Abstract
Canine babesiosis is an emerging tick-borne disease of major veterinary concern in Europe. Its prevalence has increased in the last two decades and is spreading rapidly toward the north. The aim of this study was to investigate the genetic diversity of Babesia spp. strains isolated from naturally infected dogs in a tick-endemic area (Dobrogea) in southeastern Romania. For this purpose, a total of twenty-three samples from dogs diagnosed with various clinical forms of babesiosis, evaluated by means of clinical history, physical examination, and hematological tests, were subjected to a molecular investigation using PCR, sequencing analysis, and genetic characterization. A microscopic examination of thin Diff-quick-stained blood smears revealed large intra-erythrocytic Babesia piroplasms in all dogs. The PCR and sequencing analysis results indicated the presence of Babesia canis in 22 dogs (95.7%) and Babesia vogeli in 1 dog (4.3%). Among the B. canis isolates, two genotypes were distinguished based on two nucleotide substitutions (GA→AG) observed in the 18S rRNA gene sequences (at positions 609 and 610), with the AG genotype predominating (54.5% of samples), while the GA variant was identified in 9.1% of samples. In the remaining isolates (36.4%), both variants were identified. The B. vogeli-positive dog also tested positive for antibodies against Ehrlichia canis and displayed severe disease. This study reports, for the first time, the presence of genetically heterogenic B. canis strains in dogs with clinical babesiosis in Romania. These findings provide a basis for future studies on the relationship between the genetic structure of the causative agents of canine babesiosis in Romania and the course of the disease.
1. Introduction
Canine babesiosis (CB) is a tick-borne disease that significantly impacts dogs’ health worldwide [1,2]. It is caused by large and small intra-erythrocytic protozoa (Apicomplexa: Piroplasmida) vectored by different tick species. The prevalence of babesiosis and its geographical distribution generally vary according to the tick spectrum in a given area [3,4].
Currently, CB is increasingly reported in many European countries as an emerging and rapidly expanding disease [2,5]. Moreover, several recent studies have reported the spreading and/or occurrence of CB in northern countries, such as Belgium [6], the Netherlands [7], and Norway [8], which are related to the geographic expansion of the tick vector Dermacentor reticulatus [9]. These findings emphasize the need for the continuous monitoring of CB in both endemic and non-endemic areas.
The clinical presentation of babesiosis in dogs may range from subclinical to acute, even fatal, disease [3]. The severity of infection is mainly attributed to the pathogenicity of the causative Babesia species [10,11]; however, other factors related to the dog’s age, immunity, and concomitant infections also play a role in the variable pathogenicity of piroplasms infecting dogs [12,13].
In addition to the Babesia-induced hemolytic anemia and hypoxia, some dogs develop a complicated babesiosis characterized by immune-mediated hemolytic anemia (IMHA) and/or signs of inflammatory reactions [14,15]. Subsequently, abnormalities seen in complicated canine babesiosis cases include hepatopathy, acute kidney injury (AKI), acute respiratory distress syndrome (ARDS), cerebral babesiosis, pancreatitis, rhabdomyolysis, and myocardial dysfunction [16].
Traditionally, laboratory diagnosis has relied on the microscopic detection of intra-erythrocytic piroplasms. However, this technique is laborious, time-consuming, and has limitations, depending on the disease stage. For instance, this method is more accurate during the acute stage of the disease [1]. On the other hand, the molecular diagnosis shows high sensitivity and specificity, making it useful and adequate to detect low parasitemia levels (e.g., during the subclinical or chronic disease stages) [17]. The amplification of parasite DNA indicates evidence of infection; moreover, the sequencing results are more reliable and allow differentiation between the large Babesia piroplasm species (i.e., B. canis and B. vogeli), which are morphologically indistinguishable [10,18,19].
In Romania, the epidemiology of CB has shown a continuous dynamic in the last two decades, progressing from sporadic cases to endemic areas, such as southeastern Romania, where an increasing number of clinical CB cases are reported [20,21,22,23]. However, limited data are available on the genetic characterization of the Babesia species associated with various clinical presentations of CB in Romania. Therefore, the aim of the present study was to provide new insights into the molecular epidemiology of clinical canine babesiosis, with contributions from the molecular detection, sequencing analysis, and genetic characterization of Babesia strains isolated from naturally infected Romanian dogs.
2. Materials and Methods
2.1. Study Design
Twenty-three client-owned dogs with clinical babesiosis were included in this study. These dogs were exclusive patients of the study clinic, originated from the Dobrogea region (southeastern Romania), and had no history of travelling outside Romania.
Dobrogea is a historical region in the coastal area of the Black Sea in southeastern Romania, located between the lower Danube River and the Black Sea.
Data about each dog’s breed, age, lifestyle, tick infestation history, and outdoor access were registered.
The clinical history, physical examination, and laboratory testing were corroborated to confirm the diagnosis.
A molecular analysis of the partial 18S rRNA gene was performed for species identification and genetic characterization.
2.2. Samples and Laboratory Investigations
Whole peripheral (cephalic vein) blood samples were collected from the dogs and subjected to laboratory investigations.
The diagnosis of babesiosis was established by the presence of intra-erythrocytic parasites in thin blood smears stained with Diff-quick stain [20].
To assess the hemato-biochemical alterations, selected hematological (blood cell count) and biochemical parameters were determined using automatic hematology (Abacus Vet Jr., Diatron MI ZRT, Budapest, Hungary) and biochemistry (SPOTCHEM™ EZ SP-4430, Arkray, Europe, Amstelveen, The Netherlands) analyzers, respectively.
