRef]
- Gushiken, L.F.S.; Beserra, F.P.; Bastos, J.K.; Jackson, C.J.; Pellizzon, C.H. Cutaneous Wound Healing: An Update from Physiopathology to Current Therapies. Life 2021, 11, 665. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Covington, S.; Sen, C.K.; Januszyk, M.; Kirsner, R.S.; Gurtner, G.C.; Shah, N.H. Rapid Identification of Slow Healing Wounds. Wound Repair Regen. 2016, 24, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Cheung, K.P.; Taylor, K.R.; Jameson, J.M. Immunomodulation at Epithelial Sites by Obesity and Metabolic Disease. Immunol. Res. 2012, 52, 182–199. [Google Scholar] [CrossRef] [PubMed]
- Knoedler, S.; Broichhausen, S.; Guo, R.; Dai, R.; Knoedler, L.; Kauke-Navarro, M.; Diatta, F.; Pomahac, B.; Machens, H.-G.; Jiang, D.; et al. Fibroblasts—The Cellular Choreographers of Wound Healing. Front. Immunol. 2023, 14, 1233800. [Google Scholar] [CrossRef] [PubMed]
- Faour, S.; Farahat, M.; Aijaz, A.; Jeschke, M.G. Fibrosis in Burns: An Overview of Mechanisms and Therapies. Am. J. Physiol.-Cell Physiol. 2023, 325, C1545–C1557. [Google Scholar] [CrossRef] [PubMed]
- Duscher, D.; Maan, Z.N.; Wong, V.W.; Rennert, R.C.; Januszyk, M.; Rodrigues, M.; Hu, M.; Whitmore, A.J.; Whittam, A.J.; Longaker, M.T.; et al. Mechanotransduction and Fibrosis. J. Biomech. 2014, 47, 1997–2005. [Google Scholar] [CrossRef] [PubMed]
- Wong, V.W.; Rustad, K.C.; Akaishi, S.; Sorkin, M.; Glotzbach, J.P.; Januszyk, M.; Nelson, E.R.; Levi, K.; Paterno, J.; Vial, I.N.; et al. Focal Adhesion Kinase Links Mechanical Force to Skin Fibrosis via Inflammatory Signaling. Nat. Med. 2012, 18, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Du, R.; Lei, T. Effects of Autologous Platelet-rich Plasma Injections on Facial Skin Rejuvenation. Exp. Ther. Med. 2020, 19, 3024–3030. [Google Scholar] [CrossRef]
- Kim, D.J.; Iwasaki, A.; Chien, A.L.; Kang, S. UVB-Mediated DNA Damage Induces Matrix Metalloproteinases to Promote Photoaging in an AhR- and SP1-Dependent Manner. J. Clin. Investig. 2022, 7, e156344. [Google Scholar] [CrossRef]
- Everts, P.; Onishi, K.; Jayaram, P.; Lana, J.F.; Mautner, K. Platelet-Rich Plasma: New Performance Understandings and Therapeutic Considerations in 2020. Int. J. Mol. Sci. 2020, 21, 7794. [Google Scholar] [CrossRef]
- Hersant, B.; Sid-Ahmed, M.; Braud, L.; Jourdan, M.; Baba-Amer, Y.; Meningaud, J.-P.; Rodriguez, A.-M. Platelet-Rich Plasma Improves the Wound Healing Potential of Mesenchymal Stem Cells through Paracrine and Metabolism Alterations. Stem Cells Int. 2019, 2019, 1234263. [Google Scholar] [CrossRef] [PubMed]
- Morgan, N.; Abouelgoud, M.; Haiba, D.; Arakeep, H. Comparative Study on the Effect of Injectable Platelet Rich Plasma versus Its Topical Application in the Treatment of Thermal Burn in Adult Male Albino Rat: Histological and Immunohistochemical Study. Egypt. J. Histol. 2021, 45, 125–135. [Google Scholar] [CrossRef]
- Mei-Dan, O.; Laver, L.; Nyska, M.; Mann, G. Platelet Rich Plasma—A New Biotechnology for Treatment of Sports Injuries. Harefuah 2011, 150, 453–457, 490. [Google Scholar] [PubMed]
- Nurden, A. Platelets, Inflammation and Tissue Regeneration. Thromb. Haemost. 2011, 105, S13–S33. [Google Scholar] [CrossRef] [PubMed]
- Marx, R.E.; Carlson, E.R.; Eichstaedt, R.M.; Schimmele, S.R.; Strauss, J.E.; Georgeff, K.R. Platelet-Rich Plasma. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodontol. 1998, 85, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Cole, B.J.; Seroyer, S.T.; Filardo, G.; Bajaj, S.; Fortier, L.A. Platelet-Rich Plasma: Where Are We Now and Where Are We Going? Sports Health A Multidiscip. Approach 2010, 2, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Brass, L. Understanding and Evaluating Platelet Function. Hematology 2010, 2010, 387–396. [Google Scholar] [CrossRef]
- Marx, R.E. Platelet-Rich Plasma (PRP): What Is PRP and What Is Not PRP? Implant. Dent. 2001, 10, 225–228. [Google Scholar] [CrossRef]
- Eppley, B.L.; Woodell, J.E.; Higgins, J. Platelet Quantification and Growth Factor Analysis from Platelet-Rich Plasma: Implications for Wound Healing. Plast. Reconstr. Surg. 2004, 114, 1502–1508. [Google Scholar] [CrossRef]
- Liu, Y.; Kalén, A.; Risto, O.; Wahlström, O. Fibroblast Proliferation Due to Exposure to a Platelet Concentrate in Vitro Is PH Dependent. Wound Repair Regen. 2002, 10, 336–340. [Google Scholar] [CrossRef]
- Carter, C.A.; Jolly, D.G.; Worden, C.E.; Hendren, D.G.; Kane, C.J.M. Platelet-Rich Plasma Gel Promotes Differentiation and Regeneration during Equine Wound Healing. Exp. Mol. Pathol. 2003, 74, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Eppley, B.L.; Pietrzak, W.S.; Blanton, M. Platelet-Rich Plasma: A Review of Biology and Applications in Plastic Surgery. Plast. Reconstr. Surg. 2006, 118, 147e–159e. [Google Scholar] [CrossRef] [PubMed]
- Froum, S.J.; Wallace, S.S.; Tarnow, D.P.; Cho, S.-C. Effect of Platelet-Rich Plasma on Bone Growth and Osseointegration in Human Maxillary Sinus Grafts: Three Bilateral Case Reports. Int. J. Periodontics Restor. Dent. 2002, 22, 45–53. [Google Scholar]
- Pavlovic, V.; Ciric, M.; Jovanovic, V.; Stojanovic, P. Platelet Rich Plasma: A Short Overview of Certain Bioactive Components. Open Med. 2016, 11, 242–247. [Google Scholar] [CrossRef] [PubMed]
- El-Domyati, M.; Abdel-Wahab, H.; Hossam, A. Combining Microneedling with Other Minimally Invasive Procedures for Facial Rejuvenation: A Split-face Comparative Study. Int. J. Dermatol. 2018, 57, 1324–1334. [Google Scholar] [CrossRef]
- Stivala, S.; Sorrentino, S.; Gobbato, S.; Bonetti, N.R.; Camici, G.G.; Lüscher, T.F.; Medalia, O.; Beer, J.H. Glycoprotein Ib clustering in platelets can be inhibited by α-linolenic acid as revealed by cryo-electron tomography. Haematologica 2020, 105, 1660. [Google Scholar] [CrossRef]
- Jain, N.K.; Gulati, M. Platelet-Rich Plasma: A Healing Virtuoso. Blood Res. 2016, 51, 3. [Google Scholar] [CrossRef]
- Marx, R.E. Platelet-Rich Plasma: Evidence to Support Its Use. J. Oral Maxillofac. Surg. 2004, 62, 489–496. [Google Scholar] [CrossRef]
- Senger, D.R.; Galli, S.J.; Dvorak, A.M.; Perruzzi, C.A.; Harvey, V.S.; Dvorak, H.F. Tumor Cells Secrete a Vascular Permeability Factor That Promotes Accumulation of Ascites Fluid. Science 1983, 219, 983–985. [Google Scholar] [CrossRef]
- Herbst, R.S. Review of Epidermal Growth Factor Receptor Biology. Int. J. Radiat. Oncol. Biol. Phys. 2004, 59, S21–S26. [Google Scholar] [CrossRef]
- Josefsson, E.C.; Vainchenker, W.; James, C. Regulation of Platelet Production and Life Span: Role of Bcl-XL and Potential Implications for Human Platelet Diseases. Int. J. Mol. Sci. 2020, 21, 7591. [Google Scholar] [CrossRef] [PubMed]
- Sturm, G.; Monzel, A.S.; Karan, K.R.; Michelson, J.; Ware, S.A.; Cardenas, A.; Lin, J.; Bris, C.; Santhanam, B.; Murphy, M.P.; et al. A Multi-Omics Longitudinal Aging Dataset in Primary Human Fibroblasts with Mitochondrial Perturbations. Sci. Data 2022, 9, 751. [Google Scholar] [CrossRef] [PubMed]
- Manolagas, S.C.; Parfitt, A.M. What Old Means to Bone. Trends Endocrinol. Metab. 2010, 21, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, R.G.; Santos, G.S.; Alkass, N.; Chiesa, T.L.; Azzini, G.O.; da Fonseca, L.F.; dos Santos, A.F.; Rodrigues, B.L.; Mosaner, T.; Lana, J.F. The Regenerative Mechanisms of Platelet-Rich Plasma: A Review. Cytokine 2021, 144, 155560. [Google Scholar] [CrossRef] [PubMed]
- Aslan, J.E. How Can We Use Proteomics to Learn More about Platelets? Platelets 2023, 34, 2217932. [Google Scholar] [CrossRef] [PubMed]
- Bruzek, S.; Betensky, M.; Di Paola, J.; Diacovo, T.; Goldenberg, N.; Ignjatovic, V. What Can the Plasma Proteome Tell Us about Platelets (and Vice Versa)? Platelets 2023, 34, 2186707. [Google Scholar] [CrossRef]
- Bhatlekar, S.; Manne, B.K.; Basak, I.; Edelstein, L.C.; Tugolukova, E.; Stoller, M.L.; Cody, M.J.; Morley, S.C.; Nagalla, S.; Weyrich, A.S.; et al. MiR-125a-5p Regulates Megakaryocyte Proplatelet Formation via the Actin-Bundling Protein L-Plastin. Blood 2020, 136, 1760–1772. [Google Scholar] [CrossRef]
- Caux, M.; Mansour, R.; Xuereb, J.-M.; Chicanne, G.; Viaud, J.; Vauclard, A.; Boal, F.; Payrastre, B.; Tronchère, H.; Severin, S. PIKfyve-Dependent Phosphoinositide Dynamics in Megakaryocyte/Platelet Granule Integrity and Platelet Functions. Arterioscler. Thromb. Vasc. Biol. 2022, 42, 987–1004. [Google Scholar] [CrossRef]
- Demirer, T.; Ilhan, O.; Arat, M.; Genç, Y.; Özcan, M.; Dalva, K.; Çelebi, H.; Beksaç, M.; Akan, H.; Gürman, G.; et al. CD41+ and CD42+ Hematopoietic Progenitor Cells May Predict Platelet Engraftment after Allogeneic Peripheral Blood Stem Cell Transplantation. J. Clin. Apher. 2001, 16, 67–73. [Google Scholar] [CrossRef]
- Machlus, K.R.; Italiano, J.E. Megakaryocyte Development and Platelet Formation. In Platelets; Elsevier: Amsterdam, The Netherlands, 2019; pp. 25–46. [Google Scholar]
- Wang, J.; Xie, J.; Wang, D.; Han, X.; Chen, M.; Shi, G.; Jiang, L.; Zhao, M. CXCR4high Megakaryocytes Regulate Host-Defense Immunity against Bacterial Pathogens. Elife 2022, 11, e78662. [Google Scholar] [CrossRef]
- Thon, J.N.; Italiano, J.E. Platelets: Production, Morphology and Ultrastructure. In Antiplatelet Agents; Springer: Berlin/Heidelberg, Germany, 2012; pp. 3–22. [Google Scholar]
