
Molecular Investigation of Mitochondrial RNA19 Role in the Pathogenesis of MELAS Disease




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Culture and Heteroplasmy Determination
2.2. Construct Preparation and Cterm Expression
2.3. Immunoprecipitation of Cterm-FLAG
2.4. Isokinetic Sucrose Gradient Analysis
2.5. Western Blotting
2.6. RT-qPCR Analysis
2.7. Pulse-Labelling of Mitochondrial Translation Products
3. Results
3.1. Mitoribosomal Subunits Were Normally Assembled in Translation-Defective MELAS Cybrids
3.2. RNA19 Accumulated in the Gradient Denser Fractions from Mutant Cells
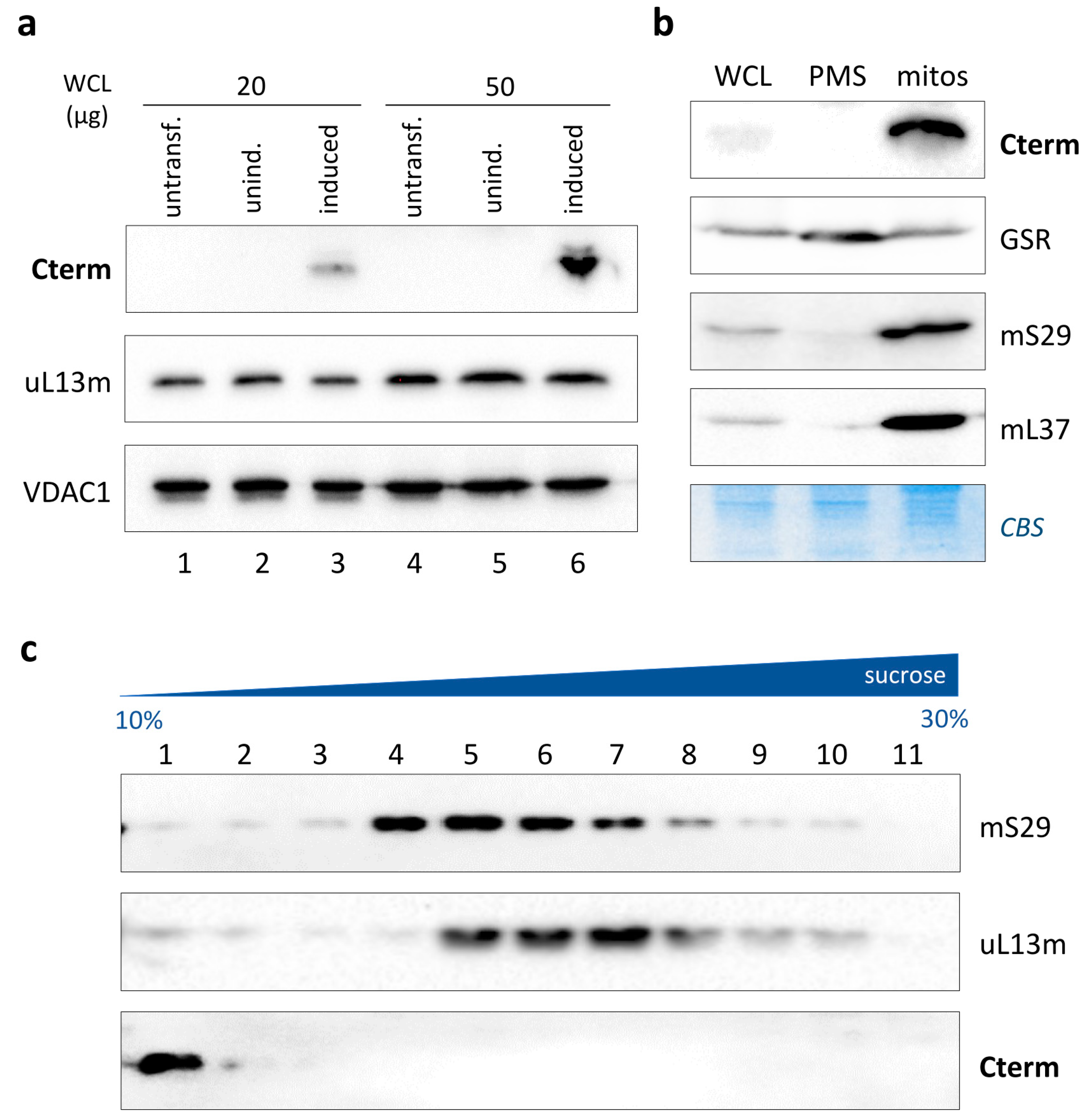
3.3. Cterm Did Not Associate with Mitoribosomal Subunits
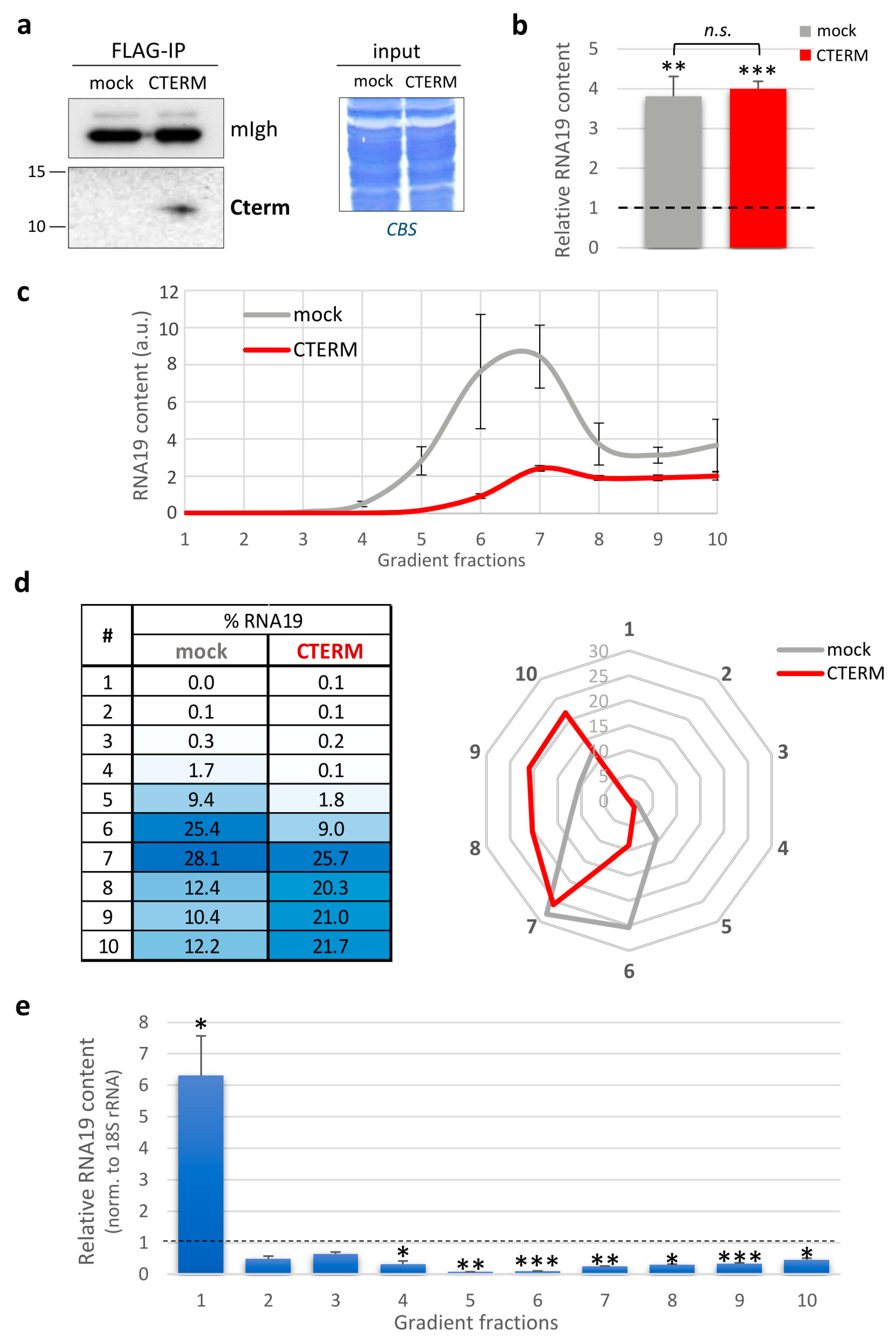
3.4. Cterm Expression Affected RNA19 Sedimentation Profile
4. Discussion
4.1. RNA19 Sedimentation Profile in Pathological Conditions
4.2. The Rescuing Activity of the Isolated LARS2 C-Terminal Domain
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anderson, S.; Bankier, A.T.; Barrell, B.G.; de Bruijn, M.H.L.; Coulson, A.R.; Drouin, J.; Eperon, I.C.; Nierlich, D.P.; Roe, B.A.; Sanger, F.; et al. Sequence and Organization of the Human Mitochondrial Genome. Nature 1981, 290, 457–465. [Google Scholar] [CrossRef]
- Antonicka, H.; Shoubridge, E.A. Mitochondrial RNA Granules Are Centers for Posttranscriptional RNA Processing and Ribosome Biogenesis. Cell Rep. 2015, 10, 920–932. [Google Scholar] [CrossRef] [PubMed]
- Jedynak-Slyvka, M.; Jabczynska, A.; Szczesny, R.J. Human Mitochondrial RNA Processing and Modifications: Overview. Int. J. Mol. Sci. 2021, 22, 7999. [Google Scholar] [CrossRef] [PubMed]
- Ojala, D.; Montoya, J.; Attardi, G. TRNA Punctuation Model of RNA Processing in Human Mitochondria. Nature 1981, 290, 470–474. [Google Scholar] [CrossRef]
- Temperley, R.J.; Wydro, M.; Lightowlers, R.N.; Chrzanowska-Lightowlers, Z.M. Human Mitochondrial MRNAs—Like Members of All Families, Similar but Different. Biochim. Biophys. Acta 2010, 1797, 1081–1085. [Google Scholar] [CrossRef]