Depending on the clinical presentation (clinical signs such as fever, depression, anorexia, and pale mucous membranes) and the severity of anemia, the cases were classified as mild or severe disease. The severity of anemia (lower values for the packed cell volume (PCV) and/or hemoglobin (Hgb)) was defined as follows: mild (30% ≤ PCV < 37% and/or 10 ≤ Hgb < 12 g/dL), moderate (20% ≤ PCV < 30% and/or 5 ≤ Hgb < 10 g/dL), or severe (PCV < 20% and/or Hgb < 5 g/dL).
The severity of thrombocytopenia (lower values for the platelet count (PLT)) was defined as follows: mild (100 ≤ PLT < 200 × 109/L), moderate (50 ≤ PLT < 100 × 109/L), or severe (PLT < 50 × 109/L).
Additionally, we classified the cases based on detected clinicopathological changes as (i) uncomplicated babesiosis when varying degrees of hemolytic anemia were present, (ii) complicated babesiosis with single organ dysfunction, and (iii) complicated babesiosis with multiple organ dysfunction syndrome (MODS), as previously described [14,23,24,25]. Briefly, for defining complicated babesiosis, in addition to the hemolytic anemia, the following criteria for organ dysfunction were used: (i) for acute renal injury: creatinine ≥ 1.5 mg/dL; (ii) for hepatopathy: two elevated liver enzymes (alkaline phosphatase (AP) > 280 U/L and alanine aminotransferase (ALT) > 60) and/or aspartate aminotransferase (AST) or a single elevated enzyme (AP > 560 U/L or ALT > 120 U/L); (iii) clinical signs of acute respiratory distress syndrome (ARDS). MODS was considered when at least two of the above dysfunctions were present.
To investigate potential coinfections and the impact of other vector-borne diseases on clinical presentation severity, the dogs were also tested using a commercial ELISA-based kit (SNAP® 4Dx®, IDEXX Laboratories, Inc. Westbrook, ME, USA). This kit can simultaneously detect circulating antibodies against Anaplasma spp., Borrelia burgdorferi sensu lato, and Ehrlichia canis (three tick-borne pathogens) and the circulating antigen of Dirofilaria immitis (a mosquito-borne pathogen).
EDTA-anticoagulated peripheral blood was stored at −20 °C for subsequent DNA extraction and analysis.
2.3. DNA Isolation, PCR Amplification, and Sequencing
Twenty-three dog blood samples were subjected to the molecular detection of Babesia spp. DNA and a sequence analysis. For this, total genomic DNA was extracted from 200 μL of EDTA whole blood from each sample using a DNeasy Tissue kit (Qiagen AG, Basel, Switzerland) according to the manufacturer’s instructions. The DNA samples were stored at −20 °C until processing.
For the molecular detection of Babesia DNA, a conventional polymerase chain reaction (PCR) method was conducted using a genus-specific set of primers that amplify a specific region of the 18S rRNA gene of Babesia spp. of approx. 450 bp [26].
The PCR reactions were performed on a Mini Thermal Cycler MJ (Bio-Rad®, Hercules, CA, USA) in a reaction volume of 50 μL containing 25 μL of 2X Taq PCR Master Mix (Qiagen AG, Basel, Switzerland), 0.5 μM of each primer, and 5 μL of the extracted DNA as a template. The cycle conditions were described previously [20].
The PCR amplicons were visualized on a 2% agarose gel and stained with ethidium bromide.
Positive PCR products were purified using a QIAQuick PCR purification kit (Qiagen, Hilden, Germany) and subsequently sequenced in both directions (CeMIA, Larissa, Greece).
2.4. DNA Sequence and Phylogenetic Analysis
The obtained electropherograms were viewed and edited using GENtle (Manske, University of Cologne). The sequences were compared to the deposited reference sequences in GenBank using the online tool BLAST (National Center for Biotechnology Information; https://www.ncbi.nlm.nih.gov/ accessed on 6 May 2023).
Phylogenetic analyses were performed with reference sequences of Babesia canis from GenBank using the neighbor-joining method with the Jukes–Cantor substitution model. Babesia gibsoni was used as an outgroup. The phylogenetic tree was tested with 1000 bootstrap replications. The analysis was conducted in MEGA X [27].
2.5. Statistical Analysis
For the statistical analysis, Fisher’s exact test was used (Quantitative Parasitology 3.0 free software) [28]. p ≤ 0.05 was considered statistically significant.
3. Results
3.1. Clinical Cases
A total of 23 dogs (17 males and 6 females) of different breeds, varying from 3 months to 13 years of age (mean age = 4.2 years; median = 2.0), that displayed various clinical signs of babesiosis were evaluated by means of clinical history, physical examination, and hematological testing.
Blood samples obtained from the dogs were also subjected to PCR, a sequencing analysis, and genetic characterization. All 23 dogs were diagnosed between February and June 2019, predominantly in April, May, and June (n = 17) but also in February and March (3 dogs each). A summary of the epidemiological data and the clinical presentation is presented in Table 1.

Table 1.
Descriptive epidemiological data on microscopically (blood smear) and molecularly (PCR and sequencing) Babesia-positive dogs in southeastern Romania (data are stratified by clinicopathological groups).
A microscopic examination of thin stained blood smears from all 23 samples revealed single and/or paired intra-erythrocytic microorganisms that were morphologically consistent with large Babesia piroplasms.
Two dogs also tested serologically positive for antibodies against other tick-borne pathogens, namely, E. canis and Anaplasma spp.