- Cramer, E.M.; Norol, F.; Guichard, J.; Breton-Gorius, J.; Vainchenker, W.; Massé, J.-M.; Debili, N. Ultrastructure of Platelet Formation by Human Megakaryocytes Cultured With the Mpl Ligand. Blood 1997, 89, 2336–2346. [Google Scholar] [CrossRef] [PubMed]
- Yeung, A.K.; Villacorta-Martin, C.; Hon, S.; Rock, J.R.; Murphy, G.J. Lung Megakaryocytes Display Distinct Transcriptional and Phenotypic Properties. Blood Adv. 2020, 4, 6204–6217. [Google Scholar] [CrossRef] [PubMed]
- Livada, A.C.; Pariser, D.N.; Morrell, C.N. Megakaryocytes in the Lung: History and Future Perspectives. Res. Pract. Thromb. Haemost. 2023, 7, 100053. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Alibhai, D.; Walsh, T.G.; Tarassova, N.; Englert, M.; Birol, S.Z.; Li, Y.; Williams, C.M.; Neal, C.R.; Burkard, P.; et al. Highly Efficient Platelet Generation in Lung Vasculature Reproduced by Microfluidics. Nat. Commun. 2023, 14, 4026. [Google Scholar] [CrossRef] [PubMed]
- Noetzli, L.J.; French, S.L.; Machlus, K.R. New Insights Into the Differentiation of Megakaryocytes From Hematopoietic Progenitors. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1288–1300. [Google Scholar] [CrossRef] [PubMed]
- Machlus, K.R.; Italiano, J.E. The Incredible Journey: From Megakaryocyte Development to Platelet Formation. J. Cell Biol. 2013, 201, 785–796. [Google Scholar] [CrossRef]
- Vainchenker, W.; Raslova, H. Megakaryocyte Polyploidization: Role in Platelet Production. Platelets 2020, 31, 707–716. [Google Scholar] [CrossRef]
- Guo, T.; Wang, X.; Qu, Y.; Yin, Y.; Jing, T.; Zhang, Q. Megakaryopoiesis and Platelet Production: Insight into Hematopoietic Stem Cell Proliferation and Differentiation. Stem Cell Investig. 2015, 2, 3. [Google Scholar] [CrossRef]
- Mazzi, S.; Lordier, L.; Debili, N.; Raslova, H.; Vainchenker, W. Megakaryocyte and Polyploidization. Exp. Hematol. 2018, 57, 1–13. [Google Scholar] [CrossRef]
- Lefrançais, E.; Ortiz-Muñoz, G.; Caudrillier, A.; Mallavia, B.; Liu, F.; Sayah, D.M.; Thornton, E.E.; Headley, M.B.; David, T.; Coughlin, S.R.; et al. The Lung Is a Site of Platelet Biogenesis and a Reservoir for Haematopoietic Progenitors. Nature 2017, 544, 105–109. [Google Scholar] [CrossRef]
- Schulze, H.; Korpal, M.; Hurov, J.; Kim, S.-W.; Zhang, J.; Cantley, L.C.; Graf, T.; Shivdasani, R.A. Characterization of the Megakaryocyte Demarcation Membrane System and Its Role in Thrombopoiesis. Blood 2006, 107, 3868–3875. [Google Scholar] [CrossRef] [PubMed]
- Kowata, S.; Isogai, S.; Murai, K.; Ito, S.; Tohyama, K.; Hitomi, J.; Ishida, Y. Novel Concept of Platelet Production from Megakaryocyte in Intact Bone Marrow: Proplatelet and Thick Protrusion. Blood 2012, 120, 2300. [Google Scholar] [CrossRef]
- Heijnen, H.F.G.; Korporaal, S.J.A. Platelet Morphology and Ultrastructure. In Platelets in Thrombotic and Non-Thrombotic Disorders; Springer International Publishing: Cham, Switzerland, 2017; pp. 21–37. [Google Scholar]
- Bender, M.; Thon, J.N.; Ehrlicher, A.J.; Wu, S.; Mazutis, L.; Deschmann, E.; Sola-Visner, M.; Italiano, J.E.; Hartwig, J.H. Microtubule Sliding Drives Proplatelet Elongation and Is Dependent on Cytoplasmic Dynein. Blood 2015, 125, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Becker, I.C.; Scheller, I.; Wackerbarth, L.M.; Beck, S.; Heib, T.; Aurbach, K.; Manukjan, G.; Gross, C.; Spindler, M.; Nagy, Z.; et al. Actin/Microtubule Crosstalk during Platelet Biogenesis in Mice Is Critically Regulated by Twinfilin1 and Cofilin1. Blood Adv. 2020, 4, 2124–2134. [Google Scholar] [CrossRef] [PubMed]
- Boscher, J.; Guinard, I.; Eckly, A.; Lanza, F.; Léon, C. Blood Platelet Formation at a Glance. J. Cell Sci. 2020, 133. [Google Scholar] [CrossRef] [PubMed]
- Thon, J.N.; Montalvo, A.; Patel-Hett, S.; Devine, M.T.; Richardson, J.L.; Ehrlicher, A.; Larson, M.K.; Hoffmeister, K.; Hartwig, J.H.; Italiano, J.E. Cytoskeletal Mechanics of Proplatelet Maturation and Platelet Release. J. Cell Biol. 2010, 191, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Go, S.; Jeong, D.; Chung, J.; Kim, G.; Song, J.; Moon, E.; Huh, Y.H.; Kim, D. Super-Resolution Imaging Reveals Cytoskeleton-Dependent Organelle Rearrangement within Platelets at Intermediate Stages of Maturation. Structure 2021, 29, 810–822.e3. [Google Scholar] [CrossRef]
- Kemble, S.; Dalby, A.; Lowe, G.C.; Nicolson, P.L.R.; Watson, S.P.; Senis, Y.; Thomas, S.G.; Harrison, P. Analysis of Preplatelets and Their Barbell Platelet Derivatives by Imaging Flow Cytometry. Blood Adv. 2022, 6, 2932–2946. [Google Scholar] [CrossRef]
- Yadav, S.; Storrie, B. The Cellular Basis of Platelet Secretion: Emerging Structure/Function Relationships. Platelets 2017, 28, 108–118. [Google Scholar] [CrossRef]
- Yadav, S.; Williamson, J.K.; Aronova, M.A.; Prince, A.A.; Pokrovskaya, I.D.; Leapman, R.D.; Storrie, B. Golgi Proteins in Circulating Human Platelets Are Distributed across Non-Stacked, Scattered Structures. Platelets 2017, 28, 400–408. [Google Scholar] [CrossRef]
- Melchinger, H.; Jain, K.; Tyagi, T.; Hwa, J. Role of Platelet Mitochondria: Life in a Nucleus-Free Zone. Front. Cardiovasc. Med. 2019, 6, 153. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, P.K.; Kim, S.; Kim, S. Shedding Light on the Cell Biology of Platelet-Derived Extracellular Vesicles and Their Biomedical Applications. Life 2023, 13, 1403. [Google Scholar] [CrossRef] [PubMed]
- Soslau, G. Platelet Protein Synthesis, Regulation, and Post-Translational Modifications: Mechanics and Function. Crit. Rev. Biochem. Mol. Biol. 2023, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Weyrich, A.S.; Schwertz, H.; Kraiss, L.W.; Zimmerman, G.A. Protein Synthesis by Platelets: Historical and New Perspectives. J. Thromb. Haemost. 2009, 7, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Hur, W.S.; Juang, L.J.; Mazinani, N.; Munro, L.; Jefferies, W.A.; Kastrup, C.J. Post-Translational Modifications of Platelet-Derived Amyloid Precursor Protein by Coagulation Factor XIII-A*. Biochemistry 2020, 59, 4449–4455. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, P.; Solari, F.A.; Sickmann, A.; Garcia, A.; Jurk, K.; Heemskerk, J.W.M. Molecular Proteomics and Signalling of Human Platelets in Health and Disease. Int. J. Mol. Sci. 2021, 22, 9860. [Google Scholar] [CrossRef]
- Zeiler, M.; Moser, M.; Mann, M. Copy Number Analysis of the Murine Platelet Proteome Spanning the Complete Abundance Range. Mol. Cell. Proteom. 2014, 13, 3435–3445. [Google Scholar] [CrossRef]
- Burkhart, J.M.; Gambaryan, S.; Watson, S.P.; Jurk, K.; Walter, U.; Sickmann, A.; Heemskerk, J.W.M.; Zahedi, R.P. What Can Proteomics Tell Us About Platelets? Circ. Res. 2014, 114, 1204–1219. [Google Scholar] [CrossRef]
- Zellner, M. How Can Platelet Proteomics Best Be Used to Interrogate Disease? Platelets 2023, 34, 2220046. [Google Scholar] [CrossRef]
- Potts, K.S.; Farley, A.; Dawson, C.A.; Rimes, J.; Biben, C.; de Graaf, C.; Potts, M.A.; Stonehouse, O.J.; Carmagnac, A.; Gangatirkar, P.; et al. Membrane Budding Is a Major Mechanism of in Vivo Platelet Biogenesis. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef]
- Schulze, H.; Stegner, D. Imaging Platelet Biogenesis in Vivo. Res. Pract. Thromb. Haemost. 2018, 2, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Sim, X.; Poncz, M.; Gadue, P.; French, D.L. Understanding Platelet Generation from Megakaryocytes: Implications for in Vitro–Derived Platelets. Blood 2016, 127, 1227–1233. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, N.; Eto, K. Generation and Manipulation of Human IPSC-Derived Platelets. Cell. Mol. Life Sci. 2021, 78, 3385–3401. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Eguren, A.; Gómez-Álvarez, M.; Francés-Herrero, E.; Romeu, M.; Ferrero, H.; Seli, E.; Cervelló, I. Human Umbilical Cord-Based Therapeutics: Stem Cells and Blood Derivatives for Female Reproductive Medicine. Int. J. Mol. Sci. 2022, 23, 15942. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Wang, L.; Wang, H.; Wang, H.; Dai, W.; Jiang, Y. Good Manufacturing Practice-Grade of Megakaryocytes Produced by a Novel Ex Vivo Culturing Platform. Clin. Transl. Sci. 2020, 13, 1115–1126. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zheng, J. Platelet Generation in Vivo and in Vitro. Springerplus 2016, 5, 787. [Google Scholar] [CrossRef]
- Cuenca-Zamora, E.J.; Ferrer-Marín, F.; Rivera, J.; Teruel-Montoya, R. Tubulin in Platelets: When the Shape Matters. Int. J. Mol. Sci. 2019, 20, 3484. [Google Scholar] [CrossRef]
- Italiano, J.E.; Lecine, P.; Shivdasani, R.A.; Hartwig, J.H. Blood Platelets Are Assembled Principally at the Ends of Proplatelet Processes Produced by Differentiated Megakaryocytes. J. Cell Biol. 1999, 147, 1299–1312. [Google Scholar] [CrossRef]
- De Simone, I.; Baaten, C.C.F.M.J.; Gibbins, J.M.; Ten Cate, H.; Heemskerk, J.W.M.; Jones, C.I.; van der Meijden, P.E.J. Repeated Platelet Activation and the Potential of Previously Activated Platelets to Contribute to Thrombus Formation. J. Thromb. Haemost. 2023, 21, 1289–1306. [Google Scholar] [CrossRef]
- Tang, L.; Liu, C.; Rosenberger, P. Platelet Formation and Activation Are Influenced by Neuronal Guidance Proteins. Front. Immunol. 2023, 14, 1206906. [Google Scholar] [CrossRef]
- Muravlev, I.A.; Dobrovolsky, A.B.; Antonova, O.A.; Khaspekova, S.G.; Mazurov, A.V. Effects of Platelets Activated by Different Agonists on Fibrin Formation and Thrombin Generation. Platelets 2023, 34, 2139365. [Google Scholar] [CrossRef] [PubMed]