- Bindoff, L.A.; Howell, N.; Poulton, J.; McCullough, D.A.; Morten, K.J.; Lightowlers, R.N.; Turnbull, D.M.; Weber, K. Abnormal RNA Processing Associated with a Novel TRNA Mutation in Mitochondrial DNA. A Potential Disease Mechanism. J. Biol. Chem. 1993, 268, 19559–19564. [Google Scholar] [CrossRef] [PubMed]
- King, M.P.; Koga, Y.; Davidson, M.; Schon, E.A. Defects in Mitochondrial Protein Synthesis and Respiratory Chain Activity Segregate with the TRNA(Leu(UUR)) Mutation Associated with Mitochondrial Myopathy, Encephalopathy, Lactic Acidosis, and Strokelike Episodes. Mol. Cell Biol. 1992, 12, 480–490. [Google Scholar] [CrossRef] [PubMed]
- Schon, E.A.; Koga, Y.; Davidson, M.; Moraes, C.T.; King, M.P. The Mitochondrial TRNA(Leu)(UUR)) Mutation in MELAS: A Model for Pathogenesis. Biochim. Biophys. Acta 1992, 1101, 206–209. [Google Scholar]
- Koga, Y.; Davidson, M.; Schon, E.A.; King, M.P. Fine Mapping of Mitochondrial RNAs Derived from the MtDNA Region Containing a Point Mutation Associated with MELAS. Nucleic Acids Res. 1993, 21, 657–662. [Google Scholar] [CrossRef]
- Rossmanith, W.; Karwan, R.M. Impairment of TRNA Processing by Point Mutations in Mitochondrial TRNALeu(UUR) Associated with Mitochondrial Diseases. FEBS Lett. 1998, 433, 269–274. [Google Scholar] [CrossRef]
- Levinger, L.; Oestreich, I.; Florentz, C.; Mörl, M. A Pathogenesis-Associated Mutation in Human Mitochondrial TRNALeu(UUR) Leads to Reduced 3′-End Processing and CCA Addition. J. Mol. Biol. 2004, 337, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Chomyn, A.; Enriquez, J.A.; Micol, V.; Fernandez-Silva, P.; Attardi, G. The Mitochondrial Myopathy, Encephalopathy, Lactic Acidosis, and Stroke-like Episode Syndrome-Associated Human Mitochondrial TRNALeu(UUR) Mutation Causes Aminoacylation Deficiency and Concomitant Reduced Association of MRNA with Ribosomes. J. Biol. Chem. 2000, 275, 19198–19209. [Google Scholar] [CrossRef] [PubMed]