3.2. PCR and DNA Sequence Analysis
Positive PCR results were obtained for all canine blood samples. The length of the PCR product corresponded to the expected size, at approximately 450 bp.
All PCR products were sequenced, and the BLASTn analysis of the sequences revealed the presence of two Babesia species, namely, Babesia canis in 22/23 samples (95.7%) and Babesia vogeli in 1 sample (4.3%) (Table 1).
The nucleotide sequence analysis showed that two different genetic variants could be distinguished among the B. canis isolates based on two nucleotide substitutions (GA→AG) occurring in the 18S rRNA gene sequence. The two genotypes differ at two consecutive sites, 129 and 130, in our gene fragment [26], which, if the complete B. canis 18S rRNA gene sequence is taken as a reference (GenBank accession No. AY072926) [29], are located at nucleotide positions 609 and 610 of the whole gen (Table 2). Twelve samples (54.5%; 12/22) showed the bases adenine and guanine (AG), while two samples (9.1%; 2/22) showed guanine and adenine (GA) at these positions. In the remaining 8/22 samples (36.4%), both adenine and guanine were observed at each of these positions in the electropherogram, displaying A/G double peaks (RR), which may indicate infection with both variants.
Table 2.
Differences in the nucleotide sequences of the 18S rRNA gene of the Babesia canis isolates analyzed in this study (#) and the partial B. canis 18S rRNA gene reference (AY072926). Positions are numbered with reference to the full-length sequence.
The BLAST analysis of the AG sequences showed 100% similarity to isolates previously identified in Romania [20] as well as other countries, such as Slovenia [30], Hungary [31], Russia [32], Poland [33], Croatia [34], Estonia [35], Austria [36], the Republic of Moldavia [37], and Turkey (MN704759.1; KF499115.1).
The GA variant also showed identity with isolates from other countries, such as France [38], Italy [29], the Netherlands [7], Croatia [34], Lithuania [39], Poland [40], Slovakia (MK5088770), Spain (MK591947), Latvia [41], and Russia [32]. The sequence alignment of the partial 18S rDNA sequence of the B. canis isolates analyzed in this study that showed differences (AG/GA/RR) in comparison with the reference gene sequence (AY072926) is depicted in Table 2. The phylogenetic analysis showed that the sequences from this study were identical to other B. canis isolates that display the AG and GA genotypes (Figure 1).
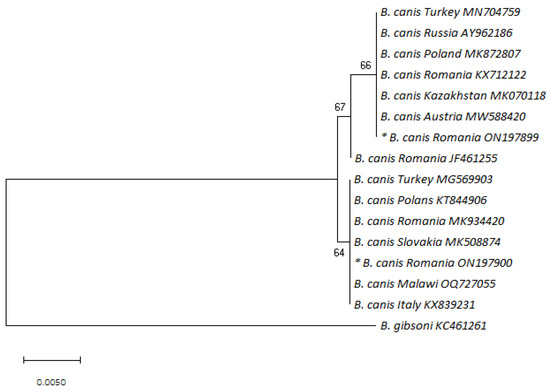
Figure 1.
Phylogenetic tree of Babesia canis based on neighbor-joining analysis of 447 bp long 18S rRNA fragments; B. gibsoni was used as an outgroup. Sequence names consist of the species, the country of origin, and the corresponding accession number. Bootstrap values are shown on the nodes. The sequences discovered in the present study are marked with an asterisk (nucleotide sequence * ON197899 corresponds to the AG variant; nucleotide sequence * ON197900 corresponds to the GA variant).
The B. vogeli sequence was identical to the reference sequence JF461252, which was previously detected in Romania [20]. This genetic variant was also found in Egypt [42] and Israel [43].
The partial 18S rRNA sequences of the AG and GA B. canis variants and the B. vogeli variant were deposited in GenBank (accession numbers: ON197899.1, ON197900.1, and ON197901.1, respectively).
3.3. Clinicopathological Findings in B. canis-Infected Dogs
All B. canis-positive dogs were symptomatic, displaying clinical signs consistent with canine babesiosis. All the dogs showed lethargy and anorexia, while fever and hemoglobinuria were registered in 95.5% (21/22) and 68.2% (15/22) of the cases, respectively.
The dogs showed various degrees of clinical presentation: mild in 27.3% (6/22), moderate in 31.8% (7/22), and severe in 40.1% (9/22) of cases. Additionally, according to the pathological findings (hematological and biochemical abnormalities), 40.9% (9/22) of the dogs were diagnosed with uncomplicated babesiosis, whereas 59.1% (13/22) of the dogs were diagnosed with complicated babesiosis, with either single organ dysfunction (SOD) (40.9%; 9/22) or multiple organ dysfunction syndrome (MODS) (18.2%; 4/22) (Table 1). Based on the defining criteria, the following complications were diagnosed: acute renal injury was diagnosed in nine dogs as single organ dysfunction (n = 5) or MODS (n = 4); hepatopathy was diagnosed in eight dogs as single (n = 4) or multiple (n = 4) organ dysfunction; and ARDS was diagnosed in one case as MODs.
The most common hematological changes in B. canis-positive dogs included normocytic normochromic anemia (63.6%; 14/22) and thrombocytopenia (59.1%; 13/22). Dogs with anemia were classified as mild (14.2%), moderate (57.1%), and severe (28.6%). Of the dogs with thrombocytopenia, severe thrombocytopenia (PLT < 50 × 109/L) was registered in >50% (53.8%; 7/13) of cases (Table 3).
Table 3.