- Bornert, A.; Boscher, J.; Pertuy, F.; Eckly, A.; Stegner, D.; Strassel, C.; Gachet, C.; Lanza, F.; Léon, C. Cytoskeletal-based mechanisms differently regulate in vivo and in vitro proplatelet formation. Haematologica 2020, 106, 1368–1380. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, J.; Bompard, G.; Cau, J.; Kunishima, S.; Rabeharivelo, G.; Mateos-Langerak, J.; Cazevieille, C.; Cavelier, P.; Boizet-Bonhoure, B.; Delsert, C.; et al. Microtubule Polyglutamylation and Acetylation Drive Microtubule Dynamics Critical for Platelet Formation. BMC Biol. 2018, 16, 116. [Google Scholar] [CrossRef] [PubMed]
- Zelená, A.; Blumberg, J.; Probst, D.; Gerasimaitė, R.; Lukinavičius, G.; Schwarz, U.S.; Köster, S. Force Generation in Human Blood Platelets by Filamentous Actomyosin Structures. Biophys. J. 2023, 122, 3340–3353. [Google Scholar] [CrossRef] [PubMed]
- Durán-Saenz, N.Z.; Serrano-Puente, A.; Gallegos-Flores, P.I.; Mendoza-Almanza, B.D.; Esparza-Ibarra, E.L.; Godina-González, S.; González-Curiel, I.E.; Ayala-Luján, J.L.; Hernández-Barrales, M.; Cueto-Villalobos, C.F.; et al. Platelet Membrane: An Outstanding Factor in Cancer Metastasis. Membranes 2022, 12, 182. [Google Scholar] [CrossRef] [PubMed]
- Protty, M.B.; Jenkins, P.V.; Collins, P.W.; O’Donnell, V.B. The Role of Procoagulant Phospholipids on the Surface of Circulating Blood Cells in Thrombosis and Haemostasis. Open Biol. 2022, 12, 210318. [Google Scholar] [CrossRef] [PubMed]
- Sakuragi, T.; Nagata, S. Regulation of Phospholipid Distribution in the Lipid Bilayer by Flippases and Scramblases. Nat. Rev. Mol. Cell Biol. 2023, 24, 576–596. [Google Scholar] [CrossRef]
- Pollet, H.; Conrard, L.; Cloos, A.-S.; Tyteca, D. Plasma Membrane Lipid Domains as Platforms for Vesicle Biogenesis and Shedding? Biomolecules 2018, 8, 94. [Google Scholar] [CrossRef]
- Lorent, J.H.; Levental, K.R.; Ganesan, L.; Rivera-Longsworth, G.; Sezgin, E.; Doktorova, M.; Lyman, E.; Levental, I. Author Correction: Plasma Membranes Are Asymmetric in Lipid Unsaturation, Packing and Protein Shape. Nat. Chem. Biol. 2020, 16, 710. [Google Scholar] [CrossRef]
- Lagoutte-Renosi, J.; Allemand, F.; Ramseyer, C.; Rabani, V.; Davani, S. Influence of Antiplatelet Agents on the Lipid Composition of Platelet Plasma Membrane: A Lipidomics Approach with Ticagrelor and Its Active Metabolite. Int. J. Mol. Sci. 2021, 22, 1432. [Google Scholar] [CrossRef]
- Nenna, A.; Nappi, F.; Lusini, M.; Satriano, U.M.; Schilirò, D.; Spadaccio, C.; Chello, M. Effect of Statins on Platelet Activation and Function: From Molecular Pathways to Clinical Effects. Biomed. Res. Int. 2021, 2021, 6661847. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.G. The Structure of Resting and Activated Platelets. In Platelets; Elsevier: Amsterdam, The Netherlands, 2019; pp. 47–77. [Google Scholar]
- Offermanns, S. Activation of Platelet Function Through G Protein–Coupled Receptors. Circ. Res. 2006, 99, 1293–1304. [Google Scholar] [CrossRef] [PubMed]
- Patel-Hett, S.; Wang, H.; Begonja, A.J.; Thon, J.N.; Alden, E.C.; Wandersee, N.J.; An, X.; Mohandas, N.; Hartwig, J.H.; Italiano, J.E. The Spectrin-Based Membrane Skeleton Stabilizes Mouse Megakaryocyte Membrane Systems and Is Essential for Proplatelet and Platelet Formation. Blood 2011, 118, 1641–1652. [Google Scholar] [CrossRef] [PubMed]
- Hanke, J.; Ranke, C.; Perego, E.; Köster, S. Human Blood Platelets Contract in Perpendicular Direction to Shear Flow. Soft Matter 2019, 15, 2009–2019. [Google Scholar] [CrossRef] [PubMed]
- Behnke, O. The Formation of Fusiform Proplatelets and Their Transformation to Discoid Platelets. Platelets 1993, 4, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, Y.; Fitch-Tewfik, J.L.; Qiu, Y.; Ahn, B.; Myers, D.R.; Tran, R.; Fay, M.E.; Ding, L.; Spearman, P.W.; Michelson, A.D.; et al. Platelet Geometry Sensing Spatially Regulates α-Granule Secretion to Enable Matrix Self-Deposition. Blood 2015, 126, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Stalker, T.J.; Welsh, J.D.; Brass, L.F. Shaping the Platelet Response to Vascular Injury. Curr. Opin. Hematol. 2014, 21, 410–417. [Google Scholar] [CrossRef]
- O’Sullivan, L.R.; Cahill, M.R.; Young, P.W. The Importance of Alpha-Actinin Proteins in Platelet Formation and Function, and Their Causative Role in Congenital Macrothrombocytopenia. Int. J. Mol. Sci. 2021, 22, 9363. [Google Scholar] [CrossRef]
- Woronowicz, K.; Dilks, J.R.; Rozenvayn, N.; Dowal, L.; Blair, P.S.; Peters, C.G.; Woronowicz, L.; Flaumenhaft, R. The Platelet Actin Cytoskeleton Associates with SNAREs and Participates in α-Granule Secretion. Biochemistry 2010, 49, 4533–4542. [Google Scholar] [CrossRef]
- Golebiewska, E.M.; Poole, A.W. Secrets of Platelet Exocytosis—What Do We Really Know about Platelet Secretion Mechanisms? Br. J. Haematol. 2014, 165, 204–216. [Google Scholar] [CrossRef]
- Ménasché, G.; Longé, C.; Bratti, M.; Blank, U. Cytoskeletal Transport, Reorganization, and Fusion Regulation in Mast Cell-Stimulus Secretion Coupling. Front. Cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.L.; Meneses-Salas, E.; Ramadass, M.; Monfregola, J.; Rahman, F.; Carvalho Gontijo, R.; Kiosses, W.B.; Pestonjamasp, K.; Allen, D.; Zhang, J.; et al. Differential Dysregulation of Granule Subsets in WASH-Deficient Neutrophil Leukocytes Resulting in Inflammation. Nat. Commun. 2022, 13, 5529. [Google Scholar] [CrossRef] [PubMed]
- Ebstrup, M.L.; Dias, C.; Heitmann, A.S.B.; Sønder, S.L.; Nylandsted, J. Actin Cytoskeletal Dynamics in Single-Cell Wound Repair. Int. J. Mol. Sci. 2021, 22, 10886. [Google Scholar] [CrossRef]
- White, J.G.; Clawson, C.C. The Surface-Connected Canalicular System of Blood Platelets--a Fenestrated Membrane System. Am. J. Pathol. 1980, 101, 353–364. [Google Scholar] [PubMed]
- Pokrovskaya, I.D.; Tobin, M.; Desai, R.; Joshi, S.; Kamykowski, J.A.; Zhang, G.; Aronova, M.A.; Whiteheart, S.W.; Leapman, R.D.; Storrie, B. Canalicular System Reorganization during Mouse Platelet Activation as Revealed by 3D Ultrastructural Analysis. Platelets 2021, 32, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Pokrovskaya, I.D.; Aronova, M.A.; Kamykowski, J.A.; Prince, A.A.; Hoyne, J.D.; Calco, G.N.; Kuo, B.C.; He, Q.; Leapman, R.D.; Storrie, B. STEM Tomography Reveals That the Canalicular System and A-granules Remain Separate Compartments during Early Secretion Stages in Blood Platelets. J. Thromb. Haemost. 2016, 14, 572–584. [Google Scholar] [CrossRef]
- Neumüller, J.; Ellinger, A.; Wagner, T. Transmission Electron Microscopy of Platelets FROM Apheresis and Buffy-Coat-Derived Platelet Concentrates. In The Transmission Electron Microscope—Theory and Applications; InTech: Sydney, Australia, 2015. [Google Scholar]
- Gruba, S.M.; Koseoglu, S.; Meyer, A.F.; Meyer, B.M.; Maurer-Jones, M.A.; Haynes, C.L. Platelet Membrane Variations and Their Effects on δ-Granule Secretion Kinetics and Aggregation Spreading among Different Species. Biochim. Biophys. Acta (BBA)-Biomembr. 2015, 1848, 1609–1618. [Google Scholar] [CrossRef]
- Escolar, G.; Leistikow, E.; White, J. The Fate of the Open Canalicular System in Surface and Suspension- Activated Platelets. Blood 1989, 74, 1983–1988. [Google Scholar] [CrossRef]
- Eckly, A.; Heijnen, H.; Pertuy, F.; Geerts, W.; Proamer, F.; Rinckel, J.-Y.; Léon, C.; Lanza, F.; Gachet, C. Biogenesis of the Demarcation Membrane System (DMS) in Megakaryocytes. Blood 2014, 123, 921–930. [Google Scholar] [CrossRef]
- Selvadurai, M.V.; Hamilton, J.R. Structure and Function of the Open Canalicular System—The Platelet’s Specialized Internal Membrane Network. Platelets 2018, 29, 319–325. [Google Scholar] [CrossRef]
- Osman, S.; Dalmay, D.; Mahaut-Smith, M. Fluorescence Approaches to Image and Quantify the Demarcation Membrane System in Living Megakaryocytes. In Platelets and Megakaryocytes; Springer: Berlin/Heidelberg, Germany, 2018; pp. 195–215. [Google Scholar]
- Pokrovskaya, I.D.; Yadav, S.; Rao, A.; McBride, E.; Kamykowski, J.A.; Zhang, G.; Aronova, M.A.; Leapman, R.D.; Storrie, B. 3D Ultrastructural Analysis of A-granule, Dense Granule, Mitochondria, and Canalicular System Arrangement in Resting Human Platelets. Res. Pract. Thromb. Haemost. 2020, 4, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Noetzli, L.J.; Italiano, J.E. Unlocking the Molecular Secrete(s) of α-Granule Biogenesis. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 2539–2541. [Google Scholar] [CrossRef] [PubMed]
- McBride, E.L.; Rao, A.; Zhang, G.; Hoyne, J.D.; Calco, G.N.; Kuo, B.C.; He, Q.; Prince, A.A.; Pokrovskaya, I.D.; Storrie, B.; et al. Comparison of 3D Cellular Imaging Techniques Based on Scanned Electron Probes: Serial Block Face SEM vs. Axial Bright-Field STEM Tomography. J. Struct. Biol. 2018, 202, 216–228. [Google Scholar] [CrossRef] [PubMed]
- Jonnalagadda, D.; Izu, L.T.; Whiteheart, S.W. Platelet Secretion Is Kinetically Heterogeneous in an Agonist-Responsive Manner. Blood 2012, 120, 5209–5216. [Google Scholar] [CrossRef] [PubMed]