- Koga, A.; Koga, Y.; Akita, Y.; Fukiyama, R.; Ueki, I.; Yatsuga, S.; Matsuishi, T. Increased Mitochondrial Processing Intermediates Associated with Three TRNALeu(UUR) Gene Mutations. Neuromuscul. Disord. 2003, 13, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Maniura-Weber, K.; Helm, M.; Engemann, K.; Eckertz, S.; Möllers, M.; Schauen, M.; Hayrapetyan, A.; von Kleist-Retzow, J.-C.; Lightowlers, R.N.; Bindoff, L.A.; et al. Molecular Dysfunction Associated with the Human Mitochondrial 3302A>G Mutation in the MTTL1 (Mt-TRNALeu(UUR)) Gene. Nucleic Acids Res. 2006, 34, 6404–6415. [Google Scholar] [CrossRef] [PubMed]
- Siira, S.J.; Spåhr, H.; Shearwood, A.-M.J.; Ruzzenente, B.; Larsson, N.-G.; Rackham, O.; Filipovska, A. LRPPRC-Mediated Folding of the Mitochondrial Transcriptome. Nat. Commun. 2017, 8, 1532. [Google Scholar] [CrossRef]
- Tani, H.; Ishikawa, K.; Tamashiro, H.; Ogasawara, E.; Yasukawa, T.; Matsuda, S.; Shimizu, A.; Kang, D.; Hayashi, J.-I.; Wei, F.-Y.; et al. Aberrant RNA Processing Contributes to the Pathogenesis of Mitochondrial Diseases in Trans-Mitochondrial Mouse Model Carrying Mitochondrial TRNALeu(UUR) with a Pathogenic A2748G Mutation. Nucleic Acids Res. 2022, 50, 9382–9396. [Google Scholar] [CrossRef]
- Capriglia, F.; Rizzo, F.; Petrosillo, G.; Morea, V.; d’Amati, G.; Cantatore, P.; Roberti, M.; Loguercio Polosa, P.; Bruni, F. Exploring the Ability of LARS2 Carboxy-Terminal Domain in Rescuing the MELAS Phenotype. Life 2021, 11, 674. [Google Scholar] [CrossRef]
- Perli, E.; Giordano, C.; Pisano, A.; Montanari, A.; Campese, A.F.; Reyes, A.; Ghezzi, D.; Nasca, A.; Tuppen, H.A.; Orlandi, M.; et al. The Isolated Carboxy-Terminal Domain of Human Mitochondrial Leucyl-TRNA Synthetase Rescues the Pathological Phenotype of Mitochondrial TRNA Mutations in Human Cells. EMBO Mol. Med. 2014, 6, 169–182. [Google Scholar] [CrossRef]
- Hornig-Do, H.T.; Montanari, A.; Rozanska, A.; Tuppen, H.A.; Almalki, A.A.; Abg-Kamaludin, D.P.; Frontali, L.; Francisci, S.; Lightowlers, R.N.; Chrzanowska-Lightowlers, Z.M. Human Mitochondrial Leucyl TRNA Synthetase Can Suppress Non Cognate Pathogenic Mt-TRNA Mutations. EMBO Mol. Med. 2014, 6, 183–193. [Google Scholar] [CrossRef]