Relative frequencies of common pathological changes in 22 Babesia canis-positive dogs in southeastern Romania.
Regarding the biochemical abnormalities, the most frequent changes included hyperbilirubinemia in 72.7% (16/22) of cases and azotemia (increased serum creatinine and/or blood urea nitrogen (BUN)) in 59.1% (13/22) of cases (for nine dogs, concurrent increased creatinine and BUN were registered, while four showed increased BUN). Hepatic enzymes (alanine aminotransferase (ALT), alkaline phosphatase (AP), and/or aspartate aminotransferase (AST)) were elevated in 45.4% (10/22) of cases, while pancreatic amylase was increased in 4.5% (2/22) of dogs.
The clinicopathological findings were compared to the B. canis genotypes, and a statistically significant relationship was found between the genotype and clinical presentation severity (p = 0.02), severe anemia, and severe thrombocytopenia (p < 0.05), with a higher severity for the GA genotype (Table 2). However, since the GA genotype was found in only two cases, further studies are necessary to demonstrate the genotype-associated risks for severe disease.
No statistically significant correlation between the genotype and complications (complicated babesiosis with SOD and/or MOD) was found (p > 0.05).
Table 3 summarizes the data on the most frequent complications and hematological and biochemical alterations among B. canis-positive dogs.
3.4. Clinicopathological Findings in the B. vogeli-Infected Dog
The B. vogeli-positive dog (a seven-year-old male mongrel) also tested positive for antibodies against E. canis, a tick-borne pathogen also vectored by R. sanguineus. This dog had severe anemia, thrombocytopenia, and elevated hepatic (ALT and AST) and pancreatic (AMY) enzyme values.
The B. vogeli-positive dog displayed a severe disease characterized by lethargy, anorexia, fever (39.6 °C), severe anemia (PCV = 9.1% and Hgb = 2.5 g/dL), and severe thrombocytopenia (PLT = 17 × 109/L).
4. Discussion
As demonstrated in previous studies, CB is recognized as an endemic disease in southeastern Romania, including the study area [20,23]. The findings of the present study confirm the presence of both large Babesia piroplasm species, namely, B. canis and B. vogeli, in dogs with clinical babesiosis [44,45,46].
B. canis is recognized as the most common agent of canine babesiosis in Europe [2]. Our molecular analysis also identified B. canis as the most common causative agent of babesiosis in symptomatic dogs in the investigated region.
Additionally, to our knowledge, this is the first molecular study on the genetic diversity of B. canis strains isolated from naturally infected dogs in Romania by amplifying and sequencing a portion of the 18S ribosomal rRNA gene. Subsequently, our results indicated the presence of two different genotypes of B. canis in Romanian dogs. These genotypes differ by two nucleotide substitutions (GA→AG) at positions 129 and 130 of the partial 18S rRNA gene sequence obtained in our study, as shown in Table 2.
Previous studies have also observed that B. canis can differ at these sequence positions. Although they did not find differences in the entire length of the 18S rRNA gene between Croatian, Italian, and Polish isolates, their complete 18S rRNA sequence (AY072926) served as a reference for other studies [29]. Thus, when comparing their Slovenian samples to the reference AY072926, Duh et al. [30] found three genotypes differing at two consecutive positions, 609 and 610 (GA/AG/AA), of the complete gene [29,30]. Since then, other molecular surveys have reported on this single-nucleotide polymorphism as an intra-specific genetic variation of B. canis isolates from different European countries, such as Poland [33,47], Croatia [34], Switzerland [48], Lithuania [39,49], Latvia [41], and Russia [32].
Currently, four genotypes of the B. canis 18S rRNA gene with nucleotide substitutions (GA/AG/AA/TT) at these positions are reported in Europe. However, since most studies, including this one, have not examined the complete 18S rRNA gene, it cannot be excluded that there are additional genotypes. The B. canis genotypes found in our study correspond to the previously reported molecular findings on the nucleotide polymorphism related to the GA→AG substitutions (Table 2).
Infections with only the AG genotype were most common and were revealed in 54.5% (12/22) of the isolates in our study. The AG sequence was identical to dog isolates previously identified in Romania [20] and others reported in Central and Eastern Europe with prevalence values ranging from 23.4% (23/98 isolates) in Croatia [34] to 35.0% (84/240 isolates) in Poland [47] and up to 54.5% (6/11 isolates) in Slovenia [30]. The AG genotype was also reported as the most prevalent, at 80.9% (17/21 isolates), among dog isolates in Russia [32].
The GA variant, displaying guanine and adenine at the aforementioned positions, was found in our study as a single infection in only two samples (9.1%). This genetic variant was detected for the first time in Romanian dogs but is considered common in other European countries where it is found most often, such as Croatia, at 58.1% (57/98 isolates); Poland, at 44.6% (107/240 isolates); and Lithuania, at 34.2% (13/38 isolates) [34,39,47].
The double peaks (RR) found in eight of the dog isolates (36.4%) in the present study have also previously been described in studies from Croatia (18.3%; 18/98 isolates) [34], Lithuania (65.8%; 25/38 isolates) [39], Latvia (91%; 39/43 isolates) [41], and Switzerland (100%; 19/19 isolates) [48]. The presence of both nucleotides (A/G, RR) at these positions can be explained either by a mixed infection with two B. canis genotypes or by the possibility of genetic heterogeneity occurring among the 18S rRNA gene copies [34,48]. In this respect, investigations performed on some Babesia species identified three (or four) copies of the ssrRNA genes, i.e., three ribosomal RNA (rRNA) transcription units (rDNA units) in Babesia bigemina and Babesia bovis and four in B. canis [50,51]. However, very limited heterogeneity in the ssrRNA copies within each species was reported [50]. Since all dogs in our study originated from the same area and ticks could harbor different B. canis genotypes, mixed infections seem to be the more likely explanation of the present finding.