- Starlinger, P.; Haegele, S.; Offensperger, F.; Oehlberger, L.; Pereyra, D.; Kral, J.B.; Schrottmaier, W.C.; Badrnya, S.; Reiberger, T.; Ferlitsch, A.; et al. The Profile of Platelet A-granule Released Molecules Affects Postoperative Liver Regeneration. Hepatology 2016, 63, 1675–1688. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.; Whiteheart, S.W. The Nuts and Bolts of the Platelet Release Reaction. Platelets 2017, 28, 129–137. [Google Scholar] [CrossRef]
- Maynard, D.M.; Heijnen, H.F.G.; Gahl, W.A.; Gunay-Aygun, M. The A-granule Proteome: Novel Proteins in Normal and Ghost Granules in Gray Platelet Syndrome. J. Thromb. Haemost. 2010, 8, 1786–1796. [Google Scholar] [CrossRef]
- Maynard, D.M.; Heijnen, H.F.G.; Horne, M.K.; White, J.G.; Gahl, W.A. Proteomic Analysis of Platelet A-granules Using Mass Spectrometry. J. Thromb. Haemost. 2007, 5, 1945–1955. [Google Scholar] [CrossRef]
- Mosher, D.F.; Pesciotta, D.M.; Loftus, J.C.; Albrecht, R.M. Secreted Alpha Granule Proteins. In Platelet Membrane Glycoproteins; Springer US: Boston, MA, USA, 1985; pp. 171–191. [Google Scholar]
- Woods, V.M.A.; Latorre-Rey, L.J.; Schenk, F.; Rommel, M.G.E.; Moritz, T.; Modlich, U. Targeting Transgenic Proteins to Alpha Granules for Platelet-Directed Gene Therapy. Mol. Ther. Nucleic Acids 2022, 27, 774–786. [Google Scholar] [CrossRef]
- Berger, G.; Masse, J.; Cramer, E. Alpha-Granule Membrane Mirrors the Platelet Plasma Membrane and Contains the Glycoproteins Ib, IX, and V. Blood 1996, 87, 1385–1395. [Google Scholar] [CrossRef]
- Blair, P.; Flaumenhaft, R. Platelet α-Granules: Basic Biology and Clinical Correlates. Blood Rev. 2009, 23, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Abrams, C.S. The Yin-Yang of Platelet Granules. Blood 2008, 111, 979. [Google Scholar] [CrossRef]
- Italiano, J.E.; Richardson, J.L.; Patel-Hett, S.; Battinelli, E.; Zaslavsky, A.; Short, S.; Ryeom, S.; Folkman, J.; Klement, G.L. Angiogenesis Is Regulated by a Novel Mechanism: Pro- and Antiangiogenic Proteins Are Organized into Separate Platelet α Granules and Differentially Released. Blood 2008, 111, 1227–1233. [Google Scholar] [CrossRef] [PubMed]
- Everts, P.A.; Lana, J.F.; Onishi, K.; Buford, D.; Peng, J.; Mahmood, A.; Fonseca, L.F.; van Zundert, A.; Podesta, L. Angiogenesis and Tissue Repair Depend on Platelet Dosing and Bioformulation Strategies Following Orthobiological Platelet-Rich Plasma Procedures: A Narrative Review. Biomedicines 2023, 11, 1922. [Google Scholar] [CrossRef] [PubMed]
- Da Costa Marques, R.; Simon, J.; d’Arros, C.; Landfester, K.; Jurk, K.; Mailänder, V. Proteomics Reveals Differential Adsorption of Angiogenic Platelet Lysate Proteins on Calcium Phosphate Bone Substitute Materials. Regen. Biomater. 2022, 9, rbac044. [Google Scholar] [CrossRef] [PubMed]
- Beirat, A.F.; Menakuru, S.R.; Kalra, M. Platelet Delta (δ)-Storage Pool Deficiency: A Case Series and Review of the Literature. Hematol. Rep. 2023, 15, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Gunning, W.T.; Raghavan, M.; Calomeni, E.P.; Turner, J.N.; Roysam, B.; Roysam, S.; Smith, M.R.; Kouides, P.A.; Lachant, N.A. A Morphometric Analysis of Platelet Dense Granules of Patients with Unexplained Bleeding: A New Entity of Delta-Microgranular Storage Pool Deficiency. J. Clin. Med. 2020, 9, 1734. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yuan, Y.; Li, W. Sorting Machineries: How Platelet-Dense Granules Differ from α-Granules. Biosci. Rep. 2018, 38, BSR20180458. [Google Scholar] [CrossRef]
- Flaumenhaft, R.; Sharda, A. Platelet Secretion. In Platelets; Elsevier: Amsterdam, The Netherlands, 2019; pp. 349–370. [Google Scholar]
- Bai, M.-J.; Wang, J.-L.; Elsheikha, H.M.; Liang, Q.-L.; Chen, K.; Nie, L.-B.; Zhu, X.-Q. Functional Characterization of Dense Granule Proteins in Toxoplasma gondii RH Strain Using CRISPR-Cas9 System. Front. Cell Infect. Microbiol. 2018, 8, 300. [Google Scholar] [CrossRef]
- Griffith, M.B.; Pearce, C.S.; Heaslip, A.T. Dense Granule Biogenesis, Secretion, and Function in Toxoplasma gondii. J. Eukaryot. Microbiol. 2022, 69, e12904. [Google Scholar] [CrossRef]
- Pan, M.; Li, M.; Li, L.; Song, Y.; Hou, L.; Zhao, J.; Shen, B. Identification of Novel Dense-Granule Proteins in Toxoplasma gondii by Two Proximity-Based Biotinylation Approaches. J. Proteome Res. 2018, 18, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Iyer, J.; Badin, M.; Graf, L.; Rivard, G.E.; Paterson, A.D.; Pare, G.; Hayward, C.P.M. Bleeding Risks Associated with Confirmed Platelet Dense Granule Deficiency and/or Impaired Aggregation Responses. Blood 2016, 128, 3728. [Google Scholar] [CrossRef]
- Maatouk, A.F.; Brockington, J.; Gudena, V. A Case of Platelet Storage Pool Deficiency of Dense Granules (Delta SPD). Am. J. Case Rep. 2011, 12, 23–26. [Google Scholar] [CrossRef]
- Kim, J.G.; Leem, Y.-E.; Kwon, I.; Kang, J.-S.; Bae, Y.M.; Cho, H. Estrogen Modulates Serotonin Effects on Vasoconstriction through Src Inhibition. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Konradt, C.; Corken, A.; Ware, J.; Nieswandt, B.; Di Paola, J.; Yu, M.; Wang, D.; Nieman, M.T.; Whiteheart, S.W.; et al. Hemostasis vs. Homeostasis: Platelets Are Essential for Preserving Vascular Barrier Function in the Absence of Injury or Inflammation. Proc. Natl. Acad. Sci. USA 2020, 117, 24316–24325. [Google Scholar] [CrossRef] [PubMed]
- Ho-Tin-Noé, B.; Le Chapelain, O.; Camerer, E. Platelets Maintain Vascular Barrier Function in the Absence of Injury or Inflammation. J. Thromb. Haemost. 2021, 19, 1145–1148. [Google Scholar] [CrossRef] [PubMed]
- Golebiewska, E.M.; Poole, A.W. Platelet Secretion: From Haemostasis to Wound Healing and Beyond. Blood Rev. 2015, 29, 153–162. [Google Scholar] [CrossRef]
- Koupenova, M.; Clancy, L.; Corkrey, H.A.; Freedman, J.E. Circulating Platelets as Mediators of Immunity, Inflammation, and Thrombosis. Circ. Res. 2018, 122, 337–351. [Google Scholar] [CrossRef]
- Polasek, J. Platelet Secretory Granules or Secretory Lysosomes? Platelets 2005, 16, 500–501. [Google Scholar] [CrossRef]
- Monis, B.; Wasserkrug, H. Histochemistry of Glycosidases of Megakaryocytes and Platelets. Histochemie 1967, 10, 362–368. [Google Scholar] [CrossRef]
- Badimón, L.; Vilahur, G.; Padró, T. Lipoproteins, Platelets, and Atherothrombosis. Rev. Española Cardiol. Engl. Ed. 2009, 62, 1161–1178. [Google Scholar] [CrossRef]
- Pokrovskaya, I.D.; Joshi, S.; Tobin, M.; Desai, R.; Aronova, M.A.; Kamykowski, J.A.; Zhang, G.; Whiteheart, S.W.; Leapman, R.D.; Storrie, B. SNARE-Dependent Membrane Fusion Initiates α-Granule Matrix Decondensation in Mouse Platelets. Blood Adv. 2018, 2, 2947–2958. [Google Scholar] [CrossRef] [PubMed]
- Flaumenhaft, R. Molecular Basis of Platelet Granule Secretion. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1152–1160. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Ma, J.; Zhu, D.; Wang, Z.; Li, Y.; He, X.; Zhang, G.; Kang, X. The Role of S100A6 in Human Diseases: Molecular Mechanisms and Therapeutic Potential. Biomolecules 2023, 13, 1139. [Google Scholar] [CrossRef] [PubMed]
- Sanada, C.; Xavier-Ferrucio, J.; Lu, Y.-C.; Min, E.; Zhang, P.-X.; Zou, S.; Kang, E.; Zhang, M.; Zerafati, G.; Gallagher, P.G.; et al. Adult Human Megakaryocyte-Erythroid Progenitors Are in the CD34+CD38mid Fraction. Blood 2016, 128, 923–933. [Google Scholar] [CrossRef] [PubMed]
- Muench, M.O.; Bárcena, A. Megakaryocyte Growth and Development Factor Is a Potent Growth Factor for Primitive Hematopoietic Progenitors in the Human Fetus. Pediatr. Res. 2004, 55, 1050–1056. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sawai, N.; Koike, K.; Higuchi, T.; Ogami, K.; Oda, M. Thrombopoietin Enhances the Production of Myeloid Cells, but Not Megakaryocytes, in Juvenile Chronic Myelogenous Leukemia. Blood 1998, 91, 4065–4073. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, C.S.; Charó, N.; Tatti, S.; Gómez, R.M.; D’Atri, L.P.; Schattner, M. Regulation of Megakaryo/Thrombopoiesis by Endosomal Toll-like Receptor 7 and 8 Activation of CD34+ Cells in a Viral Infection Model. Res. Pract. Thromb. Haemost. 2023, 7, 100184. [Google Scholar] [CrossRef]
- Frydman, G.H.; Ellett, F.; Jorgensen, J.; Marand, A.L.; Zukerberg, L.; Selig, M.K.; Tessier, S.N.; Wong, K.H.K.; Olaleye, D.; Vanderburg, C.R.; et al. Megakaryocytes Respond during Sepsis and Display Innate Immune Cell Behaviors. Front. Immunol. 2023, 14, 1083339. [Google Scholar] [CrossRef]
- Huang, H.; Cantor, A.B. Common Features of Megakaryocytes and Hematopoietic Stem Cells: What’s the Connection? J. Cell Biochem. 2009, 107, 857–864. [Google Scholar] [CrossRef]
- Bendas, G.; Schlesinger, M. The GPIb-IX Complex on Platelets: Insight into Its Novel Physiological Functions Affecting Immune Surveillance, Hepatic Thrombopoietin Generation, Platelet Clearance and Its Relevance for Cancer Development and Metastasis. Exp. Hematol. Oncol. 2022, 11, 19. [Google Scholar] [CrossRef] [PubMed]
- Mazzeffi, M.; Tanaka, K.; Wu, Y.-F.; Zhang, A.; Kareddy, N.; Tadjou Tito, E.; Rock, P.; Michelson, A.D.; Frelinger, A.L. Platelet Surface GPIbα, Activated GPIIb-IIIa, and P-Selectin Levels in Adult Veno-Arterial Extracorporeal Membrane Oxygenation Patients. Platelets 2022, 33, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, O.; Sánchez-Madrid, F. Molecular Basis of Leukocyte–Endothelium Interactions during the Inflammatory Response. Rev. Española Cardiol. Engl. Ed. 2009, 62, 552–562. [Google Scholar] [CrossRef]