- Bruni, F.; Gramegna, P.; Oliveira, J.M.A.; Lightowlers, R.N.; Chrzanowska-Lightowlers, Z.M.A. REXO2 Is an Oligoribonuclease Active in Human Mitochondria. PLoS ONE 2013, 8, e64670. [Google Scholar] [CrossRef]
- Cámara, Y.; Asin-Cayuela, J.; Park, C.B.; Metodiev, M.D.; Shi, Y.; Ruzzenente, B.; Kukat, C.; Habermann, B.; Wibom, R.; Hultenby, K.; et al. MTERF4 Regulates Translation by Targeting the Methyltransferase NSUN4 to the Mammalian Mitochondrial Ribosome. Cell Metab. 2011, 13, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Lopez Sanchez, M.I.G.; Krüger, A.; Shiriaev, D.I.; Liu, Y.; Rorbach, J. Human Mitoribosome Biogenesis and Its Emerging Links to Disease. Int. J. Mol. Sci. 2021, 22, 3827. [Google Scholar] [CrossRef] [PubMed]
- Perks, K.L.; Rossetti, G.; Kuznetsova, I.; Hughes, L.A.; Ermer, J.A.; Ferreira, N.; Busch, J.D.; Rudler, D.L.; Spahr, H.; Schöndorf, T.; et al. PTCD1 Is Required for 16S RRNA Maturation Complex Stability and Mitochondrial Ribosome Assembly. Cell Rep. 2018, 23, 127–142. [Google Scholar] [CrossRef] [PubMed]
- Metodiev, M.D.; Thompson, K.; Alston, C.L.; Morris, A.A.M.; He, L.; Assouline, Z.; Rio, M.; Bahi-Buisson, N.; Pyle, A.; Griffin, H.; et al. Recessive Mutations in TRMT10C Cause Defects in Mitochondrial RNA Processing and Multiple Respiratory Chain Deficiencies. Am. J. Human. Genet. 2016, 98, 993–1000. [Google Scholar] [CrossRef]
- Rackham, O.; Busch, J.D.; Matic, S.; Siira, S.J.; Kuznetsova, I.; Atanassov, I.; Ermer, J.A.; Shearwood, A.-M.J.; Richman, T.R.; Stewart, J.B.; et al. Hierarchical RNA Processing Is Required for Mitochondrial Ribosome Assembly. Cell Rep. 2016, 16, 1874–1890. [Google Scholar] [CrossRef]
- Siira, S.J.; Rossetti, G.; Richman, T.R.; Perks, K.; Ermer, J.A.; Kuznetsova, I.; Hughes, L.; Shearwood, A.-M.J.; Viola, H.M.; Hool, L.C.; et al. Concerted Regulation of Mitochondrial and Nuclear Non-Coding RNAs by a Dual-Targeted RNase Z. EMBO Rep. 2018, 19, e46198. [Google Scholar] [CrossRef]