The clinicopathological changes in B. canis-infected dogs in our study were consistent with canine babesiosis. The most common abnormalities were anemia (63.6%), thrombocytopenia (59.1%), hyperbilirubinemia (72.7%), and azotemia (59.1%).
When the clinicopathological changes were compared with the B. canis genotypes, a statistically significant correlation was found between the genotype and the severity of the clinical presentation, anemia, and thrombocytopenia (p < 0.05). However, no correlation between the genotype and babesiosis complications (p > 0.05) was found. In a recent study on the genetic analysis of B. canis strains in dogs in Poland, there were four polymorphic groups reported based on two nucleotide substitutions in the 18S rRNA gene fragment (GA, AG, TT, and a fourth with variable nucleotides). Of these, the GA (107 isolates), AG (84 isolates), and TT (7 isolates) genotypes were associated with acute babesiosis. In contrast, representatives of the TT (31 isolates) and fourth (11 isolates) groups were associated with atypical and subclinical babesiosis, respectively [47]. Similar studies analyzing the clinicopathological findings in 26 symptomatic dogs with acute babesiosis did not determine any relationship between the B. canis genotypes GA/RR and the disease’s severity [41]. In our study, the GA genotype was found to be more severe in relation to anemia and thrombocytopenia than the AG or mixed genotypes. However, the low number of single GA genotypes identified (only two isolates) could have biased the results. Therefore, the relationship between the genetic structure of the protozoan pathogen and the disease course in dogs still requires future research studies, particularly regarding its diagnostic and/or prognostic value.
Apart from B. canis, we also molecularly detected B. vogeli in naturally infected dogs.
B. vogeli is vectored by Rhipicephalus sanguineus [10] and has mainly been detected in tropical and subtropical areas of Africa, Europe, Asia, and North and South America [42,52]. Although B. vogeli is considered the least pathogenic large species of Babesia, leading to mild or moderate and often clinically unapparent or subclinical infections, at least in adult dogs, it can cause severe disease in young dogs and when coinfections occur [53,54,55].
The clinicopathological findings of our study are in agreement with a recent study of the clinicopathological profile of B. vogeli infection and B. vogeli/E. canis coinfection [55]. In their study, a hematological data comparison between the only B. vogeli-positive group and the E. canis coinfection group showed statistically significant differences in red blood cell (RBC) parameters, including the RBC count, hemoglobin concentration, hematocrit, and RBC distribution width. However, the pathogenic mechanisms underlying this infection, such as the destruction of RBCs, require further investigation.
The mixed infection in the dog with B. vogeli and E. canis found in our study suggests a more severe clinical disease. Coinfections with other vector-borne pathogens have also been reported in recent studies of dogs in southeastern Romania [21,22]. Since coinfections often result in higher pathogenicity and complications, coinfected cases are of major clinical relevance [56].
These findings suggest that mixed infections, namely, Babesia spp. and other vector-borne pathogens, can be expected in Romanian dogs. Therefore, the simultaneous testing of dogs for multiple vector-borne pathogens is required for accurate diagnosis and prognosis and effective therapy.
The detection of both causative agents of canine babesiosis in Romanian dogs, namely, B. canis and B. vogeli, is in agreement with the sympatric occurrence and distribution of the two different tick vectors for the two Babesia species, i.e., D. reticulatus for B. canis and R. sanguineus for B. vogeli, in the study area, as recently reported [23,57]. D. reticulatus and R. sanguineus are the most common ticks infesting dogs in Romania, especially in southeastern Romania. The tick population has reportedly increased in recent years, including in the investigated area [23,57,58]. As none of the examined dogs had traveled outside Romania, we presumed that the B. canis and B. vogeli infections were autochthonously acquired.
5. Conclusions
This study reports, for the first time, the presence of two different B. canis genotypes in dogs with clinical babesiosis in Romania. The mixed infection with B. vogeli and E. canis of a symptomatic dog suggests that the simultaneous testing of dogs for multiple vector-borne pathogens is required for accurate diagnosis and prognosis and effective therapy. These findings provide a basis for future studies on the relationship between the genetic structure of the causative agents of canine babesiosis in Romania and the course of the disease.
Author Contributions
Conceptualization, M.I. and I.L.M.; methodology, M.I., L.L., M.W. and I.L.M.; investigation, L.L., E.M. and I.M.N.; validation, M.I., M.W. and I.L.M.; writing—original draft preparation, M.I.; writing—review and editing, M.I., M.W., I.M.N., E.M., L.L. and I.L.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study protocol was approved by the Bioethics Commission of the Faculty of Veterinary Medicine, Bucharest (Nr.28/30.03.2023).