- Hamilos, M.; Petousis, S.; Parthenakis, F. Interaction between Platelets and Endothelium: From Pathophysiology to New Therapeutic Options. Cardiovasc. Diagn. Ther. 2018, 8, 568–580. [Google Scholar] [CrossRef] [PubMed]
- Nishijima, K.; Kiryu, J.; Tsujikawa, A.; Miyamoto, K.; Honjo, M.; Tanihara, H.; Nonaka, A.; Yamashiro, K.; Katsuta, H.; Miyahara, S.; et al. Platelets Adhering to the Vascular Wall Mediate Postischemic Leukocyte–Endothelial Cell Interactions in Retinal Microcirculation. Investig. Opthalmology Vis. Sci. 2004, 45, 977. [Google Scholar] [CrossRef] [PubMed]
- Sachs, U.J.H. Interaction of Platelets, Leukocytes and the Endothelium*. Transfus. Med. Hemotherapy 2006, 33, 169–176. [Google Scholar] [CrossRef]
- Neubauer, K.; Zieger, B. Endothelial Cells and Coagulation. Cell Tissue Res. 2022, 387, 391–398. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Oswald, B.E.; Sullivan, J.A.; Dahmani, F.Z.; Pasman, Y.; Liu, Z.; Chen, P.; Ni, H. Platelet Physiology and Immunology: Pathogenesis and Treatment of Classical and Non-Classical Fetal and Neonatal Alloimmune Thrombocytopenia. Ann. Blood 2019, 4, 29. [Google Scholar] [CrossRef]
- Amalia, L. The Role of Platelet-Selectin as a Marker of Thrombocyte Aggregation on Cerebral Sinus Venous Thrombosis. J. Blood Med. 2022, 13, 267–274. [Google Scholar] [CrossRef]
- Fabricius, H.-Å.; Starzonek, S.; Lange, T. The Role of Platelet Cell Surface P-Selectin for the Direct Platelet-Tumor Cell Contact During Metastasis Formation in Human Tumors. Front. Oncol. 2021, 11, 642761. [Google Scholar] [CrossRef]
- Conley, H.E.; Sheats, M.K. Targeting Neutrophil Β2-Integrins: A Review of Relevant Resources, Tools, and Methods. Biomolecules 2023, 13, 892. [Google Scholar] [CrossRef] [PubMed]
- Finsterbusch, M.; Schrottmaier, W.C.; Kral-Pointner, J.B.; Salzmann, M.; Assinger, A. Measuring and Interpreting Platelet-Leukocyte Aggregates. Platelets 2018, 29, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Rossaint, J.; Margraf, A.; Zarbock, A. Role of Platelets in Leukocyte Recruitment and Resolution of Inflammation. Front. Immunol. 2018, 9, 2712. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tang, C. Targeting Platelet in Atherosclerosis Plaque Formation: Current Knowledge and Future Perspectives. Int. J. Mol. Sci. 2020, 21, 9760. [Google Scholar] [CrossRef]
- Burnouf, T.; Chou, M.-L.; Lundy, D.J.; Chuang, E.-Y.; Tseng, C.-L.; Goubran, H. Expanding Applications of Allogeneic Platelets, Platelet Lysates, and Platelet Extracellular Vesicles in Cell Therapy, Regenerative Medicine, and Targeted Drug Delivery. J. Biomed. Sci. 2023, 30, 79. [Google Scholar] [CrossRef]
- Dai, Z.; Zhao, T.; Song, N.; Pan, K.; Yang, Y.; Zhu, X.; Chen, P.; Zhang, J.; Xia, C. Platelets and Platelet Extracellular Vesicles in Drug Delivery Therapy: A Review of the Current Status and Future Prospects. Front. Pharmacol. 2022, 13, 1026386. [Google Scholar] [CrossRef]
- Soriano Jerez, E.M.; Gibbins, J.M.; Hughes, C.E. Targeting Platelet Inhibition Receptors for Novel Therapies: PECAM-1 and G6b-B. Platelets 2021, 32, 761–769. [Google Scholar] [CrossRef]
- Chatterjee, M. Platelet Lipidome: Dismantling the “Trojan Horse” in the Bloodstream. J. Thromb. Haemost. 2020, 18, 543–557. [Google Scholar] [CrossRef]
- Marcinczyk, N.; Misztal, T.; Gromotowicz-Poplawska, A.; Zebrowska, A.; Rusak, T.; Radziwon, P.; Chabielska, E. Utility of Platelet Endothelial Cell Adhesion Molecule 1 in the Platelet Activity Assessment in Mouse and Human Blood. Int. J. Mol. Sci. 2021, 22, 9611. [Google Scholar] [CrossRef]
- Jansen, E.E.; Braun, A.; Jansen, P.; Hartmann, M. Platelet-Therapeutics to Improve Tissue Regeneration and Wound Healing—Physiological Background and Methods of Preparation. Biomedicines 2021, 9, 869. [Google Scholar] [CrossRef]
- Boilard, E.; Bellio, M. Platelet Extracellular Vesicles and the Secretory Interactome Join Forces in Health and Disease. Immunol. Rev. 2022, 312, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Komatsuya, K.; Kaneko, K.; Kasahara, K. Function of Platelet Glycosphingolipid Microdomains/Lipid Rafts. Int. J. Mol. Sci. 2020, 21, 5539. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, M.; Ehrenberg, A.; Toska, L.M.; Metz, L.M.; Klier, M.; Krueger, I.; Reusswig, F.; Elvers, M. Molecular Drivers of Platelet Activation: Unraveling Novel Targets for Anti-Thrombotic and Anti-Thrombo-Inflammatory Therapy. Int. J. Mol. Sci. 2020, 21, 7906. [Google Scholar] [CrossRef] [PubMed]
- Akkaya Fırat, A. Chemokines Effective on Platelet Functions. In Chemokines Updates; IntechOpen: London, UK, 2023. [Google Scholar]
- Giusti, I.; Rughetti, A.; D’Ascenzo, S.; Millimaggi, D.; Pavan, A.; Dell’Orso, L.; Dolo, V. Identification of an Optimal Concentration of Platelet Gel for Promoting Angiogenesis in Human Endothelial Cells. Transfusion 2009, 49, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Roweth, H.G.; Battinelli, E.M. Platelets and (Lymph) Angiogenesis. Cold Spring Harb. Perspect. Med. 2023, 13, a041174. [Google Scholar] [CrossRef]
- Ocansey, D.K.W.; Pei, B.; Xu, X.; Zhang, L.; Olovo, C.V.; Mao, F. Cellular and Molecular Mediators of Lymphangiogenesis in Inflammatory Bowel Disease. J. Transl. Med. 2021, 19, 254. [Google Scholar] [CrossRef]
- Hur, J.; Jang, J.H.; Oh, I.-Y.; Choi, J.-I.; Yun, J.-Y.; Kim, J.; Choi, Y.-E.; Ko, S.-B.; Kang, J.-A.; Kang, J.; et al. Human Podoplanin-Positive Monocytes and Platelets Enhance Lymphangiogenesis Through the Activation of the Podoplanin/CLEC-2 Axis. Mol. Ther. 2014, 22, 1518–1529. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, N.; Yu, J.; Xu, W.; Gao, J.; Lv, X.; Wen, Z. The Role of Podoplanin in the Immune System and Inflammation. J. Inflamm. Res. 2022, 15, 3561–3572. [Google Scholar] [CrossRef]
- Suzuki-Inoue, K.; Tsukiji, N.; Otake, S. Crosstalk between Hemostasis and Lymphangiogenesis. J. Thromb. Haemost. 2020, 18, 767–770. [Google Scholar] [CrossRef]
- Suzuki-Inoue, K.; Tsukiji, N. Platelet CLEC-2 and Lung Development. Res. Pract. Thromb. Haemost. 2020, 4, 481–490. [Google Scholar] [CrossRef]
- Bautch, V.L.; Caron, K.M. Blood and Lymphatic Vessel Formation. Cold Spring Harb. Perspect. Biol. 2015, 7, a008268. [Google Scholar] [CrossRef] [PubMed]
- Puricelli, C.; Boggio, E.; Gigliotti, C.L.; Stoppa, I.; Sutti, S.; Giordano, M.; Dianzani, U.; Rolla, R. Platelets, Protean Cells with All-Around Functions and Multifaceted Pharmacological Applications. Int. J. Mol. Sci. 2023, 24, 4565. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.; Li, Z.-T.; Xu, L.-H.; Su, T.-Y.; Han, Y.; Bao, M.; Liu, Z.; Fan, Y.-J.; Lou, Y.; Chen, Y.; et al. Platelet-Derived Extracellular Vesicles Increase Col8a1 Secretion and Vascular Stiffness in Intimal Injury. Front. Cell Dev. Biol. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Alberti, S.; Zhang, Q.; D’Agostino, I.; Bruno, A.; Tacconelli, S.; Contursi, A.; Guarnieri, S.; Dovizio, M.; Falcone, L.; Ballerini, P.; et al. The Antiplatelet Agent Revacept Prevents the Increase of Systemic Thromboxane A2 Biosynthesis and Neointima Hyperplasia. Sci. Rep. 2020, 10, 21420. [Google Scholar] [CrossRef] [PubMed]
- Subbotin, V.M. Analysis of Arterial Intimal Hyperplasia: Review and Hypothesis. Theor. Biol. Med. Model. 2007, 4, 41. [Google Scholar] [CrossRef] [PubMed]
- Liao, K.; Zhang, X.; Liu, J.; Teng, F.; He, Y.; Cheng, J.; Yang, Q.; Zhang, W.; Xie, Y.; Guo, D.; et al. The Role of Platelets in the Regulation of Tumor Growth and Metastasis: The Mechanisms and Targeted Therapy. MedComm 2023, 4, e350. [Google Scholar] [CrossRef] [PubMed]
- Bian, X.; Yin, S.; Yang, S.; Jiang, X.; Wang, J.; Zhang, M.; Zhang, L. Roles of Platelets in Tumor Invasion and Metastasis: A Review. Heliyon 2022, 8, e12072. [Google Scholar] [CrossRef]
- Schlesinger, M. Role of Platelets and Platelet Receptors in Cancer Metastasis. J. Hematol. Oncol. 2018, 11, 125. [Google Scholar] [CrossRef]
- Anvari, S.; Osei, E.; Maftoon, N. Interactions of Platelets with Circulating Tumor Cells Contribute to Cancer Metastasis. Sci. Rep. 2021, 11, 15477. [Google Scholar] [CrossRef]
- Mendoza-Almanza, G.; Burciaga-Hernández, L.; Maldonado, V.; Melendez-Zajgla, J.; Olmos, J. Role of Platelets and Breast Cancer Stem Cells in Metastasis. World J. Stem Cells 2020, 12, 1237–1254. [Google Scholar] [CrossRef]
- Eisinger, F.; Patzelt, J.; Langer, H.F. The Platelet Response to Tissue Injury. Front Med. 2018, 5, 317. [Google Scholar] [CrossRef] [PubMed]
- Hajdu, S.I. A Note from History: The Discovery of Blood Cells. Ann. Clin. Lab. Sci. 2003, 33, 237–238. [Google Scholar] [PubMed]
- Stassen, J.M.; Nyström, Å. A Historical Review of Hemostasis, Thrombosis, and Antithrombotic Therapy. Ann. Plast. Surg. 1997, 39, 317–329. [Google Scholar] [CrossRef] [PubMed]
- De Gaetano, G. Historical Overview of the Role of Platelets in Hemostasis and Thrombosis. Haematologica 2001, 86, 349–356. [Google Scholar] [PubMed]
- Sánchez-González, D.J.; Méndez-Bolaina, E.; Trejo-Bahena, N.I. Platelet-Rich Plasma Peptides: Key for Regeneration. Int. J. Pept. 2012, 2012, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zielins, E.R.; Atashroo, D.A.; Maan, Z.N.; Duscher, D.; Walmsley, G.G.; Hu, M.; Senarath-Yapa, K.; McArdle, A.; Tevlin, R.; Wearda, T.; et al. Wound Healing: An Update. Regen. Med. 2014, 9, 817–830. [Google Scholar] [CrossRef] [PubMed]