- Zuccotti, P.; Modelska, A. Studying the Translatome with Polysome Profiling. Methods Mol. Biol. 2016, 1358, 59–69. [Google Scholar] [CrossRef]
- Jourdain, A.A.; Koppen, M.; Wydro, M.; Rodley, C.D.; Lightowlers, R.N.; Chrzanowska-Lightowlers, Z.M.; Martinou, J.-C. GRSF1 Regulates RNA Processing in Mitochondrial RNA Granules. Cell Metab. 2013, 17, 399–410. [Google Scholar] [CrossRef]
- Minczuk, M.; He, J.; Duch, A.M.; Ettema, T.J.; Chlebowski, A.; Dzionek, K.; Nijtmans, L.G.J.; Huynen, M.A.; Holt, I.J. TEFM (C17orf42) Is Necessary for Transcription of Human MtDNA. Nucleic Acids Res. 2011, 39, 4284–4299. [Google Scholar] [CrossRef]
- Jiang, S.; Koolmeister, C.; Misic, J.; Siira, S.; Kühl, I.; Silva Ramos, E.; Miranda, M.; Jiang, M.; Posse, V.; Lytovchenko, O.; et al. TEFM Regulates Both Transcription Elongation and RNA Processing in Mitochondria. EMBO Rep. 2019, 20, e48101. [Google Scholar] [CrossRef] [PubMed]
- Rackham, O.; Davies, S.M.K.; Shearwood, A.-M.J.; Hamilton, K.L.; Whelan, J.; Filipovska, A. Pentatricopeptide Repeat Domain Protein 1 Lowers the Levels of Mitochondrial Leucine TRNAs in Cells. Nucleic Acids Res. 2009, 37, 5859–5867. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Itoh, Y.; Del’Olio, S.; Hassan, A.; Naschberger, A.; Flygaard, R.K.; Nobe, Y.; Izumikawa, K.; Aibara, S.; Andrell, J.; et al. Structure of Mitoribosome Reveals Mechanism of MRNA Binding, TRNA Interactions with L1 Stalk, Roles of Cofactors and RRNA Modifications. bioRxiv 2023. [Google Scholar] [CrossRef]
- Greber, B.J.; Boehringer, D.; Leitner, A.; Bieri, P.; Voigts-Hoffmann, F.; Erzberger, J.P.; Leibundgut, M.; Aebersold, R.; Ban, N. Architecture of the Large Subunit of the Mammalian Mitochondrial Ribosome. Nature 2014, 505, 515–519. [Google Scholar] [CrossRef]
- Bruni, F.; Proctor-Kent, Y.; Lightowlers, R.N.; Chrzanowska-Lightowlers, Z.M. Messenger RNA Delivery to Mitoribosomes—Hints from a Bacterial Toxin. FEBS J. 2021, 288, 437–451. [Google Scholar] [CrossRef]