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included in this article. The accession numbers of the obtained DNA sequences are mentioned in the Results and are available in the GenBank (https://www.ncbi.nlm.nih.gov/nuccore, accessed on 6 May 2023).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Boozer, A.L.; Macintire, D.K. Canine babesiosis. Vet. Clin. N. Am. Small Anim. Pract. 2003, 33, 885–904. [Google Scholar] [CrossRef] [PubMed]
- Solano-Gallego, L.; Sainz, Á.; Roura, X.; Estrada-Peña, A.; Miró, G. A review of canine babesiosis: The European perspective. Parasites Vectors 2016, 9, 336. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.P. Canine babesiosis: From molecular taxonomy to control. Parasites Vectors 2009, 2 (Suppl. S1), S4. [Google Scholar] [CrossRef] [PubMed]
- Solano-Gallego, L.; Baneth, G. Babesiosis in dogs and cats–expanding parasitological and clinical spectra. Vet. Parasitol. 2011, 181, 48–60. [Google Scholar] [CrossRef]
- Schnittger, L.; Rodriguez, A.E.; Florin-Christensen, M.; Morrison, D.A. Babesia: A world emerging. Infect. Genet. Evol. 2012, 12, 1788–1809. [Google Scholar] [CrossRef]
- Halos, L.; Lebert, I.; Abrial, D.; Danlois, F.; Garzik, K.; Rodes, D.; Schillmeier, M.; Ducrot, C.; Guillot, J. Questionnaire-based survey on the distribution and incidence of canine babesiosis in countries of Western Europe. Parasite 2014, 21, 13. [Google Scholar] [CrossRef]
- Matjila, T.P.; Nijhof, A.M.; Taoufik, A.; Houwers, D.; Teske, E.; Penzhorn, B.L.; Lange, T.; Jongejan, F. Autochthonous canine babesiosis in The Netherlands. Vet. Parasitol. 2005, 131, 23–29. [Google Scholar] [CrossRef]
- Øines, Ø.; Storli, K.; Brun-Hansen, H. First case of babesiosis caused by Babesia canis canis in a dog from Norway. Vet. Parasitol. 2010, 171, 350–353. [Google Scholar] [CrossRef]
- Rubel, F.; Brugger, K.; Monazahian, M.; Habedank, B.; Dautel, H.; Leverenz, S.; Kahl, O. The first German map of georeferenced ixodid tick locations. Parasites Vectors 2014, 7, 477. [Google Scholar] [CrossRef]
- Uilenberg, G.; Franssen, F.F.J.; Perie, M.; Spanger, A.A.M. Three groups of Babesia canis distinguished and a proposal for nomenclature. Vet. Q. 1989, 11, 33–40. [Google Scholar] [CrossRef]
- Schetters, T.P.; Moubri, K.; Precigout, E.; Kleuskens, J.; Scholtes, N.C.; Gorenflot, A. Different Babesia canis isolates, different diseases. Parasitology 1997, 115, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Brandao, L.P.; Hagiwara, M.K.; Myiashiro, S.I. Humoral immunity and reinfection resistance in dogs experimentally inoculated with Babesia canis and either treated or untreated with imidocarb dipropionate. Vet. Parasitol. 2003, 114, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Penzhorn, B.L. Why is Southern African canine babesiosis so virulent? An evolutionary perspective. Parasites Vectors 2011, 4, 51. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, L.S.; Clark, A.I. The pathophysiology of canine babesiosis: New approaches to an old puzzle. J. S. Afr. Vet. Assoc. 1994, 65, 134–145. [Google Scholar]
- Reyers, F.; Leisewitz, A.L.; Lobetti, R.G.; Milner, R.J.; Jacobson, L.S.; van Zyl, M. Canine babesiosis in South Africa: More than one disease. Does this serve as a model for falciparum malaria? Ann. Trop. Med. Parasitol. 1998, 92, 503–511. [Google Scholar] [CrossRef]
- Köster, L.S.; Lobetti, R.G.; Kelly, P. Canine babesiosis: A perspective on clinical complications, biomarkers, and treatment. Vet. Med. Res. Rep. 2015, 6, 119–128. [Google Scholar]
- Shaw, E.S.; Michael, J.D.; Birtles, R.J.; Edward, B.B. Tick-borne infectious diseases of dogs. Trends Parasitol. 2001, 17, 74–80. [Google Scholar] [CrossRef]
- Hauschild, S.; Schein, E. The subspecies specificity of Babesia canis. Berl. Munch. Tierarztl. Wochenschr. 1996, 109, 216–219. [Google Scholar]
- Zahler, M.; Schein, E.; Rinder, H.; Gothe, R. Characteristic genotypes discriminate between Babesia canis isolates of differing vector specificity and pathogenicity to dogs. Parasitol. Res. 1998, 84, 544–548. [Google Scholar] [CrossRef]
- Ionita, M.; Mitrea, I.L.; Pfister, K.; Hamel, D.; Buzatu, C.M.; Silaghi, C. Canine babesiosis in Romania due to Babesia canis and Babesia vogeli: A molecular approach. Parasitol. Res. 2012, 110, 1659–1664. [Google Scholar] [CrossRef]
- Anghel, R.G.; Mitrea, I.L.; Ionita, M. Retrospective study on prevalence of canine vector borne diseases in Bucharest area. Rev. Rom. Med. Vet. 2016, 26, 27–32. [Google Scholar]