- Cronkite, E.P. Use of Thrombin and Fibrinogen in Skin Grafting. J. Am. Med. Assoc. 1944, 124, 976. [Google Scholar] [CrossRef]
- Raeissadat, S.A.; Babaee, M.; Rayegani, S.M.; Hashemi, Z.; Hamidieh, A.A.; Mojgani, P.; Vanda, H.F. An Overview of Platelet Products (PRP, PRGF, PRF, Etc.) in the Iranian Studies. Future Sci. OA 2017, 3, FSO231. [Google Scholar] [CrossRef]
- Fredriksson, L.; Li, H.; Eriksson, U. The PDGF Family: Four Gene Products Form Five Dimeric Isoforms. Cytokine Growth Factor. Rev. 2004, 15, 197–204. [Google Scholar] [CrossRef]
- Conde Montero, E.; Fernández Santos, M.E.; Suárez Fernández, R. Plasma Rico En Plaquetas: Aplicaciones En Dermatología. Actas Dermosifiliogr. 2015, 106, 104–111. [Google Scholar] [CrossRef]
- De Angelis, B.; D’Autilio, M.F.L.M.; Orlandi, F.; Pepe, G.; Garcovich, S.; Scioli, M.G.; Orlandi, A.; Cervelli, V.; Gentile, P. Wound Healing: In Vitro and In Vivo Evaluation of a Bio-Functionalized Scaffold Based on Hyaluronic Acid and Platelet-Rich Plasma in Chronic Ulcers. J. Clin. Med. 2019, 8, 1486. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Parayaruthottam, P.; Roshan, G.; Menon, V.; Fidha, M.; Fernandes, A. Platelets and Their Pathways in Dentistry: Systematic Review. J. Int. Soc. Prev. Community Dent. 2017, 7, 55. [Google Scholar] [CrossRef]
- Cervelli, V.; Bocchini, I.; Di Pasquali, C.; De Angelis, B.; Cervelli, G.; Curcio, C.B.; Orlandi, A.; Scioli, M.G.; Tati, E.; Delogu, P.; et al. P.R.L. Platelet Rich Lipotransfert: Our Experience and Current State of Art in the Combined Use of Fat and PRP. Biomed. Res. Int. 2013, 2013, 434191. [Google Scholar] [CrossRef] [PubMed]
- Ghanaati, S.; Herrera-Vizcaino, C.; Al-Maawi, S.; Lorenz, J.; Miron, R.J.; Nelson, K.; Schwarz, F.; Choukroun, J.; Sader, R. Fifteen Years of Platelet Rich Fibrin in Dentistry and Oromaxillofacial Surgery: How High Is the Level of Scientific Evidence? J. Oral Implantol. 2018, 44, 471–492. [Google Scholar] [CrossRef] [PubMed]
- Cieslik-Bielecka, A.; Choukroun, J.; Odin, G.; Dohan Ehrenfest, D.M. L-PRP/L-PRF in Esthetic Plastic Surgery, Regenerative Medicine of the Skin and Chronic Wounds. Curr. Pharm. Biotechnol. 2012, 13, 1266–1277. [Google Scholar] [CrossRef] [PubMed]
- Tözüm, T.F.; Demiralp, B. Platelet-Rich Plasma: A Promising Innovation in Dentistry. J. Can. Dent. Assoc. 2003, 69, 664. [Google Scholar]
- Anitua, E. Plasma Rich in Growth Factors: Preliminary Results of Use in the Preparation of Future Sites for Implants. Int. J. Oral Maxillofac. Implant. 1999, 14, 529–535. [Google Scholar]
- Ra Hara, G.; Basu, T. Platelet-Rich Plasma in Regenerative Medicine. Biomed. Res. Ther. 2014, 1, 5. [Google Scholar] [CrossRef]
- Miron, R.J.; Fujioka-Kobayashi, M.; Hernandez, M.; Kandalam, U.; Zhang, Y.; Ghanaati, S.; Choukroun, J. Injectable Platelet Rich Fibrin (i-PRF): Opportunities in Regenerative Dentistry? Clin. Oral. Investig. 2017, 21, 2619–2627. [Google Scholar] [CrossRef]
- Maisel-Campbell, A.L.; Ismail, A.; Reynolds, K.A.; Poon, E.; Serrano, L.; Grushchak, S.; Farid, C.; West, D.P.; Alam, M. A Systematic Review of the Safety and Effectiveness of Platelet-Rich Plasma (PRP) for Skin Aging. Arch. Dermatol. Res. 2020, 312, 301–315. [Google Scholar] [CrossRef]
- Gentile, P.; Garcovich, S.; Bielli, A.; Scioli, M.G.; Orlandi, A.; Cervelli, V. The Effect of Platelet-Rich Plasma in Hair Regrowth: A Randomized Placebo-Controlled Trial. Stem Cells Transl. Med. 2015, 4, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Gentile, P.; Calabrese, C.; De Angelis, B.; Dionisi, L.; Pizzicannella, J.; Kothari, A.; De Fazio, D.; Garcovich, S. Impact of the Different Preparation Methods to Obtain Autologous Non-Activated Platelet-Rich Plasma (A-PRP) and Activated Platelet-Rich Plasma (AA-PRP) in Plastic Surgery: Wound Healing and Hair Regrowth Evaluation. Int. J. Mol. Sci. 2020, 21, 431. [Google Scholar] [CrossRef] [PubMed]
- Cervelli, V.; Garcovich, S.; Bielli, A.; Cervelli, G.; Curcio, B.C.; Scioli, M.G.; Orlandi, A.; Gentile, P. The Effect of Autologous Activated Platelet Rich Plasma (AA-PRP) Injection on Pattern Hair Loss: Clinical and Histomorphometric Evaluation. Biomed. Res. Int. 2014, 2014, 760709. [Google Scholar] [CrossRef] [PubMed]
- Uebel, C.O.; da Silva, J.B.; Cantarelli, D.; Martins, P. The Role of Platelet Plasma Growth Factors in Male Pattern Baldness Surgery. Plast. Reconstr. Surg. 2006, 118, 1458–1466. [Google Scholar] [CrossRef] [PubMed]
- Al Taweel, A.I.; Al Refae, A.A.; Hamed, A.M.; Kamal, A.M. Comparative Study of the Efficacy of Platelet-rich Plasma Combined with Carboxytherapy vs Its Use with Fractional Carbon Dioxide Laser in Atrophic Acne Scars. J. Cosmet. Dermatol. 2019, 18, 150–155. [Google Scholar] [CrossRef] [PubMed]
- El-Domyati, M.; Abdel-Wahab, H.; Hossam, A. Microneedling Combined with Platelet-rich Plasma or Trichloroacetic Acid Peeling for Management of Acne Scarring: A Split-face Clinical and Histologic Comparison. J. Cosmet. Dermatol. 2018, 17, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Bos-Mikich, A.; de Oliveira, R.; Frantz, N. Platelet-Rich Plasma Therapy and Reproductive Medicine. J. Assist. Reprod. Genet. 2018, 35, 753–756. [Google Scholar] [CrossRef]
- Matz, E.L.; Pearlman, A.M.; Terlecki, R.P. Safety and Feasibility of Platelet Rich Fibrin Matrix Injections for Treatment of Common Urologic Conditions. Investig. Clin. Urol. 2018, 59, 61. [Google Scholar] [CrossRef]
- García-Conca, V.; Abad-Collado, M.; Hueso-Abancens, J.R.; Mengual-Verdú, E.; Piñero, D.P.; Aguirre-Balsalobre, F.; Molina, J.C. Efficacy and Safety of Treatment of Hyposecretory Dry Eye with Platelet-rich Plasma. Acta Ophthalmol. 2019, 97, E170–E178. [Google Scholar] [CrossRef]
- Arshdeep, M.; Kumaran, S. Platelet-Rich Plasma in Dermatology: Boon or a Bane? Indian J. Dermatol. Venereol. Leprol. 2014, 80, 5. [Google Scholar] [CrossRef]
- Dohan Ehrenfest, D.M.; Andia, I.; Zumstein, M.A.; Zhang, C.-Q.; Pinto, N.R.; Bielecki, T. Classification of Platelet Concentrates (Platelet-Rich Plasma-PRP, Platelet-Rich Fibrin-PRF) for Topical and Infiltrative Use in Orthopedic and Sports Medicine: Current Consensus, Clinical Implications and Perspectives. Muscles Ligaments Tendons J. 2014, 4, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Kon, E.; Di Matteo, B.; Delgado, D.; Cole, B.J.; Dorotei, A.; Dragoo, J.L.; Filardo, G.; Fortier, L.A.; Giuffrida, A.; Jo, C.H.; et al. Platelet-Rich Plasma for the Treatment of Knee Osteoarthritis: An Expert Opinion and Proposal for a Novel Classification and Coding System. Expert. Opin. Biol. Ther. 2020, 20, 1447–1460. [Google Scholar] [CrossRef] [PubMed]
- Everts, P.A.; Malanga, G.A.; Paul, R.V.; Rothenberg, J.B.; Stephens, N.; Mautner, K.R. Assessing Clinical Implications and Perspectives of the Pathophysiological Effects of Erythrocytes and Plasma Free Hemoglobin in Autologous Biologics for Use in Musculoskeletal Regenerative Medicine Therapies. A Review. Regen. Ther. 2019, 11, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Oneto, P.; Zubiry, P.R.; Schattner, M.; Etulain, J. Anticoagulants Interfere With the Angiogenic and Regenerative Responses Mediated by Platelets. Front. Bioeng. Biotechnol. 2020, 8, 223. [Google Scholar] [CrossRef] [PubMed]
- Finnerty, C.C.; Jeschke, M.G.; Branski, L.K.; Barret, J.P.; Dziewulski, P.; Herndon, D.N. Hypertrophic Scarring: The Greatest Unmet Challenge after Burn Injury. Lancet 2016, 388, 1427–1436. [Google Scholar] [CrossRef]
- Clementoni, M.T.; B-Roscher, M.; Munavalli, G.S. Photodynamic Photorejuvenation of the Face with a Combination of Microneedling, Red Light, and Broadband Pulsed Light. Lasers Surg. Med. 2010, 42, 150–159. [Google Scholar] [CrossRef]
- Elsayed, M. Evaluation of the Effect of Platelet-Rich Plasma on Post-Burn Scars. Open Access J. Surg. 2017, 5. [Google Scholar] [CrossRef]
- Wang, S.-L.; Liu, X.-L.; Kang, Z.-C.; Wang, Y.-S. Platelet-Rich Plasma Promotes Peripheral Nerve Regeneration after Sciatic Nerve Injury. Neural Regen. Res. 2023, 18, 375. [Google Scholar] [CrossRef]
- Hosseini, M.S.; Nouri, M.; Zarrabi, M.; Fatemi, M.J.; Shpichka, A.; Timashev, P.; Hassan, M.; Vosough, M. Platelet-Rich Plasma in Regenerative Medicine: Possible Applications in Management of Burns and Post-Burn Scars: A Review. Cell J. 2023, 25, 281–290. [Google Scholar] [CrossRef]
- Horgos, M.S.; Pop, O.L.; Sandor, M.; Borza, I.L.; Negrean, R.A.; Cote, A.; Neamtu, A.-A.; Grierosu, C.; Sachelarie, L.; Huniadi, A. Platelets Rich Plasma (PRP) Procedure in the Healing of Atonic Wounds. J. Clin. Med. 2023, 12, 3890. [Google Scholar] [CrossRef]
- Xu, P.; Wu, Y.; Zhou, L.; Yang, Z.; Zhang, X.; Hu, X.; Yang, J.; Wang, M.; Wang, B.; Luo, G.; et al. Platelet-Rich Plasma Accelerates Skin Wound Healing by Promoting Re-Epithelialization. Burns Trauma. 2020, 8, tkaa028. [Google Scholar] [CrossRef] [PubMed]
- Knightly, N.; Lee, C.; O’Brien, L.; Qayyum, T.; Hurley, C.; Kelly, J. Role for Platelet Rich Plasma as an Adjuvant Therapy in Wound Healing and Burns. Eur. J. Plast. Surg. 2023, 46, 465–474. [Google Scholar] [CrossRef]
- Imam, M.S.; Alotaibi, A.A.S.; Alotaibi, N.O.M.; Alosaimi, N.S.; Alotaibi, S.G.M.; Abdelrahim, M.E.A. Efficiency of Platelet-rich Plasma in the Management of Burn Wounds: A Meta-analysis. Int. Wound J. 2023. [Google Scholar] [CrossRef] [PubMed]