- Montoya, J.; Ojala, D.; Attardi, G. Distinctive Features of the 5′-Terminal Sequences of the Human Mitochondrial MRNAs. Nature 1981, 290, 465–470. [Google Scholar] [CrossRef]
- Brown, A.; Amunts, A.; Bai, X.C.; Sugimoto, Y.; Edwards, P.C.; Murshudov, G.; Scheres, S.H.W.; Ramakrishnan, V. Structure of the Large Ribosomal Subunit from Human Mitochondria. Science 2014, 346, 718–722. [Google Scholar] [CrossRef]
- Remes, C.; Khawaja, A.; Pearce, S.F.; Dinan, A.M.; Gopalakrishna, S.; Cipullo, M.; Kyriakidis, V.; Zhang, J.; Dopico, X.C.; Yukhnovets, O.; et al. Translation Initiation of Leaderless and Polycistronic Transcripts in Mammalian Mitochondria. Nucleic Acids Res. 2023, 51, 891–907. [Google Scholar] [CrossRef]
- Jones, C.N.; Wilkinson, K.A.; Hung, K.T.; Weeks, K.M.; Spremulli, L.L. Lack of Secondary Structure Characterizes the 5′ Ends of Mammalian Mitochondrial MRNAs. RNA 2008, 14, 862–871. [Google Scholar] [CrossRef]
- Hirano, M.; Emmanuele, V.; Quinzii, C.M. Emerging Therapies for Mitochondrial Diseases. Essays Biochem. 2018, 62, 467–481. [Google Scholar] [CrossRef]
- Perli, E.; Pisano, A.; Pignataro, M.G.; Campese, A.F.; Pelullo, M.; Genovese, I.; Turris, V.; Ghelli, A.M.; Cerbelli, B.; Giordano, C.; et al. Exogenous Peptides Are Able to Penetrate Human Cell and Mitochondrial Membranes, Stabilize Mitochondrial TRNA Structures, and Rescue Severe Mitochondrial Defects. FASEB J. 2020, 34, 7675–7686. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Davidson, E.; King, M.P. The Pathogenic A3243G Mutation in Human Mitochondrial TRNALeu(UUR) Decreases the Efficiency of Aminoacylation. Biochemistry 2003, 42, 958–964. [Google Scholar] [CrossRef]
- Suzuki, T.; Nagao, A.; Suzuki, T. Human Mitochondrial Diseases Caused by Lack of Taurine Modification in Mitochondrial TRNAs. Wiley Interdiscip. Rev. RNA 2011, 2, 376–386. [Google Scholar] [CrossRef]