- Anghel, R.G.; Mitrea, I.L.; Ionita, M. Clinico-pathological findings in vector-borne pathogen co-infections in dogs, from Bucharest area. Sci. Works Ser. C Vet. Med. 2017, 63, 45–49. [Google Scholar]
- Leica, L.; Mitrea, I.L.; Ionita, M. Clinical occurrence of canine babesiosis in the coastal area of the Black Sea (Dobrogea) in southeastern Romania and associated epidemiological implications. J. Parasitol. 2019, 105, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Lobetti, R.G. Canine babesiosis. In Manual of Canine and Feline Haematology and Transfusion Medicine; Day, M., Mackin, A., Littlewood, J., Eds.; British Small Animal Veterinary Association: Gloucester, UK, 2000; pp. 85–91. [Google Scholar]
- Jacobson, L.S. The south african form of severe and complicated canine babesiosis: Clinical advances 1994–2004. Vet. Parasitol. 2006, 138, 126–139. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Rozsa, L.; Reiczigel, J.; Majoros, G. Quantifying parasites in samples of hosts. J. Parasitol. 2000, 86, 228–232. [Google Scholar] [CrossRef]
- Casati, S.; Sager, H.; Gern, L.; Piffaretti, J.C. Presence of potentially pathogenic Babesia sp. for human in Ixodes ricinus in Switzerland. Ann. Agric. Environ. Med. 2006, 13, 65–70. [Google Scholar]
- Caccio, S.M.; Antunovic, B.; Moretti, A.; Mangili, V.; Marinculic, A.; Baric, R.R.; Slemenda, S.B.; Pieniazek, N.J. Molecular characterisation of Babesia canis canis and Babesia canis vogeli from naturally infected European dogs. Vet. Parasitol. 2002, 106, 285–292. [Google Scholar] [CrossRef]
- Duh, D.; Tozon, N.; Pertovec, M.; Strasek, K.; Avsic-Zupanc, T. Canine babesiosis in Slovenia: Molecular evidence of Babesia canis canis and Babesia canis vogeli. Vet. Res. 2004, 35, 363–368. [Google Scholar] [CrossRef]
- Földvari, G.; Hell, E.; Farkas, R. Babesia canis canis in dogs from Hungary: Detection by PCR and sequencing. Vet. Parasitol. 2005, 127, 221–226. [Google Scholar] [CrossRef]
- Rar, V.A.; Maksimova, T.G.; Zakharenko, L.P.; Bolykhina, S.A.; Dobrotvorsky, A.K.; Morozova, O.V. Babesia DNA detection in canine blood and Dermacentor reticulatus ticks in southwestern Siberia, Russia. Vector Borne Zoonotic Dis. 2005, 5, 285–287. [Google Scholar] [CrossRef] [PubMed]
- Adaszek, L.; Winiarczyk, S. Molecular characterization of Babesia canis canis isolates from naturally infected dogs in Poland. Vet. Parasitol. 2008, 152, 235–241. [Google Scholar] [CrossRef]
- Beck, R.; Vojta, L.; Mrljak, V.; Marinculic, A.; Beck, A.; Zivicnjak, T.; Caccio, S.M. Diversity of Babesia and Theileria species in symptomatic and asymptomatic dogs in Croatia. Int. J. Parasitol. 2009, 39, 843–848. [Google Scholar] [CrossRef]
- Tiskina, V.; Capligina, V.; Must, K.; Berzina, I.; Ranka, R.; Jokelainen, P. Fatal Babesia canis canis infection in a splenectomized Estonian dog. Acta Vet. Scand. 2016, 58, 7. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hodzic, A.; Mrowietz, N.; Cezanne, R.; Bruckschwaiger, P.; Punz, S.; Habler, V.E.; Tomsik, V.; Lazar, J.; Duscher, G.G.; Glawischnig, W.; et al. Occurrence and diversity of arthropod-transmitted pathogens in red foxes (Vulpes vulpes) in western Austria, and possible vertical (transplacental) transmission of Hepatozoon canis. Parasitology 2018, 145, 335–344. [Google Scholar] [CrossRef]
- D’Amico, G.; Ionică, A.M.; Györke, A.; Dumitrache, M.O. Epidemiological Survey of the Main Tick-Borne Pathogens Infecting Dogs from the Republic of Moldova. Pathogens 2022, 11, 1267. [Google Scholar] [CrossRef]
- Carret, C.; Walas, F.; Carey, B.; Grande, N.; Precigout, E.; Moubri, K.; Schetters, T.P.; Gorenflot, A. Babesia canis canis, Babesia canis vogeli, Babesia canis rossi: Differentiation of the three subspecies by a restriction fragment length polymorphism analysis on amplified small subunit ribosomal RNA genes. J. Eukaryot. Microbiol. 1999, 46, 298–303. [Google Scholar] [CrossRef]
- Paulauskas, A.; Radzijevskaja, J.; Karveliene, B.; Grigonis, A.; Aleksandraviciene, A.; Zamokas, G.; Babickaitė, L.; Sabūnas, V.; Petkevičius, S. Detection and molecular characterization of canine babesiosis causative agent Babesia canis in the naturally infected dog in Lithuania. Vet. Parasitol. 2014, 205, 702–706. [Google Scholar] [CrossRef] [PubMed]
- Mierzejewska, E.J.; Pawelczyk, A.; Radkowski, M.; Welc-Faleciak, R.; Bajer, A. Pathogens vectored by the tick, Dermacentor reticulatus, in endemic regions and zones of expansion in Poland. Parasites Vectors 2015, 8, 490. [Google Scholar] [CrossRef]
- Seleznova, M.; Kivrane, A.; Namina, A.; Krumins, R.; Aleinikova, D.; Lazovska, M.; Akopjana, S.; Capligina, V.; Ranka, R. Babesiosis in Latvian domestic dogs, 2016–2019. Ticks Tick. Borne Dis. 2020, 11, 101459. [Google Scholar] [CrossRef]