- Tammam, B.M.H.; Habotta, O.A.; El-khadragy, M.; Abdel Moneim, A.E.; Abdalla, M.S. Therapeutic Role of Mesenchymal Stem Cells and Platelet-Rich Plasma on Skin Burn Healing and Rejuvenation: A Focus on Scar Regulation, Oxido-Inflammatory Stress and Apoptotic Mechanisms. Heliyon 2023, 9, e19452. [Google Scholar] [CrossRef] [PubMed]
- Dogra, S.; Sarangal, R. What′s New in Cicatricial Alopecia? Indian. J. Dermatol. Venereol. Leprol. 2013, 79, 576. [Google Scholar] [CrossRef] [PubMed]
- França, K.; Rodrigues, T.S.; Ledon, J.; Savas, J.; Chacon, A. Comprehensive Overview and Treatment Update on Hair Loss. J. Cosmet. Dermatol. Sci. Appl. 2013, 3, 35881. [Google Scholar] [CrossRef]
- Sadgrove, N.; Batra, S.; Barreto, D.; Rapaport, J. An Updated Etiology of Hair Loss and the New Cosmeceutical Paradigm in Therapy: Clearing ‘the Big Eight Strikes’. Cosmetics 2023, 10, 106. [Google Scholar] [CrossRef]
- Sibbald, C. Alopecia Areata: An Updated Review for 2023. J. Cutan. Med. Surg. 2023, 27, 241–259. [Google Scholar] [CrossRef]
- Kaliyadan, F.; Nambiar, A.; Vijayaraghavan, S. Androgenetic Alopecia: An Update. Indian J. Dermatol. Venereol. Leprol. 2013, 79, 613. [Google Scholar] [CrossRef]
- Daunton, A.; Harries, M.; Sinclair, R.; Paus, R.; Tosti, A.; Messenger, A. Chronic Telogen Effluvium: Is It a Distinct Condition? A Systematic Review. Am. J. Clin. Dermatol. 2023, 24, 513–520. [Google Scholar] [CrossRef]
- Cindy Gasnier, R.B. Autologous Platelet Rich Plasma as a Treatment of Male Androgenetic Alopecia: Study of 14 Cases. J. Clin. Exp. Dermatol. Res. 2015, 6. [Google Scholar] [CrossRef]
- Gentile, P.; Cole, J.; Cole, M.; Garcovich, S.; Bielli, A.; Scioli, M.; Orlandi, A.; Insalaco, C.; Cervelli, V. Evaluation of Not-Activated and Activated PRP in Hair Loss Treatment: Role of Growth Factor and Cytokine Concentrations Obtained by Different Collection Systems. Int. J. Mol. Sci. 2017, 18, 408. [Google Scholar] [CrossRef] [PubMed]
- Singhal, P.; Agarwal, S.; Dhot, P.; Sayal, S. Efficacy of Platelet-Rich Plasma in Treatment of Androgenic Alopecia. Asian J. Transfus. Sci. 2015, 9, 159. [Google Scholar] [CrossRef] [PubMed]
- Khatu, S.; More, Y.; Gokhale, N.; Chavhan, D.; Bendsure, N. Platelet-Rich Plasma in Androgenic Alopecia: Myth or an Effective Tool. J. Cutan. Aesthet. Surg. 2014, 7, 107. [Google Scholar] [CrossRef] [PubMed]
- Greco, J.; Brandt, R. The Effects of Autologus Platelet Rich Plasma and Various Growth Factors on Non-Transplanted Miniaturized Hair. Int. Soc. Hair Restor. Surg. 2009, 19, 49–50. [Google Scholar] [CrossRef]
- Stevens, J.; Khetarpal, S. Platelet-rich plasma for androgenetic alopecia: A review of the literature and proposed treatment protocol. Int. J. Women Dermatol. 2019, 5, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Garg, S. Outcome of Intra-Operative Injected Platelet-Rich Plasma Therapy during Follicular Unit Extraction Hair Transplant: A Prospective Randomised Study in Forty Patients. J. Cutan. Aesthet. Surg. 2016, 9, 157. [Google Scholar] [CrossRef]
- Natarelli, N.; Gahoonia, N.; Sivamani, R.K. Integrative and Mechanistic Approach to the Hair Growth Cycle and Hair Loss. J. Clin. Med. 2023, 12, 893. [Google Scholar] [CrossRef]
- Shimizu, Y.; Ntege, E.H.; Sunami, H.; Inoue, Y. Regenerative Medicine Strategies for Hair Growth and Regeneration: A Narrative Review of Literature. Regen. Ther. 2022, 21, 527–539. [Google Scholar] [CrossRef]
- Kang, J.-S.; Zheng, Z.; Choi, M.J.; Lee, S.-H.; Kim, D.-Y.; Cho, S.B. The Effect of CD34+ Cell-containing Autologous Platelet-rich Plasma Injection on Pattern Hair Loss: A Preliminary Study. J. Eur. Acad. Dermatol. Venereol. 2014, 28, 72–79. [Google Scholar] [CrossRef]
- Li, Z.J.; Choi, H.-I.; Choi, D.-K.; Sohn, K.-C.; Im, M.; Seo, Y.-J.; Lee, Y.-H.; Lee, J.-H.; Lee, Y. Autologous Platelet-Rich Plasma: A Potential Therapeutic Tool for Promoting Hair Growth. Dermatol. Surg. 2012, 38, 1040–1046. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Won, C.H.; Chung, W.-K.; Park, B.-S. Up-to-Date Clinical Trials of Hair Regeneration Using Conditioned Media of Adipose-Derived Stem Cells in Male and Female Pattern Hair Loss. Curr. Stem Cell Res. Ther. 2017, 12, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Leo, M.S.; Kumar, A.S.; Kirit, R.; Konathan, R.; Sivamani, R.K. Systematic Review of the Use of Platelet-rich Plasma in Aesthetic Dermatology. J. Cosmet. Dermatol. 2015, 14, 315–323. [Google Scholar] [CrossRef]
- Trink, A.; Sorbellini, E.; Bezzola, P.; Rodella, L.; Rezzani, R.; Ramot, Y.; Rinaldi, F. A Randomized, Double-Blind, Placebo- and Active-Controlled, Half-Head Study to Evaluate the Effects of Platelet-Rich Plasma on Alopecia Areata. Br. J. Dermatol. 2013, 169, 690–694. [Google Scholar] [CrossRef]
- Alkhalifah, A.; Alsantali, A.; Wang, E.; McElwee, K.J.; Shapiro, J. Alopecia Areata Update. J. Am. Acad. Dermatol. 2010, 62, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Teng, H.; Chen, H. Platelet-rich Plasma in the Treatment of Alopecia Areata after COVID-19 Vaccination. Clin. Case Rep. 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Todorova, L.N.; Abadjieva, T.I. Platelet-Rich Plasma in Alopecia Areata: A Case Report With a Mini Review of Literature. Cureus 2023, 15, e38751. [Google Scholar] [CrossRef] [PubMed]
- Pensato, R.; Al-Amer, R.; La Padula, S. Platelet-Rich Plasma for Treating Androgenic Alopecia: A Systematic Review. Aesthetic Plast. Surg. 2023. [Google Scholar] [CrossRef]
- Singh, S. Role of Platelet-Rich Plasma in Chronic Alopecia Areata: Our Centre Experience. Indian J. Plast. Surg. 2015, 48, 057–059. [Google Scholar] [CrossRef]
- Donovan, J. Successful Treatment of Corticosteroid-Resistant Ophiasis-Type Alopecia Areata (AA) with Platelet-Rich Plasma (PRP). JAAD Case Rep. 2015, 1, 305–307. [Google Scholar] [CrossRef][Green Version]
- El Taieb, M.A.; Ibrahim, H.; Nada, E.A.; Seif Al-Din, M. Platelets Rich Plasma versus Minoxidil 5% in Treatment of Alopecia Areata: A Trichoscopic Evaluation. Dermatol. Ther. 2017, 30, e12437. [Google Scholar] [CrossRef] [PubMed]
- Meyers, A.; Jin, A.; Kwiecien, G.J.; Gatherwright, J.; Khetarpal, S.; Zins, J.E. Platelet-Rich Plasma for Treatment of Hair Loss Improves Patient-Reported Quality of Life. Aesthetic Plast. Surg. 2023, 47, 1528–1534. [Google Scholar] [CrossRef] [PubMed]
- El-Dawla, R.E.; Abdelhaleem, M.; Abdelhamed, A. Evaluation of the Safety and Efficacy of Platelet-Rich Plasma in the Treatment of Female Patients with Chronic Telogen Effluvium: A Randomised, Controlled, Double-Blind, Pilot Clinical Trial. Indian J. Dermatol. Venereol. Leprol. 2022, 89, 195. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.-M.; Li, Z.-X.; Zhang, D.-Y.; Yang, Y.-C.; Fu, S.; Zhang, Z.-Q.; Yang, R.-H.; Xiong, K. A Systematic Summary of Survival and Death Signalling during the Life of Hair Follicle Stem Cells. Stem. Cell Res. Ther. 2021, 12, 453. [Google Scholar] [CrossRef] [PubMed]
- Paththinige, N.D.; Akarawita, J.K.W.; Jeganathan, G. The Clinical Efficacy and Safety of Autologous Activated Platelet-Rich Plasma Injection in Androgenetic Alopecia. Skin. Appendage Disord. 2020, 6, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Gentile, P.; Garcovich, S. Systematic Review of Platelet-Rich Plasma Use in Androgenetic Alopecia Compared with Minoxidil®, Finasteride®, and Adult Stem Cell-Based Therapy. Int. J. Mol. Sci. 2020, 21, 2702. [Google Scholar] [CrossRef] [PubMed]
- Alves, R.; Grimalt, R. A Review of Platelet-Rich Plasma: History, Biology, Mechanism of Action, and Classification. Skin. Appendage Disord. 2018, 4, 18–24. [Google Scholar] [CrossRef]
- El-Fakahany, H.; Raouf, H.A.; Medhat, W. Using Automated Microneedling with Platelet Rich Plasma for Treating Cicatricial Alopecia, Recalcitrant Alopecia Areata and Traction Alopecia, Case Report. Proc. J. Am. Acad. Dermatol. 2016, AB140. [Google Scholar]
- Verma, R.; Kumar, S.; Garg, P.; Verma, Y.K. Platelet-Rich Plasma: A Comparative and Economical Therapy for Wound Healing and Tissue Regeneration. Cell Tissue Bank. 2023, 24, 285–306. [Google Scholar] [CrossRef]
- Saxena, K.; Saxena, D.; Savant, S. Successful Hair Transplant Outcome in Cicatricial Lichen Planus of the Scalp by Combining Scalp and Beard Hair along with Platelet Rich Plasma. J. Cutan. Aesthet. Surg. 2016, 9, 51. [Google Scholar] [CrossRef]
- Pierce, G.F.; Mustoe, T.A.; Lingelbach, J.; Masakowski, V.R.; Griffin, G.L.; Senior, R.M.; Deuel, T.F. Platelet-Derived Growth Factor and Transforming Growth Factor-Beta Enhance Tissue Repair Activities by Unique Mechanisms. J. Cell Biol. 1989, 109, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Reddi, A.H. Role of Morphogenetic Proteins in Skeletal Tissue Engineering and Regeneration. Nat. Biotechnol. 1998, 16, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; Davis-Smyth, T. The Biology of Vascular Endothelial Growth Factor. Endocr. Rev. 1997, 18, 4–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Nan, W.; Wang, S.; Zhang, T.; Si, H.; Yang, F.; Li, G. Epidermal Growth Factor Promotes Proliferation and Migration of Follicular Outer Root Sheath Cells via Wnt/β-Catenin Signaling. Cell. Physiol. Biochem. 2016, 39, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Burgess, W.H.; Maciag, T. The Heparin-Binding (Fibroblast) Growth Factor Family of Proteins. Annu. Rev. Biochem. 1989, 58, 575–602. [Google Scholar] [CrossRef] [PubMed]