- Peng, G.-X.; Mao, X.-L.; Cao, Y.; Yao, S.-Y.; Li, Q.-R.; Chen, X.; Wang, E.-D.; Zhou, X.-L. RNA Granule-Clustered Mitochondrial Aminoacyl-TRNA Synthetases Form Multiple Complexes with the Potential to Fine-Tune TRNA Aminoacylation. Nucleic Acids Res. 2022, 50, 12951–12968. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Wilson, B.; Kumar, P.; Dutta, A. Non-Canonical Roles of TRNAs: TRNA Fragments and Beyond. Annu. Rev. Genet. 2020, 54, 47–69. [Google Scholar] [CrossRef]
- Amunts, A.; Brown, A.; Toots, J.; Scheres, S.H.W.; Ramakrishnan, V. The Structure of the Human Mitochondrial Ribosome. Science 2015, 348, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Greber, B.J.; Bieri, P.; Leibundgut, M.; Leitner, A.; Aebersold, R.; Boehringer, D.; Ban, N. Ribosome. The Complete Structure of the 55S Mammalian Mitochondrial Ribosome. Science 2015, 348, 303–308. [Google Scholar] [CrossRef]
- Rorbach, J.; Gao, F.; Powell, C.A.; D’Souza, A.; Lightowlers, R.N.; Minczuk, M.; Chrzanowska-Lightowlers, Z.M. Human Mitochondrial Ribosomes Can Switch Their Structural RNA Composition. Proc. Natl. Acad. Sci. USA 2016, 113, 12198–12201. [Google Scholar] [CrossRef] [PubMed]
- Bruni, F. Mitochondria: From Physiology to Pathology. Life 2021, 11, 991. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loguercio Polosa, P.; Capriglia, F.; Bruni, F. Molecular Investigation of Mitochondrial RNA19 Role in the Pathogenesis of MELAS Disease. Life 2023, 13, 1863. https://doi.org/10.3390/life13091863
Loguercio Polosa P, Capriglia F, Bruni F. Molecular Investigation of Mitochondrial RNA19 Role in the Pathogenesis of MELAS Disease. Life. 2023; 13(9):1863. https://doi.org/10.3390/life13091863
Chicago/Turabian StyleLoguercio Polosa, Paola, Francesco Capriglia, and Francesco Bruni. 2023. "Molecular Investigation of Mitochondrial RNA19 Role in the Pathogenesis of MELAS Disease" Life 13, no. 9: 1863. https://doi.org/10.3390/life13091863
APA StyleLoguercio Polosa, P., Capriglia, F., & Bruni, F. (2023). Molecular Investigation of Mitochondrial RNA19 Role in the Pathogenesis of MELAS Disease. Life, 13(9), 1863. https://doi.org/10.3390/life13091863