- Passos, L.M.F.; Geiger, S.M.; Ribeiro, M.F.B.; Pfister, K.; Zahler-Rinder, M. First molecular detection of Babesia vogeli in dogs from Brazil. Vet. Parasitol. 2005, 127, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Mumcuoglu, K.Y.; Arslan-Akveran, G.; Aydogdu, S.; Karasartova, D.; Koşar, A.; Savci, U.; Keskin, A.; Taylan-Ozkan, A. Pathogens in ticks collected in Israel: II. Bacteria and protozoa found in Rhipicephalus sanguineus sensu lato and Rhipicephalus turanicus. Ticks Tick. Borne Dis. 2022, 13, 101986. [Google Scholar] [CrossRef] [PubMed]
- Imre, M.; Farkas, R.; Ilie, M.S.; Imre, K.; Darabus, G. Survey of babesiosis in symptomatic dogs from Romania: Occurrence of Babesia gibsoni associated with breed. Ticks Tick. Dis. 2013, 4, 500–502. [Google Scholar] [CrossRef]
- Cimpan, A.A.; Nachum-Biala, Y.; Ben-Shitrit, B.; Miron, L.; Baneth, G. Epidemiological study of canine babesiosis and hepatozoonosis in the South of Romania. Acta Parasitol. 2020, 65, 669–678. [Google Scholar] [CrossRef]
- Ciucă, L.; Martinescu, G.; Miron, L.D.; Roman, C.; Acatrinei, D.; Cringoli, G.; Rinaldi, L.; Maurelli, M.P. Occurrence of Babesia species and co-infection with Hepatozoon canis in symptomatic dogs and in their ticks in Eastern Romania. Pathogens 2021, 10, 1339. [Google Scholar] [CrossRef]
- Łyp, P.; Bartnicki, M.; Staniec, M.; Winiarczyk, S.; Adaszek, Ł. Occurrence of different strains of Babesia canis in dogs in eastern Poland. J. Vet. Res. 2016, 60, 423–427. [Google Scholar] [CrossRef][Green Version]
- Schaarschmidt, D.; Gilli, U.; Gottstein, B.; Marreros, N.; Kuhnert, P.; Daeppen, J.A.; Rosenberg, G.; Hirt, D.; Frey, C.F. Questing Dermacentor reticulatus harbouring Babesia canis DNA associated with outbreaks of canine babesiosis in the Swiss Midlands. Ticks Tick. Borne Dis. 2013, 4, 334–340. [Google Scholar] [CrossRef][Green Version]
- Radzijevskaja, J.; Mardosaitė-Busaitienė, D.; Aleksandravičienė, A.; Karvelienė, B.; Razgūnaitė, M.; Stadalienė, I.; Paulauskas, A. Genetic Diversity of Babesia canis Strains in Dogs in Lithuania. Microorganisms 2022, 10, 1446. [Google Scholar] [CrossRef]
- Reddy, G.; Chakrabarti, D.; Yowell, C.A.; Dame, J.B. Sequence microheterogeneity of the three small subunit ribosomal RNA genes of Babesia bigemina: Expression in erythrocyte culture. Nucleic Acids Res. 1991, 19, 3641–3645. [Google Scholar] [CrossRef] [PubMed]
- Dalrymple, B.P.; Dimmock, C.M.; Parrodi, F.; Wright, I.G. Babesia bovis, Babesia bigemina, Babesia canis, Babesia microti and Babesia rodhaini: Comparison of ribosomal RNA gene organization. Int. J. Parasitol. 1992, 22, 851–855. [Google Scholar] [CrossRef]
- Birkenheuer, A.J.; Levy, M.G.; Breitschwerdt, E.B. Development and evaluation of a seminested PCR for detection and differentiation of Babesia gibsoni (Asian genotype) and B. canis DNA in canine blood samples. J. Clin. Microbiol. 2003, 41, 4172–4177. [Google Scholar] [CrossRef]
- Schoeman, J.P. Canine babesiosis. Onderstepoort J. Vet. Res. 2009, 76, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Solano-Gallego, L.; Trotta, M.; Carli, E.; Carcy, B.; Caldin, M.; Furlanello, T. Babesia canis canis and Babesia canis vogeli clinicopathological findings and DNA detection by means of PCR-RFLP in blood from Italian dogs suspected of tick-borne disease. Vet. Parasitol. 2008, 157, 211–221. [Google Scholar] [CrossRef]
- Rawangchue, T.; Sungpradit, S. Clinicopathological and molecular profiles of Babesia vogeli infection and Ehrlichia canis coinfection. Vet. World 2020, 13, 1294–1302. [Google Scholar] [CrossRef] [PubMed]
- De Tommasi, A.S.; Otranto, D.; Dantas-Torres, F.; Capelli, G.; Breitschwerdt, E.B.; De Caprariis, D. Are vector-borne pathogen co-infections complicating the clinical presentation in dogs? Parasites Vectors 2013, 6, 97. [Google Scholar] [CrossRef] [PubMed]
- Ionita, M.; Silaghi, C.; Mitrea, I.L.; Edouard, S.; Parola, P.; Pfister, K. Molecular detection of Rickettsia conorii and other zoonotic spotted fever group rickettsiae in ticks, Romania. Ticks Tick. Borne Dis. 2016, 7, 150–153. [Google Scholar] [CrossRef]
- Ionita, M.; Mitrea, I.L.; Buzatu, M.C.; Dascalu, L. Seasonal dynamics of tick (Acari: Ixodidae) populations in different areas of Romania and the associated risks of tick-borne diseases. In Proceedings of the XIIth International Congress of Parasitology (ICOPA), Melbourne, VIC, Australia, 15–20 August 2010; Medimond International Proceedings: Bologna, Italy, 2010; pp. 91–95. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).