- Redaelli, A.; Romano, D.; Marcianó, A. Face and Neck Revitalization with Platelet-Rich Plasma (PRP): Clinical Outcome in a Series of 23 Consecutively Treated Patients. J. Drugs Dermatol. 2010, 9, 466–472. [Google Scholar]
- Elnehrawy, N.Y.; Ibrahim, Z.A.; Eltoukhy, A.M.; Nagy, H.M. Assessment of the Efficacy and Safety of Single Platelet-rich Plasma Injection on Different Types and Grades of Facial Wrinkles. J. Cosmet. Dermatol. 2017, 16, 103–111. [Google Scholar] [CrossRef]
- Yuksel, E.P.; Sahin, G.; Aydin, F.; Senturk, N.; Turanli, A.Y. Evaluation of Effects of Platelet-Rich Plasma on Human Facial Skin. J. Cosmet. Laser Ther. 2014, 16, 206–208. [Google Scholar] [CrossRef]
- Hofny, E.R.M.; Abdel-Motaleb, A.A.; Ghazally, A.; Ahmed, A.M.; Hussein, M.R.A. Platelet-Rich Plasma Is a Useful Therapeutic Option in Melasma. J. Dermatol. Treat. 2019, 30, 396–401. [Google Scholar] [CrossRef]
- Cameli, N.; Mariano, M.; Cordone, I.; Abril, E.; Masi, S.; Foddai, M.L. Autologous Pure Platelet-Rich Plasma Dermal Injections for Facial Skin Rejuvenation: Clinical, Instrumental, and Flow Cytometry Assessment. Dermatol. Surg. 2017, 43, 826–835. [Google Scholar] [CrossRef]
- BUZALAF, M.A.R.; LEVY, F.M. Autologous Platelet Concentrates for Facial Rejuvenation. J. Appl. Oral Sci. 2022, 30, e20220020. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.K.; Shin, M.K.; Lee, J.H.; Kim, N.I. Effects of Platelet-Rich Plasma on Wrinkles and Skin Tone in Asian Lower Eyelid Skin: Preliminary Results from a Prospective, Randomised, Split-Face Trial. Eur. J. Dermatol. 2014, 24, 100–101. [Google Scholar] [CrossRef] [PubMed]
- Mehryan, P.; Zartab, H.; Rajabi, A.; Pazhoohi, N.; Firooz, A. Assessment of Efficacy of Platelet-rich Plasma (PRP) on Infraorbital Dark Circles and Crow’s Feet Wrinkles. J. Cosmet. Dermatol. 2014, 13, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Totey, S.; Dhurat, R.; Kadam, P.; Sevilla, G.; Shetty, G. Safety and Efficacy of Growth Factor Concentrate in the Treatment of Nasolabial Fold Correction: Split Face Pilot Study. Indian. J. Dermatol. 2015, 60, 520. [Google Scholar] [CrossRef] [PubMed]
- Sclafani, A.P. Platelet-Rich Fibrin Matrix for Improvement of Deep Nasolabial Folds. J. Cosmet. Dermatol. 2010, 9, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Everts, P.A.; Pinto, P.C.; Girão, L. Autologous Pure Platelet-rich Plasma Injections for Facial Skin Rejuvenation: Biometric Instrumental Evaluations and Patient-reported Outcomes to Support Antiaging Effects. J. Cosmet. Dermatol. 2019, 18, 985–995. [Google Scholar] [CrossRef] [PubMed]
- Gawdat, H.I.; Hegazy, R.A.; Fawzy, M.M.; Fathy, M. Autologous Platelet Rich Plasma: Topical Versus Intradermal After Fractional Ablative Carbon Dioxide Laser Treatment of Atrophic Acne Scars. Dermatol. Surg. 2014, 40, 152–161. [Google Scholar] [CrossRef]
- Chawla, S. Split Face Comparative Study of Microneedling with PRP versus Microneedling with Vitamin C in Treating Atrophic Post Acne Scars. J. Cutan. Aesthet. Surg. 2014, 7, 209. [Google Scholar] [CrossRef]
- Alster, T.S.; Graham, P.M. Microneedling: A Review and Practical Guide. Dermatol. Surg. 2018, 44, 397–404. [Google Scholar] [CrossRef]
- Asif, M.; Kanodia, S.; Singh, K. Combined Autologous Platelet-rich Plasma with Microneedling Verses Microneedling with Distilled Water in the Treatment of Atrophic Acne Scars: A Concurrent Split-face Study. J. Cosmet. Dermatol. 2016, 15, 434–443. [Google Scholar] [CrossRef]
- Nofal, E.; Helmy, A.; Nofal, A.; Alakad, R.; Nasr, M. Platelet-Rich Plasma versus CROSS Technique with 100% Trichloroacetic Acid versus Combined Skin Needling and Platelet Rich Plasma in the Treatment of Atrophic Acne Scars: A Comparative Study. Dermatol. Surg. 2014, 40, 864–873. [Google Scholar] [PubMed]
- Ibrahim, Z.A.; El-Ashmawy, A.A.; Shora, O.A. Therapeutic Effect of Microneedling and Autologous Platelet-rich Plasma in the Treatment of Atrophic Scars: A Randomized Study. J. Cosmet. Dermatol. 2017, 16, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.K.; Ibrahim, S.M.; Salem, A.M. Skin Microneedling plus Platelet-Rich Plasma versus Skin Microneedling Alone in the Treatment of Atrophic Post Acne Scars: A Split Face Comparative Study. J. Dermatol. Treat. 2018, 29, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Faghihi, G.; Keyvan, S.; Asilian, A.; Nouraei, S.; Behfar, S.; Nilforoushzadeh, M. Efficacy of Autologous Platelet-Rich Plasma Combined with Fractional Ablative Carbon Dioxide Resurfacing Laser in Treatment of Facial Atrophic Acne Scars: A Split-Face Randomized Clinical Trial. Indian J. Dermatol. Venereol. Leprol. 2016, 82, 162. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Kim, B.J.; Kim, M.N.; Mun, S.K. The Efficacy of Autologous Platelet Rich Plasma Combined with Ablative Carbon Dioxide Fractional Resurfacing for Acne Scars: A Simultaneous Split-Face Trial. Dermatol. Surg. 2011, 37, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Kar, B.; Raj, C. Fractional CO 2 Laser vs Fractional CO 2 with Topical Platelet-Rich Plasma in the Treatment of Acne Scars: A Split-Face Comparison Trial. J. Cutan. Aesthet. Surg. 2017, 10, 136. [Google Scholar] [CrossRef]
- Min, S.; Yoon, J.Y.; Park, S.Y.; Moon, J.; Kwon, H.H.; Suh, D.H. Combination of Platelet Rich Plasma in Fractional Carbon Dioxide Laser Treatment Increased Clinical Efficacy of for Acne Scar by Enhancement of Collagen Production and Modulation of Laser-induced Inflammation. Lasers Surg. Med. 2018, 50, 302–310. [Google Scholar] [CrossRef]
- Zhu, J.-T.; Xuan, M.; Zhang, Y.-N.; Liu, H.-W.; Cai, J.-H.; Wu, Y.-H.; Xiang, X.-F.; Shan, G.-Q.; Cheng, B. The Efficacy of Autologous Platelet-Rich Plasma Combined with Erbium Fractional Laser Therapy for Facial Acne Scars or Acne. Mol. Med. Rep. 2013, 8, 233–237. [Google Scholar] [CrossRef]
- Zhao, L.; Hu, M.; Xiao, Q.; Zhou, R.; Li, Y.; Xiong, L.; Li, L. Efficacy and Safety of Platelet-Rich Plasma in Melasma: A Systematic Review and Meta-Analysis. Dermatol. Ther. 2021, 11, 1587–1597. [Google Scholar] [CrossRef]
- Ogbechie-Godec, O.A.; Elbuluk, N. Melasma: An Up-to-Date Comprehensive Review. Dermatol. Ther. 2017, 7, 305–318. [Google Scholar] [CrossRef]
- Maddaleno, A.S.; Camargo, J.; Mitjans, M.; Vinardell, M.P. Melanogenesis and Melasma Treatment. Cosmetics 2021, 8, 82. [Google Scholar] [CrossRef]
- Liu, W.; Chen, Q.; Xia, Y. New Mechanistic Insights of Melasma. Clin. Cosmet. Investig. Dermatol. 2023, 16, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Morgado-Carrasco, D.; Piquero-Casals, J.; Granger, C.; Trullàs, C.; Passeron, T. Melasma: The Need for Tailored Photoprotection to Improve Clinical Outcomes. Photodermatol. Photoimmunol. Photomed. 2022, 38, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Qari, S.; Bader, M.; Farran, E.; Borrah, R.; Khamis, S.; Alharbi, Z. Combined Synergetic Effect of Lipoconcentrate Fat Grafting, Nanofat Transfer, Platelet-Rich Plasma, Microneedling, and CO2 Fractional Laser for Plastic Regenerative and Esthetic Surgery and Cosmetic Care. Cureus 2023, 15, e44035. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Zhang, C.; Tuan, R.S. Biology of Platelet-Rich Plasma and Its Clinical Application in Cartilage Repair. Arthritis Res. Ther. 2014, 16, 204. [Google Scholar] [CrossRef]
- Chamata, E.S.; Bartlett, E.L.; Weir, D.; Rohrich, R.J. Platelet-Rich Plasma: Evolving Role in Plastic Surgery. Plast. Reconstr. Surg. 2021, 147, 219–230. [Google Scholar] [CrossRef]
- Dohan Ehrenfest, D.M.; Rasmusson, L.; Albrektsson, T. Classification of Platelet Concentrates: From Pure Platelet-Rich Plasma (P-PRP) to Leucocyte- and Platelet-Rich Fibrin (L-PRF). Trends Biotechnol. 2009, 27, 158–167. [Google Scholar] [CrossRef]
- Magalon, J.; Chateau, A.L.; Bertrand, B.; Louis, M.L.; Silvestre, A.; Giraudo, L.; Veran, J.; Sabatier, F. DEPA Classification: A Proposal for Standardising PRP Use and a Retrospective Application of Available Devices. BMJ Open Sport. Exerc. Med. 2016, 2, e000060. [Google Scholar] [CrossRef]
- Kang, Y.; Zhu, Z.; Zheng, Y.; Wan, W.; Manole, C.G.; Zhang, Q. Skin Telocytes versus Fibroblasts: Two Distinct Dermal Cell Populations. J. Cell Mol. Med. 2015, 19, 2530–2539. [Google Scholar] [CrossRef]
- Manole, C.G.; Simionescu, O. The Cutaneous Telocytes. Adv. Exp. Med. Biol. 2016, 913, 303–323. [Google Scholar] [CrossRef]
- Manole, C.G.; Gherghiceanu, M.; Ceafalan, L.C.; Hinescu, M.E. Dermal Telocytes: A Different Viewpoint of Skin Repairing and Regeneration. Cells 2022, 11, 3903. [Google Scholar] [CrossRef] [PubMed]
- Manole, C.G.; Cismaşiu, V.; Gherghiceanu, M.; Popescu, L.M. Experimental Acute Myocardial Infarction: Telocytes Involvement in Neo-Angiogenesis. J. Cell Mol. Med. 2011, 15, 2284–2296. [Google Scholar] [CrossRef] [PubMed]
- Cismasiu, V.B.; Radu, E.; Popescu, L.M. MiR-193 Expression Differentiates Telocytes from Other Stromal Cells. J. Cell Mol. Med. 2011, 15, 1071–1074. [Google Scholar] [CrossRef] [PubMed]
- Nicolescu, M.I.; Manole, C.G. Telocytes and Stem Cells in Regenerative Medicine. FASEB J. 2013, 27, 752.4. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).